內蒙古圍封草原枯落物累積對大針茅和羊草生長發育的影響
2022-08-31 06:08:08侯東杰喬鮮果
生態學報 2022年15期
侯東杰,喬鮮果,郭 柯,*
1 內蒙古農業大學,草原與資源環境學院,呼和浩特 010019 2 中國科學院植物研究所,植被與環境變化國家重點實驗室,北京 100093
內蒙古草原生態系統是中國北方地區重要的生態安全屏障,具有重要的生態功能和經濟價值[1],同時也是中國草原生態系統的主要組成部分。由于氣候變化和人類活動的加劇,內蒙古草原生態系統出現了不同程度的退化,導致其生態服務功能和經濟價值顯著降低[2]。“退牧還草”和“天然草原保護”等國家級生態修復工程先后在內蒙古草原地區實施。圍封作為生態修復工程中常用的措施,具有低成本、高效且易操作的特點[2—4]。圍封不僅顯著提高土壤全碳、全氮、全磷等養分含量[5—7],同時也提高植物群落高度、蓋度、地上生物量、物種豐富度等特征[8—10],使得植被和土壤得到有效的恢復。然而,近些年有研究表明長期連續的圍封會引起草原生態系統枯落物的過量累積,顯著改變群落微環境特征和群落結構與功能[11—14]。
植物種群的生長發育與數量特征是草原生態學研究中的熱點,也是進一步揭示草原植物群落形成、維持與演替過程的關鍵環節[15]。草原圍封后枯落物在地表處的過量累積形成了物理阻隔層,顯著改變群落光照、熱量和水分的分配格局,影響土壤溫度和土壤水分[14,16—17]。有研究表明生長季內枯落物累積對土壤溫度和土壤水分的影響逐漸減弱[14]。植物的物候和生長發育等生理過程對土壤溫度和水分的變化具有極強的敏感性。因此,枯落物累積將通過影響土壤溫度和水分間接對植物種群的生長發育產生影響。植物種群對枯落物累積具有不同的敏感性并通過植株、莖和葉等功能性狀的調整適應圍封草原新的微環境[16,18]。此過程也將對植物種群的生長發育過程產生強烈的影響。然而,圍封草原群落微環境的變化和枯落物累積所造成的機械阻礙作用將如何影響植物種群的生長發育過程,目前缺乏準確的認知。與此同時,也有研究發現草原圍封后群落物種的優勢度排序發生顯著改變,這種現象在內蒙古、寧夏和新疆等地區的草原中表現尤為典型[19—21]。然而引起這種現象產生的驅動機制目前僅從群落微環境的角度有過報道[14]。闡明枯落物累積對不同植物種群的生長發育的影響也有利于從植物種群的角度揭示圍封草原群落結構與功能改變的驅動機制。
典型草原是內蒙古草原中分布范圍廣泛且具有代表性的草原類型[22],也是內蒙古地區實施圍封措施主要的區域。大針茅(Stipagrandis)和羊草(Leymuschinensis)是內蒙古典型草原中廣泛分布的物種,也是群落中的優勢種或亞優勢種,對維持群落結構與功能具有重要作用。由于大針茅和羊草在生物學特性和生態適應性等方面具有截然不同的特性[23—24],這兩種植物成為探究圍封草原枯落物累積對植物種群生長發育過程影響的理想材料。因此,本研究以內蒙古圍封典型草原為對象,探究大針茅和羊草種群的生長發育過程對枯落物累積的動態響應特征,以期從植物種群的角度為闡明圍封草原群落結構與功能改變提供新思路和新視角,也為科學合理制定圍封政策提供理論依據和數據支持。
1 材料與方法
1.1 研究區概況
