乙型肝炎病毒核心蛋白變構調節劑的作用機理及臨床研發和應用前景
2022-09-08 03:09:22魯鳳民郭巨濤
臨床肝膽病雜志 2022年8期
劉 慧, 魯鳳民, 郭巨濤
1 北京大學基礎醫學院 病原生物學系暨感染病中心, 北京 100191;2 巴魯克布隆伯格研究所, 美國賓夕法尼亞州 18902
HBV感染是我國病毒性肝炎、肝硬化和原發性肝細胞癌的主要病因之一。HBV在感染肝細胞的有效復制是維持病毒持續感染和致病的必需環節。作為構成病毒核衣殼的結構蛋白,核心蛋白(core protein,Cp)在HBV復制周期的多個環節發揮支持和調控功能。本文將系統闡述Cp在HBV復制過程中的功能和機理,及在研的靶向HBV Cp裝配的抗HBV新藥分類、作用機制、臨床研發現狀及對乙型肝炎功能性治愈的潛在價值,以期為該類新藥的研發和臨床試驗提供指導性意見。
1 HBV Cp在病毒復制周期的多個環節發揮重要功能
完整的有感染性的HBV為直徑42 nm的球形顆粒。外層為脂質囊膜,有3種囊膜蛋白(大、中、小表面抗原蛋白)鑲嵌其中。囊膜內為直徑約32 nm的正二十面體核衣殼。核衣殼內包裝有3.2 kb的部分雙鏈松弛環狀DNA(relaxed circular DNA,rcDNA)[1]。HBV僅感染人肝細胞,這主要是因為肝細胞表面具有HBV的特異性受體Na+-牛磺膽酸共轉運多肽(sodium taurocholate cotransporting polypeptide,NTCP)[2]。HBV病毒顆粒與肝細胞表面的NTCP結合,通過內吞途徑進入細胞。隨后病毒囊膜與內吞體膜融合將核衣殼釋放入細胞質中。核衣殼依次被轉運至核孔復合體并在此解聚,將rcDNA釋放入細胞核,并被修復產生共價閉合環狀DNA (cccDNA)[3]。進入細胞核內的cccDNA與核小體結合并經進一步修飾形成微小染色體, 轉錄產生5種病毒信使RNA(mRNA)并翻譯產生以下7種病毒蛋白:3.5 kb pre-C RNA,其翻譯產生precore蛋白,該蛋白經后續切割加工最終成熟為HBeAg而被分泌至細胞外;含兩個開放閱讀框架的3.5 kb pregenomic RNA (pgRNA),為典型的雙順反子轉錄本,通過其內部起始密碼子的差異使用分別翻譯產生Cp或病毒DNA聚合酶(polymerase,pol);2.4 kb RNA可翻譯產生大表面抗原蛋白(large HBsAg);2.1 kb RNA根據內部起始密碼子的差異使用翻譯產生中表面抗原蛋白(middle HBsAg)和小表面抗原蛋白(small HBsAg);0.7 kb RNA翻譯產生病毒X蛋白(HBx)[1]。
Cp蛋白由183個氨基酸殘基組成,通常以同源二聚體形式存在。120個Cp二聚體可自發裝配形成正20面體核衣殼[4]。作為病毒核衣殼的結構蛋白,Cp參與病毒復制的多個環節。第一,在HBV初始感染肝細胞過程中,Cp與微管轉運蛋白及importin-α/β相互識別介導核衣殼被轉運至核孔復合體。核孔蛋白與Cp相互作用誘導核衣殼解聚,并將病毒基因組rcDNA釋放入核形成cccDNA[5];第二,pgRNA和由其翻譯產生的病毒DNA聚合酶復合物核衣殼化,從而為病毒DNA復制提供了相對隔離的反應場所以阻止細胞固有免疫對新合成病毒DNA的模式識別,Cp本身也在病毒DNA的逆轉錄復制過程中發揮著重要的調控作用。