乳酸菌產苯乳酸對指狀青霉的抑菌活性及作用機理
2022-09-01 02:32:38郭宇逍鄧麗莉曾凱芳
食品科學 2022年15期
關鍵詞:質量
郭宇逍,洪 陽,鄧麗莉,2,曾凱芳,2,*
(1.西南大學食品科學學院,重慶 400715;2.西南大學食品貯藏與物流研究中心,重慶 400715)
柑橘是我國重要的經濟作物之一,具有豐富的營養價值和藥用價值。柑橘果實在采摘、包裝、貯存和運輸的整個過程中都可能會受微生物侵染而發生腐爛。其中,由指狀青霉()引起的綠霉病是柑橘采后主要病害之一,在干旱地區和亞熱帶氣候區造成了嚴重的經濟損失,約占柑橘采后損失總量的90%。目前已有的防控方法中,多菌靈和咪唑類等化學殺菌劑使用最廣泛也最有效,但化學殺菌劑會引起環境污染、損害人體健康、使病原菌產生抗性等一系類問題,因此,亟待尋求一種綠色安全無污染的處理方式來減少或替代化學殺菌劑的使用。
乳酸菌在食品體系中作為生物保護菌所具有的抗真菌特性能夠有效防治草莓、蘋果、圣女果等果蔬采后真菌病害,其重要機制之一是能產生有機酸,主要包括乳酸、乙酸、苯乳酸(phenyllactic acid,PLA),此外還包括羥基苯乳酸和吲哚-3-乳酸等。有機酸分子在較低的pH值環境下具有抑菌活性,且具有親脂性,可穿過細胞質膜進入細胞發生解離并釋放出質子和陰離子,從而破壞膜的質子動力,抑制細胞的正常代謝活動,導致ATP產量下降。有機酸對病原菌的抑制作用還可能包括對酶活性的抑制,以及有機酸的非解離組分對病原菌細胞膜的破壞。
PLA作為乳酸菌產生的有機酸之一,可通過乳酸菌高效發酵的方式低成本大量獲得,且具有廣譜抑菌性,不僅能抑制腸道沙門氏菌、大腸桿菌、金黃色葡萄球菌等多種細菌生長,對青霉、黃曲霉、黑曲霉等真菌同樣具有良好的抑制效果。前人研究表明,PLA主要作用于細胞膜。Liu Fang等分析發現,PLA處理改變了細胞形態,且破壞了浮游細胞細胞膜完整性,導致細胞發生不同程度的ATP泄漏并嚴重受損。類似的,Ning Yawei等通過流式細胞儀和透射電子顯微鏡分析,證明PLA處理能破壞蠟樣芽孢桿菌細胞膜的完整性和形態。但也有研究認為,PLA還能夠作用于細胞壁、DNA。PLA對不同病原菌的抑制效果及抑制機理不同,且目前鮮見深入研究其抗病機制的報道。
因此,本實驗選擇本課題組前期在泡菜中分離篩選得到的兩株乳酸菌CKXP13和CWXP24作為實驗菌株,這兩株菌對柑橘綠霉病都有良好的控制效果。但有關乳酸菌CKXP13和CWXP24所產生的抑菌活性物質研究尚少,也鮮有研究其抑菌活性物質產量及抑菌物質作用機理的相關報道。綜上所述,本實驗旨在探究乳酸菌CKXP13和CWXP24的產酸能力和PLA的產量,并通過研究PLA對離體指狀青霉孢子的影響初步揭示PLA的抑菌機理,同時驗證PLA對柑橘綠霉病的防治效果,以期為控制柑橘采后綠霉病和抑制柑橘采后病原菌提供一定的理論依據。
