葡萄CBF4基因生物信息學及其對低溫和硅酸鉀響應分析
2022-09-07 09:45:56張紅梅王旺田王寶強王翠玲
廣西植物 2022年8期
張紅梅, 王旺田, 張 芮, 楊 科, 王寶強, 王翠玲
( 甘肅農業大學 生命科學技術學院, 甘肅省干旱生境作物學重點實驗室, 蘭州 730070 )
葡萄是世界最古老的落葉果樹之一,各地均有栽培,已成為重要的果樹經濟作物,種植面積和產量居世界首位,是我國的重要果樹。中國北方的葡萄由于低溫凍害使得產量減少,造成巨大的經濟損失。因此,研究葡萄的低溫響應機理,提高葡萄對低溫的適應性是十分必要的。
作為AP2/EREBP轉錄因子群的一個亞科,基因在對非生物脅迫的耐受性中發揮核心作用,植物在低溫脅迫下表現出依賴的應答通路。能夠與啟動子中的核心片段(CCGAC)相結合從而調控該基因的轉錄水平(Haake, 2002)。研究表明擬南芥1、2和3都位于Ⅳ染色體上,且緊密分布于短臂72.8 cM處,它們編碼與AP2/ERF家族密切相關的轉錄因子,這些轉錄因子與調控基因啟動子中存在的CRT/DRE DNA調控元件結合。2基因能夠負調控1 和3,從而調控下游COR等抗寒基因的表達來提高植物抗寒性(Novillo et al., 2004;呂勝男等,2011;董亞茹等,2017)。而4則位于擬南芥的Ⅴ號染色體上,是唯一已知的基因參與脫落酸(ABA)依賴的信號通路(沙麗娜,2009)。這些研究成果說明轉錄因子對植物的抗寒、抗旱和抗鹽堿等脅迫過程發揮著重要作用。研究表明,硅(Si)可以提高水稻(任學坤等,2007)、小麥(鄭世英等,2015)、高粱(劉朋等,2014)等植物的抗逆性,施加外源硅能夠提高低溫脅迫下葡萄葉片滲透調節物質含量,促進蔗糖轉運速率,緩解活性氧積累,增強耐寒性(鄭凱翔等,2019)。
目前,國內基因主要集中于擬南芥(Michael et al., 2010)、大豆(Kidokoro et al., 2014)、玉米(Zhang et al., 2010)、番茄(Yuasa et al., 2014)等植物的研究,對葡萄中基因的抗寒作用等方面研究較少,尤其對4在外源硅與低溫協同作用下的表達未見報道。本研究對4基因編碼的蛋白進行全面的生物信息學分析,對葡萄幼苗進行低溫和硅酸鉀處理,并對4基因進行實時熒光定量分析,以期為4 基因的表達特性及其功能研究奠定理論基礎。