在此過程中,宿主蛋白磷酸酶Ⅰ催化Cp羧基端結構域去磷酸化以調節Cp與pgRNA的相互作用,并與pgRNA-pol復合物一起被包裝入核衣殼[6]。在核衣殼內,pgRNA首先被逆轉錄產生負鏈DNA,并以此為模板進一步復制合成 rcDNA;第三,在HBV感染的肝細胞中,除包裝有pgRNA及處于復制不同階段病毒DNA的核衣殼之外, 絕大多數衣殼為不含病毒核酸的空衣殼。有別于核衣殼的裝配,在空衣殼裝配過程中并未發生Cp羧基端結構域去磷酸化。因此,與包裝有病毒RNA或DNA的核衣殼不同,空衣殼是由高度磷酸化的Cp組成[7];第四,含有rcDNA的核衣殼和空衣殼都可以通過Cp與病毒囊膜蛋白相互識別以啟動病毒顆粒的裝配和分泌[8]。最近的研究[9]發現,當成熟的rcDNA形成時,正二十面體核衣殼通過改變Cp蛋白的構象而與鑲嵌在多囊泡體膜上的病毒囊膜蛋白(preS)相互作用,從而獲得包膜并被釋放出細胞外,該過程受Cp第L60、L95、K96和I126等多個氨基酸殘基的調控。而位于Cp氨基端的裝配結構域和富含精氨酸的羧基端結構域之間的連接區可調控空病毒顆粒的包裝和分泌[10];第五,參與cccDNA池的內補充。與感染過程中進入細胞質中的核衣殼類似, 細胞質中復制產生的含有rcDNA的子代成熟核衣殼也可以脫衣殼, 將rcDNA轉運入細胞核內形成cccDNA,此即為cccDNA的細胞內擴增途徑[11]。由于感染來源的成熟病毒核衣殼和細胞內合成的子代成熟核衣殼的脫衣殼過程及位置有所差異,cccDNA的從頭合成和細胞內擴增途徑受細胞質中核酸酶的差異調控[12]。由此可見,通過核衣殼的包裝,脫衣殼以及介導核衣殼與病毒和細胞蛋白的相互作用,Cp 參與了病毒DNA復制,cccDNA合成和病毒顆粒的裝配等多個環節。
除此之外,也有研究[13]報道未參與衣殼組裝的游離Cp二聚體可作為cccDNA微小染色體的成分并調節cccDNA的轉錄活性。Cp亦被證實可招募干擾素誘導蛋白APOBEC 3A/3B至cccDNA,引起胞嘧啶脫氨化,導致cccDNA降解[14-15]。Cp也可與多種細胞RNA結合蛋白相互作用調節HBV RNA的代謝[16]。綜上所述,Cp功能的多樣性意味著靶向病毒Cp的抗病毒藥物具有抑制病毒復制周期的多個環節以及調控病毒蛋白和核酸代謝的多種潛能。
2 HBV Cp變構調節劑的發現及分類
靶向病毒Cp的抗HBV化合物是當前國內外處于研發階段的熱點新藥。但核心蛋白變構調節劑(core protein allosteric modulators,CpAM)的發現則可追溯到20世紀90年代后期。雖然第一個被發現的CpAM為苯丙烯酰胺類化合物(phenylpropenamides,PPA)[17],但目前處于臨床前和臨床研發階段的CpAM主要為雜芳基二氫嘧啶類化合物(heteroaryldihydropyrimidines,HAP)和氨磺酰苯甲酰胺類化合物(sulfamoylbenzamides,SBA)及其第二代和第三代的衍生物[18-19]。根據誘導Cp二聚體裝配的產物結構的不同,CpAM可分為兩種類型。Ⅰ型CpAM(typeⅠCpAM)主要是HAP衍生物,以Bay41-4109、GLS4等為代表,誘導Cp二聚體錯誤組裝形成多種形態的非衣殼結構聚合物,并最終在細胞內被降解[20-21]。