1 材料與方法
1.1 材料與試劑
豐臍橙(L. Osbeck)摘自重慶市北碚區。挑選無機械傷、無病害、成熟度和大小基本一致的果實,預冷后放至4 ℃冷庫中備用。
指狀青霉()為本實驗室保藏,于馬鈴薯葡萄糖瓊脂(potato dextrose agar,PDA)培養基中25 ℃培養7 d,得到孢子懸浮液后用無菌蒸餾水調至所需濃度。CKXP13和CWXP24為本實驗室前期實驗分離保藏。色譜純PLA上海阿拉丁生化科技有限公司。
1.2 儀器與設備
PB-10酸度計 德國賽多利斯股份有限公司;JSM-6510LV掃描電子顯微鏡 日本JEOL公司;SYNERGYH1MG全自動酶標儀 美國Bio-Tek公司;LC-20A高效液相色譜 日本島津公司;DDS-307A電導率儀 上海INESA公司。
1.3 方法
1.3.1活化、發酵及產酸能力的測定
乳酸菌活化/乳酸菌懸浮液制備:從-40 ℃冰箱中取出已篩選鑒定的兩株乳酸菌CKXP13和CWXP24,按1%(以體系體積計,下同)的接種量加入20 mL MRS肉湯中,37 ℃恒溫培養24 h,活化2 次后待用。
乳酸菌無菌發酵液制備:以1%的接種量接種乳酸菌培養液于MRS肉湯中,37 ℃靜置培養48 h。每株菌的培養液于4 ℃條件下5 000 r/min離心15 min,上清液用0.22 μm孔徑水系濾頭過濾除菌,4 ℃冰箱保存備用。
產酸能力的測定參照Ma Jiahong等的方法并作一定的修改,同時測定生長曲線。取活化后的兩株乳酸菌培養液各1 mL加入至100 mL MRS肉湯中,37 ℃靜置培養72 h,每6 h測定pH值與OD各一次,重復3 次。
1.3.2 高效液相色譜法測定無菌發酵液中PLA質量濃度
液相色譜條件:選用LC-20A高效液相色譜儀、液相色譜C柱(150 mm×4.6 mm,5 μm),Prominence SPD-M20A二極管陣列檢測器。流動相A:體積分數0.05%三氟乙酸-水溶液,流動相B:體積分數0.05%三氟乙酸-甲醇溶液。流速1 mL/min、柱溫30 ℃、檢測波長210 nm、進樣量10 μL。
梯度洗脫程序:0~20 min由10% B線性變化至100% B,20~23 min保持100% B,23~25 min由100% B線性變化至10% B,25~40 min保持10% B。
1.3.3孢子存活率的測定
在無菌蒸餾水中加入PLA,使其終質量濃度分別為2.5、5.0、10.0 mg/mL,再加入不同濃度的孢子懸浮液,使其終濃度為1×10個/mL,25 ℃環境下靜置16 h,取50 μL混合液于PDA培養基上涂布培養2 d,記錄培養基上的菌落總數。以無菌蒸餾水替代PLA作為對照,重復3 次。按式(1)計算孢子存活率。