1 材料與方法
1.1 序列來源
利用NCBI數據庫獲取釀酒葡萄基因4(GenBank:DQ497624.1)。
1.2 蛋白質結構預測
1.2.1 一級結構 通過Expasy進行氨基酸殘基的數目和組成以及蛋白質的一級結構在線分析。
1.2.2 二級結構 利用SOPMA進行二級結構的預測;利用ProtScale預測蛋白的親疏水性;利用TMHMM Server v.2.0預測蛋白的跨膜結構;利用SignalP 4.0預測蛋白的信號肽。
1.2.3 磷酸化位點和糖基化位點預測 分別使用KinasePhos和NetOGlyc 4. 0預測蛋白的磷酸化和糖基化位點。
1.2.4 CBF4蛋白的細胞定位 使用PSORT Prediction 預測蛋白的細胞定位。
1.2.5 結構域預測 使用NCBI中的CDD數據庫對蛋白的結構域進行分析。
1.2.6 三級結構 通過 SWISSMODEL對葡萄4基因編碼蛋白的三維結構進行同源建模。
1.3 同源性比對及系統進化樹構建
通過NCBI在線比對,找出與該蛋白同源性較高的其他植物的氨基酸序列,再利用MEGA X軟件進行系統進化樹的構建。
1.4 低溫脅迫對葡萄組織CBF4基因的熒光定量PCR分析
以釀酒葡萄試管苗貝達為試驗材料,當植株長至5片葉時,選取長勢一致的幼苗煉苗3 d后移入裝有1/2強度Haogland營養液的水培盆(10 cm × 10 cm × 9 cm),用泡沫板固定,在人工氣候培養箱中培養(溫度25 ℃;濕度60%;光照強度5 000 lx)60 d,營養液每2 d更換1次。外源硅處理一周后,按實驗設計分別進行常溫(25 ℃)及室內模擬低溫(5 ℃)處理,各處理過程持續光照,對照為不加硅酸鉀的葡萄水培苗,且其他條件相同。低溫處理3 h和9 h后分別取各組幼苗的中部幼嫩葉片和根部,提取RNA,反轉錄得到cDNA,以葡萄管家基因ubiquitin(AY684131.1)為內參基因對葡萄葉片和根部的4基因表達水平進行分析,根據TAKARA SYBR GREEN試劑盒說明書進行qRT-PCR反應液的配制,在定量PCR儀器上進行試驗,所有試樣重復3次,qRT-PCR生成的數據由定量分析軟件讀取,采用2法對所得數據進行分析。