Ⅱ型CpAM (typeⅡ CpAM)主要包括SBA及其衍生物,以DVR23(AB-423)、ABI-H0731、JNJ-56136379 和GLP-26等為代表[19,22],可誘導Cp二聚體裝配產生形態正常但未包裝pgRNA和病毒DNA聚合酶的空核衣殼。盡管這兩種類型的CpAM誘導產生的Cp二聚體包裝產物結構各異,但皆阻斷了病毒pgRNA和pol包裝入核衣殼及隨后的病毒DNA復制。
3 HBV Cp變構調節劑的作用靶點及抗病毒機理
Cp在肝細胞中以二聚體的形式作為核衣殼的基本結構單位。研究[23]表明,核衣殼的裝配過程由相鄰Cp二聚體之間的疏水性相互作用力驅動,首先發生緩慢聚合反應形成六聚體,再以此為基礎加速聚合裝配形成正20面體核衣殼結構。CpAM結合于相鄰Cp二聚體相互作用界面上的疏水性口袋(HAP pocket)以增強Cp二聚體之間的相互作用,加速Cp二聚體的裝配過程[24-25]。基于此,該類化合物被命名為CpAM或核衣殼裝配調節劑,而非核衣殼裝配抑制劑。HAP pocket由25個氨基酸殘基形成。因化學結構的差異,不同CpAM與“HAP”口袋中不同的氨基酸側鏈結合而干擾核衣殼組裝的速率和過程,從而導致裝配產物的表型差異。利用Y132A突變的Cp不能裝配形成核衣殼,但可形成穩定六聚體并與CpAM形成共結晶的特性,對比研究HAP_R01和SBA_R01與Cp的結合特性發現兩者雖皆結合Cp二聚體界面間的“HAP”口袋并與T33結合,但HAP_R01的噻唑基團可結合于其中的一個小口袋(sub-pocket),并與P25結合。與此結構生物學發現相對應,Cp氨基酸突變分析[26]證明Cp 的T33N突變對HAP_R01和SBA_R01均產生耐藥,而Cp的P25A或P25S突變只對HAP_R01耐藥。借助冷凍電鏡分析CpAM與體外裝配的空衣殼的相互作用,研究人員發現HAP1和AT-130均結合于HAP口袋并引起核衣殼四級結構的變化,但AT-130也可代償性地導致核衣殼裝配過程中三級結構的變化[24,27]。顯而易見,盡管這些結構生物學和耐藥突變分析研究結果可以輔助解釋兩種類型CpAM的表型和作用機理的差異,但CpAM對核衣殼裝配過程的調控機理有待于進一步生物物理學和結構生物學的研究。
除干擾核衣殼的裝配之外,CpAM在較高藥物濃度下也可以誘導已裝配的核衣殼結構改變甚至解聚[28-29]。如前所述,借助冷凍電鏡結構分析證實CpAM可結合于已裝配的核衣殼中的HAP口袋從而改變衣殼的三級和四級結構。這些結構的改變在一定情況下可以導致核衣殼在瓊脂糖凝膠電泳實驗中遷移率發生改變[30]。同時,可能是由于雙鏈DNA相對具有剛性并從核衣殼內部產生張力,使得CpAM結合HAP口袋后更易誘導成熟核衣殼解聚的緣故,有研究[31]發現CpAM可選擇性地誘導含有雙鏈DNA的成熟核衣殼的解聚。