1.3.4菌落直徑的測定
在PDA培養基中加入適量PLA,使其終質量濃度分別為2.5、5.0、10.0 mg/mL,再接種5 μL孢子懸浮液(1×10個/mL),25 ℃環境下培養8 d,每天觀察并測量菌落直徑。以無菌蒸餾水替代PLA為對照,重復3 次。
1.3.5菌絲表面形態的觀察
菌絲表面形態的觀察參照Tao Nengguo等的方法并稍作調整,將孢子懸浮液(1×10個/mL)接種于PDB液體培養基中,25 ℃、160 r/min培養48 h,5 000 r/min離心15 min,無菌蒸餾水洗3 次,收集菌絲體,用不同質量濃度的PLA溶液(2.5、5.0、10.0 mg/mL)處理48 h,以無菌蒸餾水代替PLA作為對照。處理結束后,收集菌絲體,置于體積分數2.5%戊二醛溶液(含0.1 mol/L磷酸緩沖液,pH 6.8)中,存放于4 ℃冰箱中過夜固定。固定結束后除去固定液,用磷酸緩沖液(0.1 mol/L、pH 6.8)清洗3 次,再用乙醇梯度脫水,分別用體積分數30%、50%、70%、90%、95%乙醇溶液脫水處理一次,每次10 min;用無水乙醇脫水處理3 次,每次10 min。脫水處理之后,依次加入體積分數50%、70%、90%、95%叔丁醇溶液進行置換處理一次,每次10 min;用無水叔丁醇置換處理兩次,每次10 min。最后,將樣品放入恒溫干燥箱65 ℃過夜干燥,挑取樣品組織塊粘到金屬臺上,進行噴金、鍍膜后在掃描電子顯微鏡(放大倍數5 000 倍、80 kV)下觀察并拍照。
1.3.6菌絲胞外電導率的測定
菌絲胞外電導率的測定參照Wang Wenjun等的方法并稍作調整,將10 mL含有孢子懸浮液(1×10個/mL)的PDB培養基置于25 ℃、160 r/min條件下振蕩培養48 h。培養結束后5 000 r/min離心15 min收集菌絲體,無菌蒸餾水洗3 次,重懸于無菌蒸餾水中。取適量PLA溶液加入菌絲懸浮液中,使懸浮液中PLA終質量濃度分別為2.5、5.0、10.0 mg/mL。在處理0、3、6、9、12、24、48 h時測定胞外電導率,每組處理重復3 次,以無菌蒸餾水替代PLA作為對照。
1.3.7 柑橘果實綠霉病干預實驗