根據基因序列設計引物,得到引物序列如下:4(正向:5′-AAGTGGGTATGCGAGGTAAG-3′,反向:5′-TTCTGAATGTCCTTGGCG-3′),退火溫度為60 ℃;ubiquitin(正向:5′-GGCTTGGGAGATGGGAAAC-3′,反向:5′-TCCTACAATACCACCAAACATAGCA-3′),退火溫度為60 ℃。
1.5 數據處理
采用Origin 9和SPSS 22.0 軟件進行數據處理和分析,運用Duncan雙因素方差分析每個處理之間的差異顯著性(α=0.05)。
2 結果與分析
2.1 蛋白質結構分析
2.1.1 一級結構 對CBF4蛋白的理化性質分析得到葡萄CBF4蛋白的分子式為CHNOS;相對分子量為24.22 kD;理論等電點(pI)為5.42;不穩定系數為51.89,總平均親水性為-0.621,表明該蛋白為不穩定的親水性蛋白;脂肪系數為61.83,表明脂溶性比較差;正電荷殘基數(Arg+Lys)和負電荷殘基數(Asp+Glu)分別為27和34。
由表1可知,葡萄CBF4蛋白的20種氨基酸中,丙氨酸(Ala)含量最多(11.0%),其次為精氨酸(Arg)(8.7%)與天冬氨酸(Asp)(8.3%),氨基酸含量最小的是谷氨酸胺(Gln) (0.9%)。其中極性氨基酸占56.9%,非極性氨基酸占43.1%。
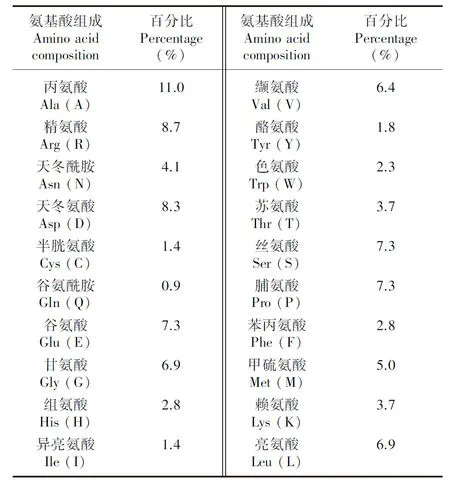
表 1 CBF4蛋白氨基酸組成分析Table 1 Analysis of amino acid compositions of CBF4 protein
2.1.2 二級結構 蛋白二級結構預測(圖1),該蛋白二級結構主要是無規則卷曲,其次是α-螺旋,其中所占比例分別為56.88%和27.52%,延伸鏈占比為13.76%,β-轉角占比最小(1.83%)。
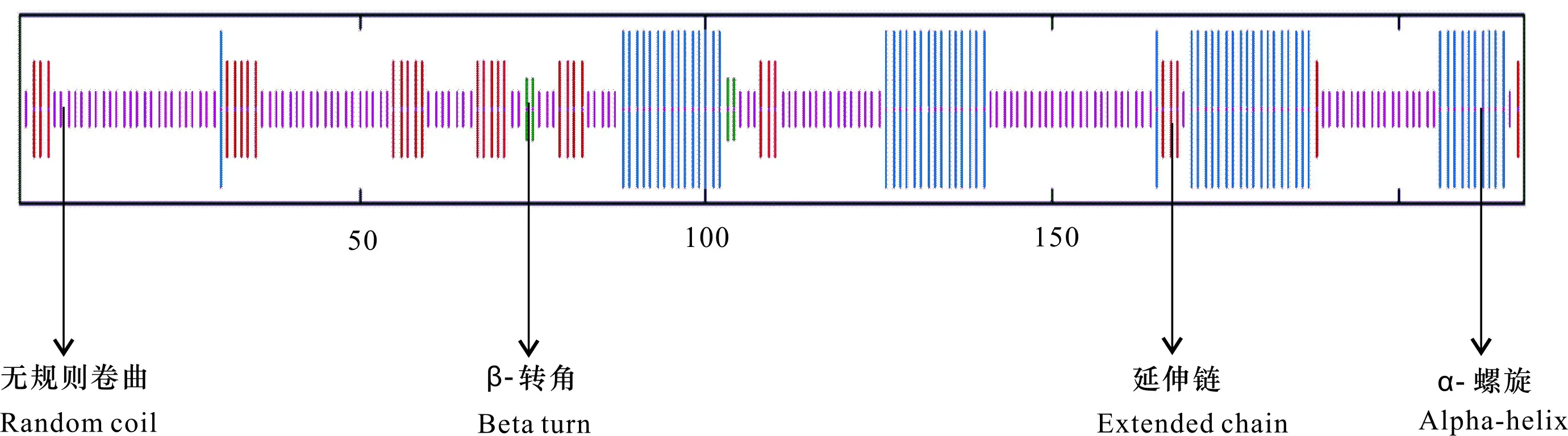
圖 1 CBF4基因編碼蛋白二級結構預測結果Fig. 1 Prediction results of secondary structure of CBF4 gene encoded protein