與CpAM對核衣殼裝配和解聚的作用相符,在HBV感染的肝細胞中,CpAM誘導Cp裝配產生非衣殼結構聚合物或空衣殼從而干擾HBV DNA復制,最終抑制感染性病毒顆粒的產生。值得注意的是,CpAM因干擾pgRNA-Pol復合體包裝入核衣殼內,因此HBV RNA病毒樣顆粒的產生也將同時受到抑制,這一現象已在ABI-H0731、GLP-26等臨床試驗中得到驗證(表1)。此外,CpAM在較高濃度(為抑制pgRNA包裝濃度的10~34倍)也可以誘導成熟核衣殼的解聚,從而抑制HBV感染及cccDNA的從頭合成[30-32]。但與上述現象相反,在一定情況下誘導細胞內產生的子代成熟核衣殼的解聚甚至可促進cccDNA的細胞內擴增。這可能是由于CpAM誘導子代成熟核衣殼的解聚發生于核孔或核孔附近,從而加速了rcDNA向細胞核內的遞送和cccDNA的合成增加[31]。筆者最近的研究[33]證實核衣殼蛋白的耐藥突變同時抵抗CpAM抑制pgRNA包裝、DNA復制和成熟核衣殼解聚及cccDNA從頭合成。這些結果進一步證明CpAM的這些抗病毒作用皆通過結合于HAP口袋,并通過與相同的氨基酸殘基作用而實現。但截止目前,尚未發現CpAM可以降低HBV感染肝細胞內病毒RNA的水平, 表明其并未干擾核心蛋白對cccDNA轉錄和病毒RNA代謝的調控作用。
4 CpAM抑制HBeAg分泌的作用機理
除了抑制病毒復制之外,Ⅰ型和Ⅱ型CpAM在較高藥物濃度下(為抑制pgRNA包裝/DNA復制濃度的100~500倍)均可降低分泌的HBeAg水平[22,34]。如圖1所示,HBeAg由precore蛋白剪切加工而來。具體來講,precore蛋白(pre-C,p25)除了共享Cp的全部序列(amino acid,aa 1~183)外,p25在其氨基端還獨有一個29個氨基酸殘基序列,位于最前端的19個氨基酸殘基為其分泌信號肽,可與信號肽識別顆粒結合,引導分子量為25 kD的p25蛋白到達內質網膜。在信號肽酶切割信號肽之后,所產生的p22蛋白被轉入內質網腔。當p22蛋白循分泌途徑抵達高爾基體內時,furin蛋白酶切割與Cp共享序列的羧基端結構域,最終形成可分泌的p17蛋白。在p17蛋白的(-10)~(-1)區域,(-7)位半胱氨酸(Cystine,C)與第61位半胱氨酸形成分子內C(-7)-C61二硫鍵,隨后進一步形成同源二聚體被分泌至細胞外,即HBeAg。由于分子內二硫鍵的形成,使得p17二聚體與Cp二聚體結構不同,不能包裝產生核衣殼樣結構[36]。 故與Cp不同,HBeAg以可溶性二聚體形式存在,并被分泌到細胞外。那么,CpAM又是怎樣抑制HBeAg分泌的呢?筆者最近的研究[33]發現CpAM并不影響細胞內p22蛋白的水平,但減少細胞內p17。進一步分析發現,并非所有的p17都形成C(-7)-C61分子內二硫鍵,有40%~50%的p17則以還原形式存在于細胞內。由于還原型p17結構與Cp類似,在環境發生改變時則可能形成Cp樣二聚體,并包裝形成空衣殼[33,37]。此外,HAP_R01可誘導還原型p17組裝產生非核衣殼樣多聚物,某些CpAM耐藥突變,如Cp T33N,也抵抗了CpAM 對HBeAg分泌的抑制作用[34]。因此,筆者推測CpAM可能結合于還原型p17二聚體間的HAP口袋誘導細胞內還原型p17組裝成非核衣殼樣多聚物并隨之降解,從而抑制HBeAg的分泌。當然,這個理論假說還需進一步的結構生物學和細胞生物化學實驗研究的驗證。由于抑制HBeAg分泌需要較高濃度的CpAM,目前為止尚未在臨床試驗中發現其可顯著降低血清中HBeAg的水平。由于HBeAg的免疫抑制作用,研發新一代同時有效抑制HBeAg分泌的靶向Cp/p17的抗病毒藥物,在抑制病毒復制的同時減少HBeAg的分泌,或可同時改善宿主的抗HBV免疫功能,促進慢性乙型肝炎的功能性治愈。