柑橘果實綠霉病干預實驗參照Ma Jiahong等的方法并稍作調整,柑橘果實先用體積分數2%的次氯酸鈉溶液浸泡2 min,用清水沖洗后于室溫下自然晾干。所有果實隨機分為3 組,打孔接種前用乙醇擦拭消毒。用無菌打孔器在果實赤道部位對稱打兩個孔(直徑3 mm、深3 mm),每個孔接種10 μL孢子懸浮液(1×10個/mL)。待果實吸收孢子懸浮液后,每個孔接種20 μL不同質量濃度PLA溶液(10、20 mg/mL)并置于室溫條件下。無菌蒸餾水替代PLA作為對照,每組處理10 個果實,每個處理重復3 次。待處理液充分吸收后,將果實單果包裝,貯藏于25 ℃、相對濕度95%的環境中。每天統計果實發病情況,發病率和病斑直徑分別按式(2)和式(3)計算。

1.4 數據處理與分析
運用Excel 2016軟件統計分析所有得到的數據,應用GraphPad Prism 8軟件、Adobe Photoshop CS6軟件繪制圖表;運用SPSS 21.0軟件對數據進行方差分析,利用Duncan’s多重比較進行差異顯著性分析,<0.05表示差異顯著。
2 結果與分析
2.1 L. plantarum產酸能力分析結果
如圖1所示,隨著培養時間的延長,兩株乳酸菌數量均逐漸增加,有機酸產物不斷積累,導致培養基pH值不斷降低,并在72 h培養結束時均穩定在4左右。生長曲線結果表明,兩株在培養6 h后均進入生長對數期,相應地,培養基pH值急劇下降,CKXP13在培養36 h后達到穩定期,CWXP24在培養42 h后達到穩定期,菌株達到穩定期后,pH值變化逐漸平緩,并最終趨于穩定。兩株均具備較強的產酸能力,且有機酸產量與兩株的生長趨勢一致。

圖1 兩株乳酸菌發酵液pH值變化及生長曲線Fig. 1 pH change of cultured broth and growth curves of two strains of L. plantarum
2.2 L. plantarum無菌發酵液中PLA質量濃度
如表1所示,根據標準品的高效液相色譜結果得到標準曲線方程為=25 720+37 660,=0.998 1,再計算出無菌發酵液中的PLA質量濃度。結果表明,兩株的無菌發酵液中均有PLA產生,且CWXP24顯示出較高的PLA合成能力,產量可達91.9 mg/L,CKXP13的PLA合成能力相對較弱,為42.4 mg/L。
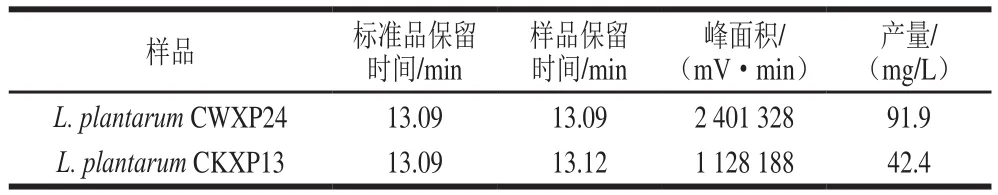
表1 兩株L. plantarum無菌發酵液中PLA的產量Table 1 Concentration of PLA in cell-free culture supernatants of two strains of L. plantarum
2.3 PLA對P. digitatum的體外抑菌機理
2.3.1 PLA對孢子存活率的影響
如圖2所示,PLA在2.5~10.0 mg/mL范圍內均表現出對孢子較強的致死能力,且隨著質量濃度的增大,對的致死能力也不斷增強。2.5 mg/mL及5.0 mg/mL PLA處理組的孢子存活率分別為對照組的43.6%和18.9%,當PLA質量濃度達到10 mg/mL時,平板上已無孢子萌發,孢子存活率幾乎降低至0。