研究區位于內蒙古錫林浩特市白音錫勒牧場內的中國科學院內蒙古草原生態系統研究定位站(43°33′37″N,116°40′12″E;1244 m)。該區域氣候類型為溫帶大陸性季風氣候,年平均氣溫為0.3℃,最冷月平均氣溫為-21.6℃,最熱月平均氣溫為19.0℃;年平均降水量為351.0 mm,其中80%的降水集中在生長季。研究區植被類型為大針茅(Stipagrandis)典型草原,主要伴生種包括:羊草(Leymuschinensis)、糙隱子草(Cleistogenessquarrosa)、羽茅(Achnatherumsibiricum)、冰草(Agropyroncristatum)、冷蒿(Artemisiafrigida)、菊葉委陵菜(Potentillatanacetifolia)、山韭(Alliumsenescens)和砂韭(Alliumbidentatum)等。依照中國土壤分類系統劃分,研究區土壤類型為栗鈣土。該區域于2009年開始圍封,到2014年為止,共圍封5年,植被得到了有效的恢復且枯落物在群落地表大量堆積。
1.2 實驗設計
實驗采用隨機區組設計,2014年生長季結束后,在圍封的大針茅草原中選取地勢開闊、平坦且植被均勻的位置分別建立3個獨立的實驗區組(22 m×50 m)。將每個區組分成兩個小區(10 m×50 m)并隨機設置枯落物累積小區和枯落物去除小區。各區組內兩小區間留有2 m的緩沖區。在2014—2016年生長季結束后,將枯落物去除小區地表處的枯落物(地表上枯死的、未分解或部分分解的植物莖、葉、花和果實的殘體)全部去除;2015—2017年對枯落物累積小區中的枯落物不做任何處理。
1.3 樣品采集
使用EM50數據采集器和ECH2O 5TE傳感器(METER公司,美國)對兩種處理不同深度的土壤溫度和土壤水分進行連續、原位監測。在樣地中隨機選取一個區組,隨后將ECH2O 5TE傳感器安裝在地勢開闊、平坦的位置,其中ECH2O 5TE傳感器分別安裝在土壤深度2.5 cm和12.5 cm處,以表示枯落物影響下土壤表層和深層的土壤溫度和土壤水分。隨后將土壤回填并將土壤表面修理平整。土壤溫度和土壤水分的測量時間間隔為1 h,土壤溫度和土壤水分的監測從2015年5月1日開始到2017年9月1日結束。
在各小區中選取大針茅(優勢種)和羊草(亞優勢種)作為研究對象,利用定株觀測法研究植物種群的生長發育過程。在2015—2017年生長季開始前,在兩種處理下的各小區中隨機設置5個1 m×1 m的固定樣方,在每個樣方內隨機選取3株大針茅和羊草,并使用插地牌標記。從2015—2017年的4月10日開始,至9月2日結束,記錄標記植株個體的生長動態,觀測期內每隔5—7天對標記的植物個體的營養高度進行測量。每種處理下各物種共45次重復(3株/樣方/小區×5個樣方/小區×3個小區)
2015—2017年的8月1日,使用1 m×1 m的樣方法對兩種處理下的各小區的植物種群數量特征進行調查。首先記錄樣方的物種組成和總蓋度,隨后對各物種隨機選取3株測量其營養高度和生殖高度,再記錄各物種的分蓋度和密度,最后使用齊地面刈割法收獲各物種的地上部分并裝入信封袋。總蓋度和分蓋度使用目測法進行測量,每個小區設置3次重復。將植物樣品帶回實驗室后置于烘箱內,在65℃的條件下烘至恒重并稱重。