5 CpAM臨床試驗現狀概覽
CpAM為目前處于臨床前和臨床研發階段的熱點新型抗HBV藥物,已有超過10種Ⅰ型和Ⅱ型CpAM進入Ⅰ期臨床試驗,并在概念驗證(proof of concept,POC)臨床試驗中證明其具有明確的抗病毒作用[38](表1)。進入Ⅱ期臨床試驗的CpAM存在單一治療和與其他不同作用機制的新藥或與已獲批抗HBV藥物聯合等模式。下面,將簡述和討論幾個具有代表性的CpAM臨床試驗結果。
NVR3-778是第一個進入臨床試驗的CpAM。在28 d的POC臨床試驗中,該藥單獨或與PEG-IFNα-2a聯用均顯著降低CHB患者血清HBV DNA和HBV RNA水平。且聯合用藥抑制病毒復制強度優于單一用藥[39]。GLS4是Ⅰ型CpAM的代表性藥物,不僅高效干擾核衣殼裝配和解聚,也對核苷類藥物耐藥的HBV有相同的抗病毒活性。該化合物在健康受試者體內單獨使用時盡管安全性和耐受性良好,但單用無法維持有效的血藥濃度。目前GLS4多與利托他韋聯用以提高GLS4的血藥濃度[40]。在為期28 d的POC臨床試驗中,GLS4與利托他韋聯用組患者耐受性良好且血清HBV DNA、HBV RNA下降明顯,同時HBsAg和HBcAg也呈一定幅度的下降。Ⅱ型CpAM AB-506可廣泛抑制A~H基因型HBV的復制,并抑制核苷(酸)類似物(NUC)耐藥突變的HBV復制。在HBV高壓水動力小鼠動物實驗中可使小鼠血清中HBV DNA下降3log10,在與RNAi和NUC聯用時表現出協同作用[41]。在為期28 d的Ⅰ期臨床POC試驗中,AB-506對血清HBV DNA、HBV RNA的抑制最高可達-2.8 log10拷貝/mL,與同期其他CpAM抗病毒水平相當[42]。但由于試驗中觀察到健康受試者人群中發生2例急性肝炎,處于安全性考慮該藥的后續臨床試驗已終止。值得一提的是,1例在治療基線發生Cp I105T自然突變的CHB患者對160 mg/d劑量的AB-506呈現無應答狀態。體外細胞抗病毒試驗進一步證實Cp I105T突變HBV對AB506耐藥[43]。此為迄今為止唯一報道的CpAM原發耐藥的病例。可以預見,隨著CpAM臨床研究的廣泛深入,更多的原發和繼發抗藥突變將被發現。這也為今后CpAM耐藥突變機理的研究,耐藥病毒的監測驗證和開發新型具有高耐藥屏障的CpAM提出了更高的要求。
Ⅱ型CpAM ABI-H0731為近期臨床試驗推進最快的CpAM之一。其安全性、有效性皆在Ⅰ期臨床試驗中得到了積極反饋,對CHB患者血清中HBV DNA、HBV pgRNA呈現出很好的抑制效應[44]。在NUC經治的、已獲得持續病毒學抑制的HBeAg陽性或陰性CHB患者中,ABI-H0731聯用NUC可幫助NUC經治的HBeAg陽性患者血清HBV DNA實現更快地下降、實現pgRNA陰轉,但HBsAg的變化不明顯[45]。在后續臨床試驗中發現,無論在NUC初治人群或經治人群,盡管相比于單獨使用恩替卡韋,聯合使用ABI-H0371與恩替卡韋呈現出更快更顯著的HBV DNA、HBV pgRNA的降低,ALT復常率明顯升高[46]。