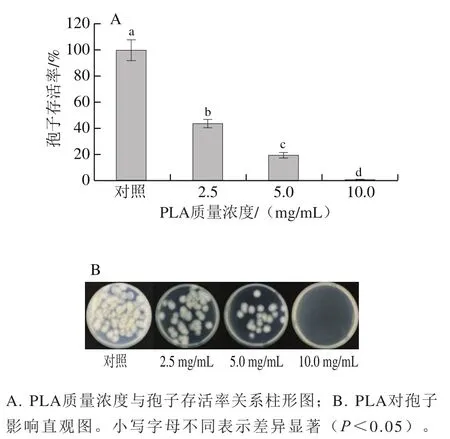
圖2 PLA對P. digitatum孢子存活率的影響Fig. 2 Effects of PLA on spore viability of P. digitatum in vitro
2.3.2 PLA對菌絲生長的影響
如圖3所示,在菌絲的整個生長過程中,不同質量濃度PLA均表現出對菌落生長較強的抑制能力,且隨著質量濃度的增大,其對菌落生長的抑制能力也不斷增強。當PLA質量濃度達到10.0 mg/mL時,菌絲生長完全被抑制。2.5 mg/mL及5.0 mg/mL處理組的菌落直徑在培養8 d后分別為對照組的66.2%和28.5%。
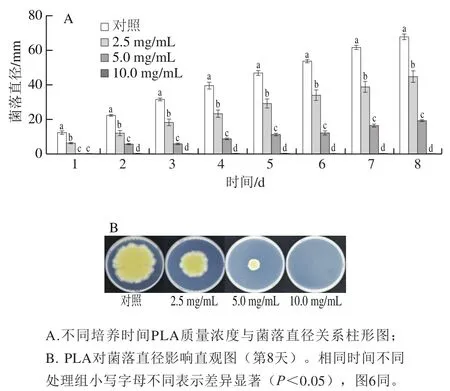
圖3 PLA對P. digitatum菌絲生長的影響Fig. 3 Effect of PLA on mycelial growth of P. digitatum in vitro
2.3.3 PLA對菌絲形態的影響
如圖4A所示,對照組的菌絲分布均勻,生長正常,菌絲表面平整光滑。2.5 mg/mL PLA處理組菌絲結構發生明顯變化,菌絲整體分布不均勻,表面變得粗糙,出現褶皺(圖4B)。高質量濃度的PLA處理組形態變化更加明顯,菌絲整體分布更加不規則,并相互纏繞、扭曲,表面褶皺更加明顯,出現塌陷和樹瘤狀凸起的情況(圖4C、D)。由此推測PLA可對細胞膜結構造成破壞,從而使內容物泄漏。
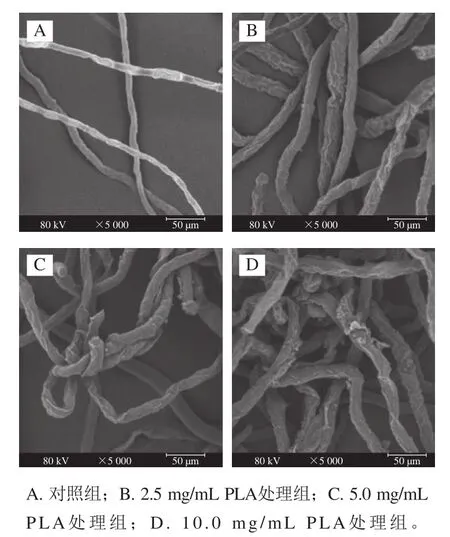
圖4 不同質量濃度PLA處理對P. digitatum菌絲形態的影響(×5 000)Fig. 4 Effect of PLA on mycelial morphology of P. digitatum (× 5 000)
2.3.4 PLA對菌絲胞外電導率的影響
如圖5所示,隨著處理時間的延長,所有組別的電導率均逐漸升高。10.0 mg/mL PLA處理3 h時,其電導率急劇上升,達到71.0 μS/cm,明顯高于對照組(27.0 μS/cm)和其他處理組。2.5 mg/mL PLA處理組在前6 h與對照組差異不明顯,5.0、10 mg/ mL PLA處理48 h時,其電導率分別達到74.3、154.0 μS/cm,均明顯高于對照組(57.3 μS/cm)。電導率分析結果進一步證明PLA處理可破壞菌絲細胞膜的通透性,細胞內容物向外釋放,導致胞外電導率升高。
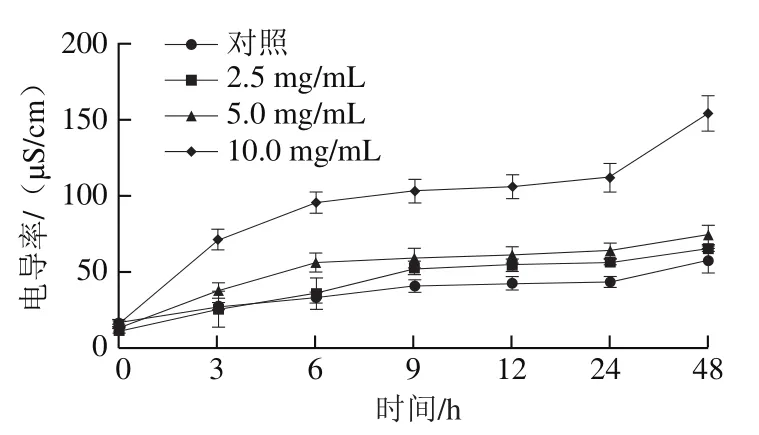