1.4 數據分析
使用5月—8月土壤溫度和土壤水分的月平均值表示生長季土壤溫度和土壤水分動態。使用相同處理下各物種營養高度的平均值表示植物生長狀態。本研究中植物的返青時間以植物第一葉片萌發時間為依據;植物從返青到最大營養高度的時間差值表示植物的營養生長期,其中返青時間和植物營養生長期均使用侏略日表示。使用重要值綜合表示植物種群數量特征,計算公式如下:

其中相對高度為物種的平均高度與樣方所有物種的平均高度總和的比值;相對蓋度為物種的平均蓋度與樣方所有物種蓋度總和的比值;相對地上生物量為物種的平均地上生物量與樣方所有物種地上生物量總和的比值。
使用獨立樣本T檢驗比較兩種處理下相同深度的土壤溫度和土壤水分間的差異。使用一般線性模型(GLM)比較兩種處理間植物的返青時間、營養生長期以及重要值間的差異,將處理因子和區組因子作為固定效應檢驗主效應。數據表示方式為平均值±標準誤,差異顯著性水平P=0.05,數據的統計分析在IBM SPSS 26.0中進行,圖表的繪制在SigmaPlot 12.5中進行。
2 研究結果
2.1 枯落物累積對土壤溫度和土壤水分的影響
2015—2017年生長季內,枯落物累積顯著降低不同深度的土壤溫度(圖1;P<0.05)。在土壤深度2.5 cm處,2015—2017年生長季枯落物累積分別降低土壤溫度1.3—2.8℃、2.2—2.5℃和2.2—4.6℃。在土壤深度12.5 cm處,實驗期間枯落物累積分別降低土壤溫度1.5—3.2℃、2.1—2.4℃和2.2—2.7℃。

圖1 生長季土壤溫度的動態特征Fig.1 Dynamic characteristics of soil temperature in the growing seasonLA:枯落物累積處理,Litter accumulation treatment; CK: 對照處理,Control treatment; 不同小寫和大寫字母分別表示在0.05和0.01水平下兩種處理間存在顯著差異
2015—2017年生長季內,枯落物累積顯著提高不同深度的土壤水分(P<0.05),且對2.5 cm處的土壤水分影響更大(圖2)。在土壤深度2.5 cm處,2015—2017年枯落物累積分別提高土壤水分1.4%—4.3%、1.4%—6.3%和1.8%—10.4%。在土壤深度12.5 cm處,實驗期間枯落物累積分別提高土壤水分1.0%—3.7%、1.6%—5.0%和1.9%—3.4%。此外,與生長季中期(7月和8月)相比,枯落物累積在生長季早期(5月和6月)對土壤水分的提高更加明顯。例如,在2017年5月,枯落物累積分別提高土壤深度2.5 cm和12.5 cm處土壤水分10.4%和3.5%;在同年8月枯落物累積對土壤深度2.5 cm和12.5 cm處土壤水分的提高僅為2.2%和2.0%。
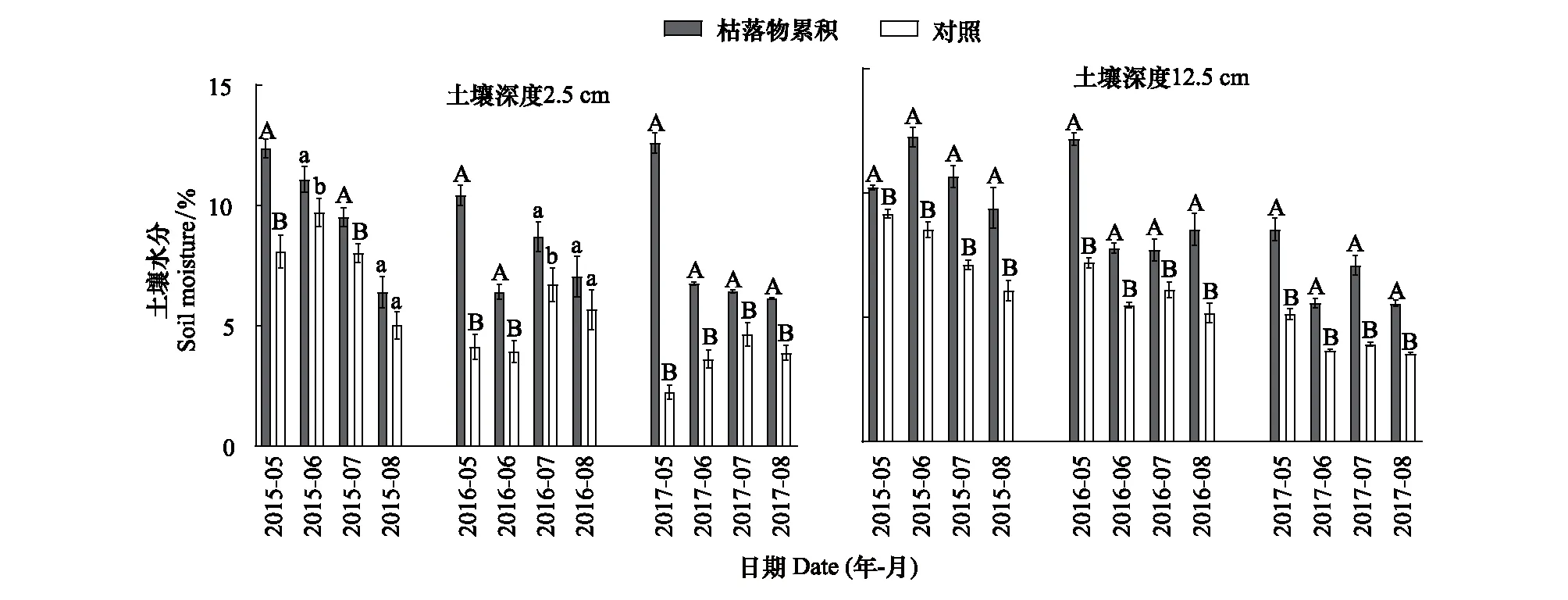
圖2 生長季土壤水分的動態特征Fig.2 Dynamic characteristics of soil moisture in the growing season
2.2 枯落物累積對大針茅和羊草種群生長發育的影響