但在HBeAg陽性的NUC未治CHB人群,ABI-0731與NUC聯用72周時仍無法實現持續性病毒學應答(血清HBV DNA、HBV RNA水平雖持續下降,但仍未低于檢測下限);在HBeAg陽性或陰性的NUC經治患者組,部分患者血清HBV DNA、HBV RNA水平低于檢測下限且HBeAg水平陰轉或≤5 IU/mL時停止治療觀察其病毒學反彈情況,結果顯示所有停止治療的患者在16周內全部病毒學反彈[47]。這些臨床試驗數據表明,現有CpAM可顯著降低CHB患者血清中的HBV DNA和HBV RNA,但并未顯著影響患者血清中病毒抗原的水平。且多數CpAM在Ⅱ期臨床試驗中選擇與現有抗HBV藥物聯合用藥,以監測其是否具有抑制HBV復制的協同作用,但與上述ABI-H0731的Ⅱ期臨床試驗結果類似,CpAM與其他藥物聯用時確實具有更好的協同抗病毒作用,但停藥后全部觀察到病毒學反彈。
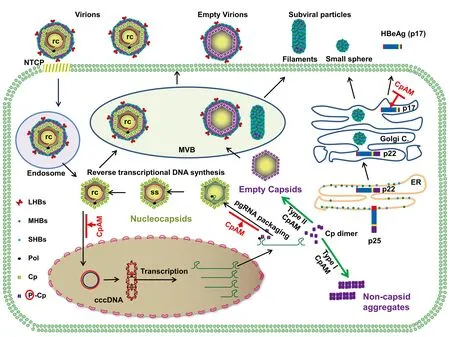
注:Ⅰ型CpAM誘導Cp二聚體形成多種形態的非衣殼結構,并最終經由自噬途徑被降解[35];Ⅱ型CpAM加速衣殼組裝過程,誘導空衣殼結構產生。
6 CpAM對促進CHB患者功能性治愈的臨床應用前景
CpAM因其作用于HBV復制和抗原分泌等多個重要環節,是一類備受期待的新型抗HBV藥物。然而,目前的臨床試驗結果表明,盡管其具有較強的抗病毒活性,能顯著降低患者血清中HBV DNA和RNA,但不顯著改變病毒蛋白水平(表1)。盡管CpAM與NUC聯用可更快更強降低HBV DNA和RNA,但對患者的血清HBsAg水平并無顯著影響,且停止給藥后仍有高的病毒學反彈,無法獲得長時間的病毒學抑制。這些研究結果表明,在絕大多數患者有限療程(1年,甚至3年)的強效抑制HBV DNA復制并不能耗竭或持續沉默cccDNA、或有效激活宿主抗病毒免疫反應,因而無法達到持久控制HBV感染的目的。此外,越來越多的證據表明在病毒復制被抑制后,有效激活宿主特異性抗病毒體液和細胞免疫為CHB功能性治愈所必需。因此,亟需研發可高效抑制cccDNA合成和HBeAg分泌的新型CpAM,并與其他類型抗病毒藥物(siRNA)和免疫治療藥物(PEG-IFNα-2a, TLR7 or TLR8激動劑,治療性疫苗等)聯合或序貫治療,以期加速cccDNA耗竭、降低病毒抗原水平并激活HBV特異性抗病毒免疫,從而控制HBV感染,實現CHB的功能性治愈。
利益沖突聲明:所有作者均聲明不存在利益沖突。
作者貢獻聲明:劉慧、郭巨濤負責擬定文章寫作思路及文章初稿的撰寫;魯鳳明、郭巨濤負責文章內容的修改并最終定稿。
猜你喜歡
保健醫苑(2022年5期)2022-06-10 07:46:38
現代臨床醫學(2022年3期)2022-06-06 07:59:40
昆明醫科大學學報(2022年1期)2022-02-28 07:43:40
哲學評論(2021年2期)2021-08-22 01:53:34
科學大眾(2020年12期)2020-08-13 03:22:22
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
中國當代醫藥(2015年17期)2015-03-01 02:03:58
現代企業(2015年9期)2015-02-28 18:56:50