圖5 PLA對P. digitatum菌絲胞外電導率的影響Fig. 5 Extracellular conductivity of P. digitatum mycelia treated with PLA
2.4 PLA對柑橘果實綠霉病的控制效果
如圖6所示,10 mg/mL和20 mg/mL PLA處理均可有效控制對柑橘果實的侵染,接種后5 d內能夠不同程度地抑制柑橘果實綠霉病的發生。接種后5 d,20 mg/mL PLA處理顯著抑制了柑橘果實病斑直徑增長,且兩處理組均有效控制了柑橘果實發病初期的發病率。相對于對照組,20 mg/mL PLA處理組接種后第3天病斑直徑與發病率分別顯著降低了90.4%與78.0%;接種后第5天病斑直徑顯著降低了33.3%。10 mg/mL PLA處理組接種后第3天病斑直徑與發病率分別顯著降低了57.8%與49.9%;接種后第5天病斑直徑降低了15.2%。
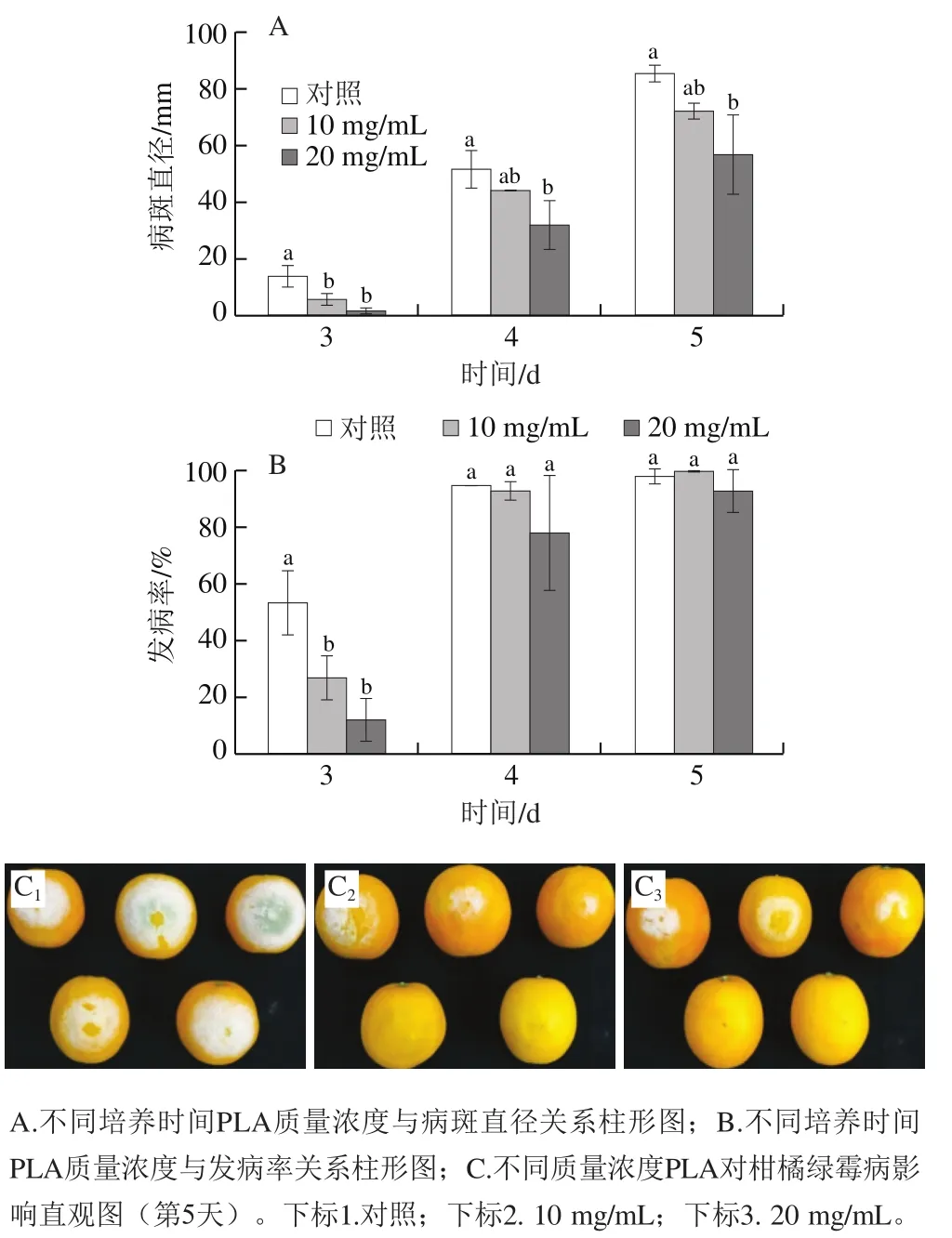
圖6 PLA對柑橘采后綠霉病病斑直徑和發病率的影響Fig. 6 Effect of PLA on lesion diameter and disease incidence of citrus fruit infected by P. digitatum
3 討 論
PLA作為具有抑菌活性的天然物質,可利用乳酸菌以高效發酵的方式進行低成本的自然生產,在果蔬采后貯藏領域顯示出極大的應用潛力。本研究證明了CKXP13和CWXP24均能產PLA,且PLA能有效抑制指狀青霉生長,控制柑橘果實綠霉病。
PLA主要由化學合成法和生物合成法兩種方法制備。相比于化學合成法,生物合成法成本低、操作簡單、條件溫和、對環境友好,因而受到廣泛關注。其中乳酸菌中已有多個菌屬被發現可產PLA,但大多乳酸菌的PLA合成量較低。本實驗中兩株均能分泌產生PLA,其中CWXP24在沒有任何優化培養的條件下產量高達91.9 mg/L,但CKXP13產量較低,僅為42.4 mg/L,以往研究也表明,同一菌種不同菌株間的PLA產量可能存在顯著差異。
本實驗發現PLA質量濃度對孢子的致死效果和對菌絲生長的抑制效果呈正相關,類似地,Guimar?es等發現,PLA質量濃度在0.1~8.0 mg/mL范圍內,的菌落直徑隨PLA質量濃度升高而減小,當PLA質量濃度升至8 mg/mL時,菌落直徑減小為對照的50%。