通過ProtScale分析CBF4蛋白質親/疏水性(圖2),預測結果說明整條多肽鏈沒有明顯的疏水區域。跨膜域的預測結果(圖3)顯示,CBF4蛋白是一個膜外蛋白,沒有發現跨膜螺旋區域,這與沒有明顯疏水區域的預測結果一致。利用SignalP 4.0在線工具預測分析信號肽,對于CBF4蛋白,總結分析得到預測的目的蛋白中不存在信號肽(圖4)。
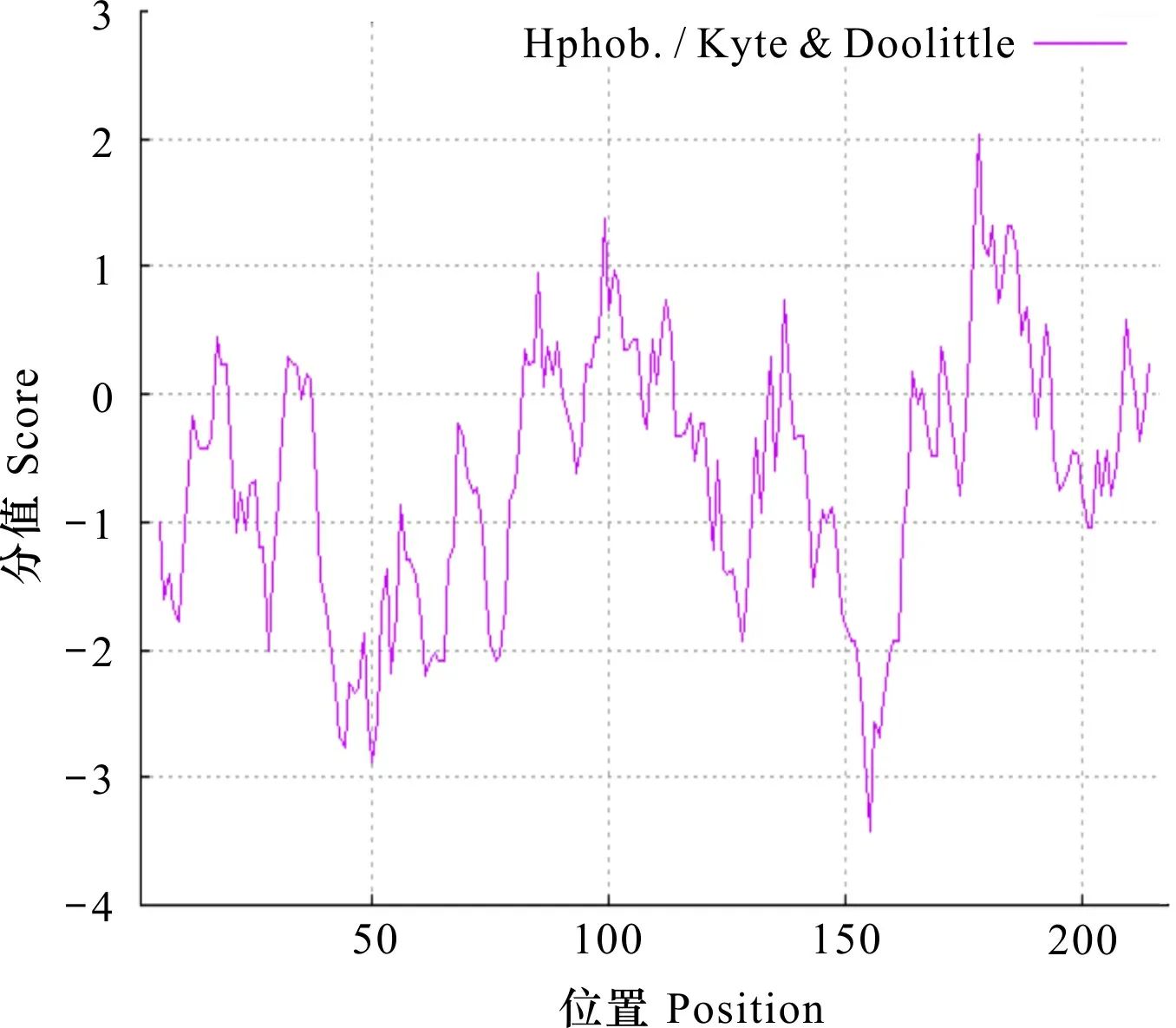
圖 2 CBF4蛋白親疏水性分析Fig. 2 Analysis of affinity and hydrophobicity of CBF4 protein
圖 3 CBF4蛋白質跨膜區結果Fig. 3 Results of CBF4 protein transmembrane region
圖 4 CBF4蛋白信號肽結果Fig. 4 Results of CBF4 protein signal peptides
2.1.3 磷酸化位點和糖基化位點預測 分別通過KinasePhos和NetOGlyc 4.0預測葡萄CBF4蛋白的磷酸化和糖基化位點,表明該蛋白含有5個磷酸化位點(圖5)和14個糖基化位點。

圖 5 磷酸化位點預測結果Fig. 5 Results of phosphorylation site prediction
2.1.4 CBF4蛋白的細胞定位 通過PSORT Prediction預測葡萄CBF4蛋白的細胞定位,結構顯示該蛋白定位在細胞核內,因此可以推斷葡萄4基因主要在細胞核內發揮生物學作用。
2.1.5 結構域預測 應用NCBI中的 CDD 數據庫預測葡萄CBF4蛋白的保守結構域,預測結果(圖6)顯示葡萄CBF4蛋白的氨基酸序列的第57位至第115位為AP2超家族結構域,在進化上非常保守,該結構域對蛋白功能的發揮非常重要。
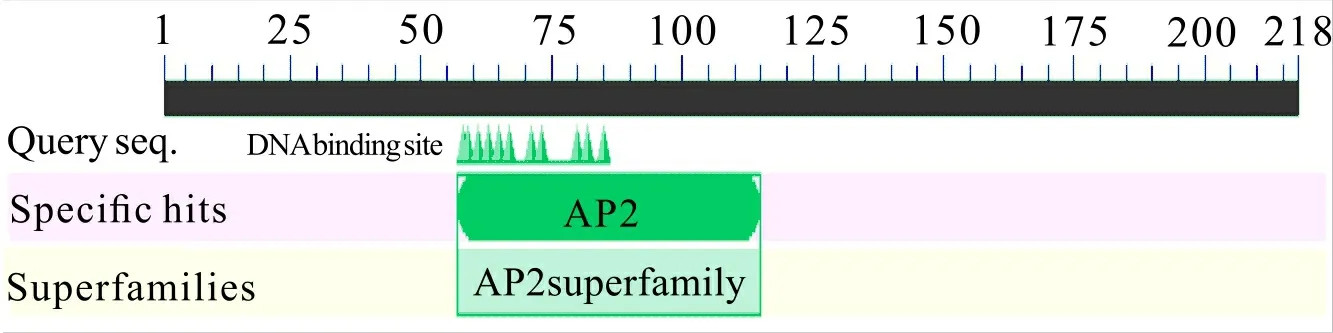
圖 6 結構域預測結果Fig. 6 Results of domain prediction
2.1.6 三級結構預測 通過同源建模方法構建葡萄CBF4基因蛋白的三級結構(圖7),結果顯示該蛋白三級結構主要由α-螺旋和無規則卷曲折疊形成,其預測結果和二級結構相一致。