大針茅和羊草種群的返青時間對枯落物累積具有一致的響應。2015—2017年枯落物累積顯著延遲大針茅和羊草種群的返青時間(圖3;P<0.05)。與羊草種群相比,大針茅種群在返青過程中受到更嚴重的消極影響。例如,實驗期間枯落物累積延遲大針茅返種群青時間高達16.9—24.2 d,而對羊草種群返青時間的延遲僅為5.2—6.7 d(圖3)。
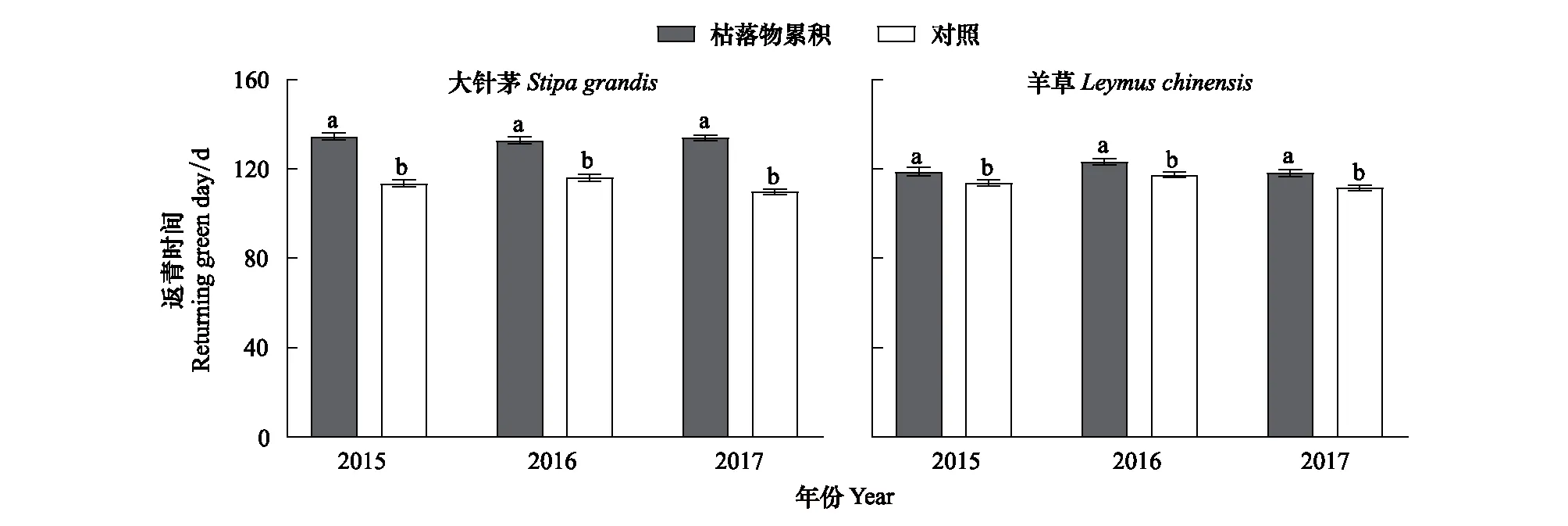
圖3 大針茅和羊草種群返青時間特征Fig.3 Characteristics of returning green time of S. grandis and L. chinensis
枯落物累積改變大針茅和羊草種群的生長發育過程。2015—2017年生長季早期,在枯落物累積影響下大針茅和羊草種群的植株高度低于對照處理中的;在生長季中期,枯落物累積影響下大針茅和羊草種群的植株高度高于對照處理中的(圖4)。例如,在2015年5月17日,枯落物累積影響下大針茅種群的植株高度僅為2.9 cm,對照處理中大針茅種群的植株高度高達15.7 cm。在2015年7月26日,枯落物累積下羊草種群的植株高度為51.5 cm,對照處理中羊草種群的植株高度僅為30.6 cm。與對照處理相比,枯落物累積影響下大針茅和羊草種群在返青后具有更快的生長速率。例如,在2016年5月9日—5月17日,枯落累積下大針茅種群的植株高度的生長速率高達1.72 cm/d,在對照處理中的僅為0.71 cm/d。
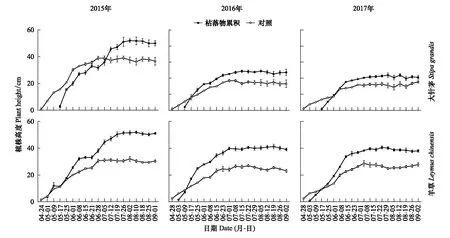
圖4 大針茅和羊草種群植株高度的動態特征Fig.4 Dynamic characteristics of plant heights of S. grandis and L. chinensis
與返青時間表現不同,枯落物累積對大針茅和羊草種群的營養生長期無顯著影響(圖5)。例如,實驗期間枯落物累積下大針茅和羊草種群的營養生長期為67—76 d和71—80 d,在對照處理中大針茅和羊草種群的營養生長期為64—78 d和65—78 d。
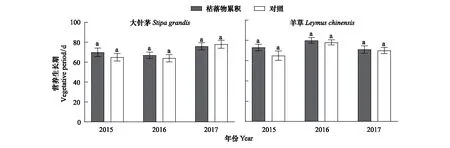
圖5 大針茅和羊草種群營養生長期特征Fig.5 Characteristics of vegetative period of S. grandis and L. chinensis
2.3 枯落物累積對大針茅和羊草種群數量特征的影響
大針茅和羊草種群數量特征對枯落物累積具有不一致的響應。枯落物累積顯著提高大針茅和羊草種群的高度(圖6;P<0.05)。枯落物累積顯著降低大針茅種群的蓋度和地上生物量,但顯著提高羊草種群的蓋度和地上生物量(除2016年羊草地上生物量外)(圖6;P<0.05)。例如,實驗期間枯落物累積降低大針茅種群的蓋度25.4%—55.2%,但提高羊草種群的蓋度達到160.9%—369.2%。由于大針茅和羊草種群的高度、蓋度和地上生物量的改變,進而導致大針茅和羊草種群的優勢度發生更替。枯落物累積顯著降低大針茅種群的重要值,但顯著提高羊草種群的重要值(圖6;P<0.05)。例如,實驗期間枯落物累積下大針茅和羊草種群在群落中的重要值分別為0.21—0.23和0.27—0.29,在對照處理中大針茅和羊草種群的重要值為0.34—0.40和0.13—0.19。