本實驗還對PLA可能的抑菌機理進行了初步研究,證明PLA處理對菌絲表面結構造成了嚴重破壞,使其發生明顯改變,這一結果與前人研究PLA處理對李斯特菌的影響所得結論相似,未使用PLA處理的李斯特菌細胞分布均勻、表面平滑,0.625 mg/mL的PLA處理后細胞外表面呈不規則褶皺,PLA質量濃度增至1.25 mg/mL時細胞膜上出現孔洞或局部破裂,部分胞內物質從細胞膜滲出,形成聚集和黏附,當PLA處理質量濃度持續升高至2.5 mg/mL,細胞損傷更為嚴重,大部分細胞表面坍塌甚至破碎成細胞碎片,進一步說明菌絲的這些變化可能是由于細胞通透性的增加,進而導致小分子物質和離子的泄漏、細胞代謝下降,最終抑制病原菌的生長。本實驗也發現PLA處理質量濃度越高,菌絲的胞外電導率增加幅度越明顯,證明PLA能夠增加菌絲細胞膜的通透性,使細胞內容物發生泄漏,這可能是PLA抑菌機理的重要組成部分。細胞膜作為病原菌細胞能量運輸及物質傳遞的屏障,既能夠保證細胞本身代謝穩定,又能起到調控物質在細胞內外進出與交換的作用。PLA本身為兩親性小分子,且帶有正電荷,PLA的化學結構決定了其更容易吸附在細胞膜表面,與細胞膜發生作用,所以細胞膜一般作為PLA與病原菌作用的第一個靶點。但袁景環等發現,PLA處理的金黃色葡萄球菌和熒光假單胞菌培養液OD值和菌落總數均下降,說明PLA除了具有殺菌作用,還具有溶菌作用,通過掃描電子顯微鏡和透射電子顯微鏡觀察還發現PLA處理后的菌體嚴重變形、固縮或破裂,細胞內容物外泄,推測PLA對這兩種細菌的作用位點之一為細胞壁。這與Dieuleveux等報道的PLA對單核細胞增生李斯特菌的抑菌機理類似,但也可能存在其他作用靶位,因此未來還需對PLA的抑菌機制進一步研究。此外,本實驗對PLA控制柑橘果實采后綠霉病的效果進行驗證,發現20 mg/mL乳酸處理可顯著減小柑橘果實的病斑直徑,且有效控制了柑橘果實的初期發病率,這一結果表明PLA對柑橘果實采后綠霉病具有一定的抑制作用。
綜上所述,本實驗所選用兩株乳酸菌均能產PLA,可為產PLA的乳酸菌的開發和利用提供理論參考,且PLA作為一種新型綠色保鮮劑,應用于柑橘果實采后保鮮具有一定的經濟價值。
猜你喜歡
中學生數理化·中考版(2022年10期)2022-11-10 09:37:42
中學生數理化·八年級物理人教版(2022年12期)2022-02-14 07:08:42
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
中國生殖健康(2019年2期)2019-08-23 08:12:08
石油化工建設(2018年6期)2018-04-22 03:16:54
產品可靠性報告(2017年7期)2017-09-05 09:49:12
中學生數理化·八年級物理人教版(2017年12期)2017-04-18 12:59:38
汽車觀察(2016年3期)2016-02-28 13:16:26
民生周刊(2014年7期)2014-03-28 01:30:54