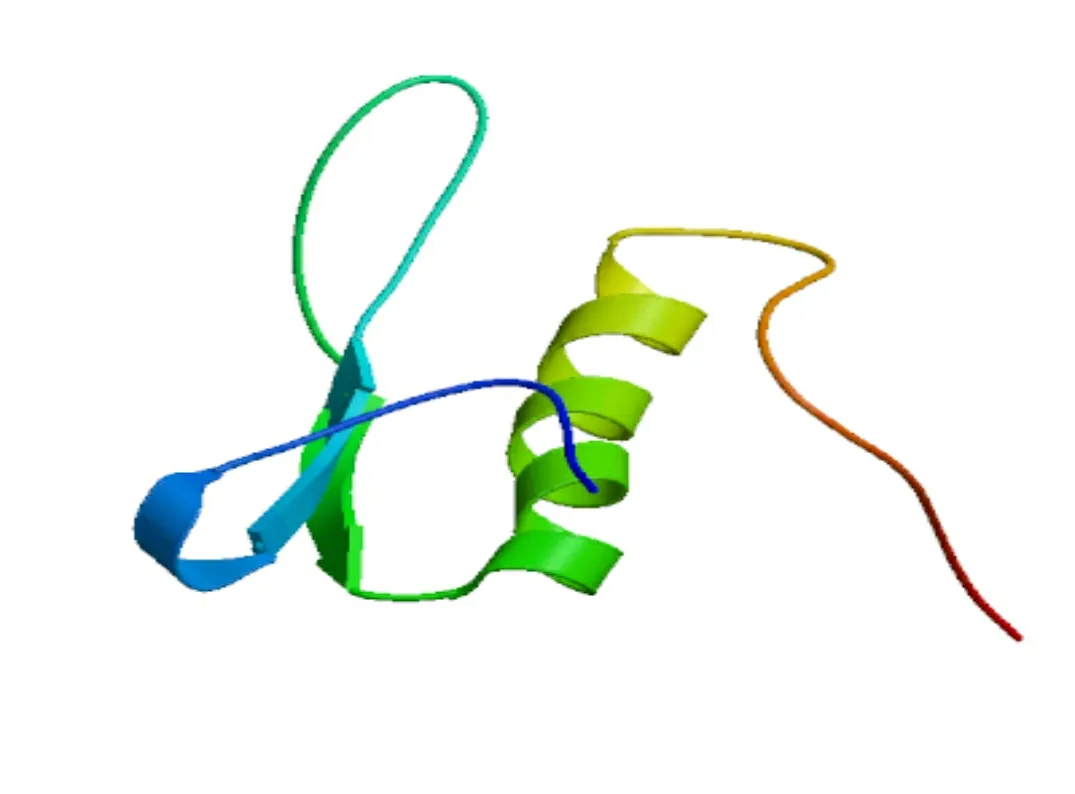
圖 7 蛋白三級結構的預測Fig. 7 Prediction of tertiary structure of protein
2.2 CBF4蛋白進化樹構建及多序列比對分析
將葡萄4基因的序列與同一物種及其他相近物種的序列進行比對分析(圖8),結果顯示釀酒葡萄與美洲葡萄同源性最高,為99.62%,與洋薊、短腳草、爬山虎、獼猴桃、白樺、花生、茶花、赤蘚和咖啡同源性分別為99.64%、92.78%、93.31%、85.71%、81.55%、81.23%、80.13%、78.51%和75.49%。

圖 8 多序列比對結果Fig. 8 Results of multiple sequence alignment
葡萄4基因與13種同物種和相近物種同源序列的系統進化樹如圖9所示,釀酒葡萄與美洲葡萄的關系最密切,其次是河岸葡萄、山葡萄、沙地葡萄、洋薊、爬山虎、短腳草,與獼猴桃、茶花、白樺、花生、赤蘚、咖啡關系較遠,遺傳距離也隨之增加。
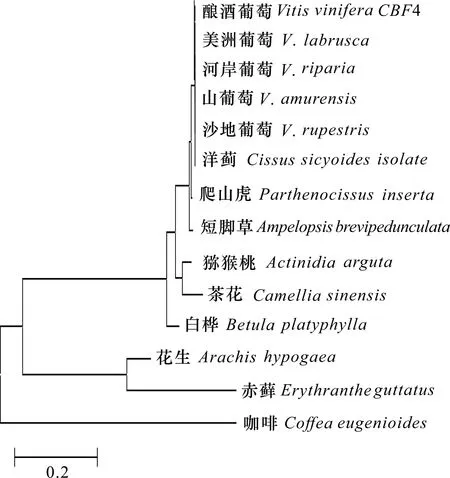
圖 9 系統進化樹構建結果Fig. 9 Results of system evolution tree construction
2.3 葡萄CBF4基因在低溫脅迫下的表達
葡萄水培苗經低溫脅迫處理后,4基因的表達結果顯示,低溫脅迫下葡萄葉片4基因隨低溫處理時間延長相對表達水平顯著上調,低溫脅迫3 h和9 h的上調幅度分別為對照的1.209倍、13.812倍;施加外源硅后常溫條件下4基因表達水平上調但差異不顯著,低溫條件下4基因表達水平下調(圖10:A)。在相同處理時間內,低溫脅迫相對常溫比較,葡萄根部相對表達水平下調,3 h和9 h的下調幅度分別為對照的0.573倍和0.422倍,在常溫及低溫條件下,施硅處理較對照,4基因表達均上調(圖10:B)。
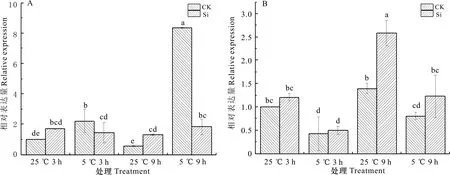
A. 葉片; B. 根部。不同小寫字母代表差異顯著(P<0.05)。A. Leaves; B. Roots. Different small letters indicate significant differences (P<0.05).圖 10 CBF4基因在低溫脅迫下的表達分析Fig. 10 Expression analysis of CBF4 gene under low temperature stress