圖6 大針茅和羊草種群數量特征Fig.6 Characteristics of population characteristics of S. grandis and L. chinensist
3 討論
3.1 枯落物累積對群落微環境的影響
圍封草原枯落物累積引起群落熱量和水分的重新分配,綜合改變群落微環境特征。枯落物累積顯著降低土壤溫度并提高土壤水分,與前人的研究結果一致[25—26]。枯落物位于土壤與大氣間,具有強烈的物理阻隔作用,直接阻礙空氣與土壤間的熱量交換。有研究表明在白天枯落物累積強烈降低土壤熱量的吸收,顯著降低土壤溫度;在夜晚枯落物累積可以有效減少土壤熱量的散失,提高土壤溫度[27]。由于內蒙古草原在生長季具有強烈的太陽輻射和較大的晝夜溫差,因此枯落物累積綜合導致生長季土壤溫度的降低。枯落物的遮陰效應可以有效減少土壤水分的蒸發,直接提高土壤水分[16]。此外,土壤溫度和土壤水分具有強烈的耦合效應,土壤溫度的降低也將減少土壤水分的蒸發,間接提高土壤水分。在內蒙古高原的春季時期,具有氣溫迅速回升且降水稀少的特征,同時此階段也是植物返青后生長過程中對水分需求最重要的階段。生長季早期枯落物累積后土壤更加濕潤,表明枯落物累積對植物返青后的生長具有積極效應,也為在枯落物累積影響下大針茅和羊草種群返青后具有較快的生長速率提供了解釋。在長時間尺度,由于枯落物累積對土壤溫度和土壤水分的影響具有累積效應,圍封草原群落微環境將趨于中生化[14]。群落微環境的中生化也進一步通過影響植物功能性狀、種群生長發育及其適應性等方面,最終驅動圍封草原植物群落結構與功能變化[13,18]。
3.2 枯落物累積對植物種群生長發育的影響
植物在適應環境變化的過程中通過改變物候過程,綜合調節其生長發育過程。圍封草原枯落物累積顯著延遲植物種群的返青時間。有研究表明溫度是影響內蒙古溫帶草原植物物候重要的生態因子且植物在返青過程中需要達到一定的積溫[28]。因此,內蒙古圍封草原枯落物累積引起的土壤溫度降低是導致植物種群返青時間延遲的主要原因。然而,Zhu等人在青藏高原圍封高寒草原研究中發現,枯落物累積顯著提前高山嵩草(Kobresiapygmaea),釘柱委陵菜(Potentillasaundersiana)和楔葉委陵菜(Potentillacuneata)的返青時間[29]。青藏高原草原地區的海拔遠高于內蒙古草原地區,因此在氣候方面形成了與內蒙古溫帶草原氣候截然不同的特征。常年低溫、強烈的太陽輻射和干旱是青藏高原草原地區主要的氣候特征[30]。植物在長期適應低溫后,對溫度變化的敏感性降低。因此,Zhu等人認為青藏高原高寒草原圍封后表層土壤水分的提高是導致這些植物返青時間提前的主要因子[29]。這些研究結果的差異也表明不同生態區域內的圍封草原植物的物候過程在受區域氣候的影響下,同時也易受到群落微環境內不同生態因子(溫度和水分)的調控。