3 討論與結論
植物受到逆境脅迫會使植物在生長發育、形態建成、物質和能量代謝等方面發生一系列的變化。相比于傳統育種,利用生物信息學能夠快速準確地探索植物抗逆基因資源,克隆相關基因并且利用相關基因提高植物抗逆性(賈翠翠,2015)。釀酒葡萄的種植會受到冷凍、干旱和高鹽的限制,低于-20 ℃的溫度會對葡萄樹造成不可逆轉的損害,影響許多葡萄栽培種的產量,降低種植者收入,若將相關抗性基因克隆并轉入釀酒葡萄中,則可以顯著提高釀酒葡萄的產量。目前,對于轉錄因子的研究較為深入和廣泛,該轉錄因子在植物非生物脅迫方面發揮著重要作用(Feng et al., 2011;Novillo et al., 2012)。研究發現1基因在馬鈴薯中超表達,能夠增強馬鈴薯的抗寒性(Pino et al., 2008)。將歐洲越桔中的1基因在擬南芥中超表達可增強其抗寒性(Oakenfull et al., 2013)。綜上所述,基因參與了植物體低溫脅迫的響應,這與本研究結果保持一致。
通過分析可知,葡萄CBF4蛋白氨基酸組成中極性氨基酸占56.9%,非極性氨基酸占43.1%,CBF4蛋白無信號肽,是一個不穩定的、親水的、脂溶性較差的膜外蛋白,與極性氨基酸所占比例一致。采用SOPMA及SWISS-MODEL分別預測CBF4 蛋白二級結構和三級結構,表明該蛋白主要結構單元是無規則卷曲,其次是α-螺旋,其中所占比例分別為56.88%和27.52%,該結果為研究4基因及其編碼產物的結構和功能提供了更多的信息。CBF4蛋白的多序列和系統進化分析表明,釀酒葡萄與美洲葡萄的同源性最高、親緣關系最近,這種同源性一方面體現出各物種間親緣關系的遠近,另一方面也表明多個不同物種的4基因編碼產物在結構特征中比較穩定,保守性較高。通過對葡萄CBF4蛋白的結構域分析得到,葡萄4包含一個AP2/EREBP結構域,屬于AP2型DNA保守結合大家族中的CBF/DREB家族,具有該家族典型的特征,含有YRG元件和WLG基序,具有高度保守性,該結構域對編碼蛋白功能發揮著極其重要的作用,可調節植物抵御低溫和干旱相關基因的表達,推測4基因與植物逆境脅迫可能密切相關(韓志萍等,2006;邵文靖等,2020),這說明4在葡萄的抗逆性中有著非常重要的作用,具有深入研究的價值。
低溫能夠誘導大多數植物體內的基因表達,如山葡萄在寒冷、鹽度、脫落酸和水楊酸處理下,4轉錄本積累增加(Dong et al., 2013)。在葡萄樹中,4基因通常通過冷處理誘導,低溫條件下葡萄4轉錄水平相對快速地增加,并且可以在葉片中保持很多天(Xiao et al., 2010)。本研究分析表明,發現4基因低溫脅迫3 h和9 h在葡萄葉片中表達水平顯著上調,可能是因為轉錄激活因子與CRT/DRE調控元件特異性結合,激活啟動子中目的基因表達,說明4基因可能參與了葡萄葉片響應外界冷脅迫的信號途徑,因此推測出4基因可能參與了葡萄葉片低溫脅迫的響應。低溫條件下施加硅酸鉀, 在葡萄葉片中的表達下調,而在葡萄根部表達上調,可能是因為葡萄根部對低溫和硅酸鉀交互作用比較敏感,揭示了該基因在不同的葡萄組織中對硅酸鉀的響應機制可能不同。目前,應對低溫傷害的方法中使用外源物質提高作物抗性更為簡單,并且效果顯著,為了防止葡萄受到低溫傷害,應該在根部追施硅肥,本研究進一步為施用外源硅提高葡萄抗寒性提供理論依據。
本研究對4基因編碼的蛋白進行全面的生物信息學及低溫和硅酸鉀響應分析,結果顯示低溫脅迫后4基因在葡萄葉片中表達水平上調,說明4基因可能參與了葡萄葉片低溫脅迫的響應。低溫條件下施加硅酸鉀后,4基因在不同的葡萄組織中對硅酸鉀的響應不同,說明該基因表達具有組織特異性。本研究對深入了解4基因在葡萄非生物逆境脅迫中的功能,分析CBF4蛋白對植物抗逆的分子機制,以及深入探究外源硅對葡萄抗寒調節機理,為甘肅乃至北方釀酒葡萄抗寒栽培提供理論基礎和實踐指導。
猜你喜歡
哲學評論(2021年2期)2021-08-22 01:53:34
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
現代企業(2015年9期)2015-02-28 18:56:50
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
終身教育研究(2014年5期)2014-02-28 01:23:06