枯落物累積通過改變群落微環境特征影響植物種群的生長發育過程。圍封草原枯落物累積后植物在返青后具有更快的生長速率。土壤水分是限制溫帶干旱—半干旱地區草原生態系統植物生長發育的關鍵因子。在植物返青后圍封草原枯落物累積通過提高土壤水分,加速植物生長[25,31]。此階段植物的高度較低,枯落物累積也通過物理阻隔作用降低群落光照強度,提高植物對光照的競爭,進而提高圍封草原植物的生長速率[14,17]。在生長季中期,隨著植株高度的增加,在圍封草原中光照對植物生長的影響逐漸減弱,土壤水分成為主導植物生長發育的關鍵因子。植物種群的返青時間和生長發育過程對圍封草原枯落物累積具有敏感的響應,因此該指標可作為圍封草原管理中植物生長狀態的監測指標。此外,枯落物累積對植物的營養生長期無顯著影響(圖5),表明與返青時間和植株高度動態等指標相比,其對圍封草原群落微環境變化具有不敏感的響應。這也可能由于植物的營養生長期主要受自身生物學特性的調控,溫度和水分等環境因子的變化對其影響微弱。圍封草原植物種群生長發育過程的變化,進一步改變不同植物種群對光照和水分等資源的競爭能力,最終改變植物種群數量特征。
圍封草原枯落物累積的影響下,大針茅種群具有更晚的返青時間,表明與羊草種群相比,枯落物累積對大針茅種群的具有更明顯的消極影響。物種生物學特性及生態適應性差異是導致大針茅和羊草種群對枯落物累積響應差異的主要原因[32—33]。在根系分布方面,與大針茅根系分布(0—30 cm)相比,羊草種群的根系分布(0—10 cm)相對較淺[34]。因此,在生長季早期羊草種群在返青過程中受到較小的影響。我們在野外也觀察到羊草為根莖禾草且其枯落物呈立枯狀態,對羊草種群的返青過程產生較小的影響;大針茅為叢生禾草其枯落物常常堆積于植株上,嚴重抑制大針茅種群的返青過程。在水分適應方面,與大針茅種群相比,羊草種群更加適應相對濕潤環境[23]。因此,圍封草原枯落物累積引起土壤水分的提高更加有利于羊草種群的生長發育過程,導致大針茅種群的蓋度、地上生物量和重要值的顯著降低和羊草種群蓋度、地上生物量和重要值的顯著增加。大針茅和羊草種群重要值排序的更替表明圍封草原植物群落發生演替。因此,本研究也從植物種群的角度為典型草原圍封后群落由叢生禾草草原向根莖禾草草原演替的觀點提供進一步的支持[14]。
4 結論
圍封草原枯落物累積通過改變群落熱量和水分等環境因子的重新分配,對不同植物種群的生長發育過程產生非對稱影響。枯落物累積降低土壤溫度并提高土壤水分,導致群落微環境趨于陰涼和濕潤。與大針茅種群相比,羊草種群在物候和生長過程等方面對枯落物累積后相對濕潤的微環境更加適應,進而提高其對資源的競爭能力。大針茅和羊草種群生長發育過程的改變綜合引起兩種植物種群數量特征的變化,導致其重要值發生更替,最終驅動圍封草原植物群落演替。本研究從植物種群的角度為闡明圍封草原群落結構與功能改變提供新視角。
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
小讀者(2021年2期)2021-03-29 05:03:48
華人時刊(2019年13期)2019-11-17 14:59:54
文苑(2018年22期)2018-11-19 02:54:14
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
中國塑料(2016年3期)2016-06-15 20:30:00
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
