模擬干旱和磷添加對熱帶低地雨林葉凋落物分解的影響
2022-08-31 06:08:26王文秀欒軍偉李絲雨梁昌強孔祥河劉世榮
生態學報 2022年15期
王文秀,欒軍偉,王 一,楊 懷,趙 陽,李絲雨,梁昌強,孔祥河,劉世榮
1 國際竹藤中心, 竹藤科學與技術重點實驗室, 北京 100102 2 中國林業科學研究院森林生態環境與自然保護研究所, 國家林業和草原局森林生態環境重點實驗室, 北京 100091 3 海南甘什嶺自然保護區, 三亞 572014
森林凋落物是森林植物在其生長發育過程中新陳代謝的產物,在森林生態系統養分循環中起著重要作用[1]。通過凋落物的分解作用,有機物質從地上植被轉移進入森林土壤,逐步把養分歸還給土壤[1—2],向大氣釋放CO2,為植物和微生物提供養分[3—4]。凋落物分解過程受自身基質質量[5—6]、環境氣候條件[7]、土壤理化性質[8]、分解者群落調控[9]。并且,在已有關于凋落物分解研究中,不同功能型(常綠落葉樹種、固氮非固氮樹種、葉質地分解難易程度)植物的葉片基質質量存在差別[10—11],落葉樹種、固氮樹種葉凋落物相比于常綠樹種、非固氮樹種葉凋落物分解較快[11—12]。凋落物分解是生態系統中碳(C)、氮(N)、磷(P)等營養元素循環的重要環節,是生物圈與大氣圈之間C、N流通的重要環節[13—15]。
溫室氣體排放的增加所導致的全球范圍內溫度升高、水文循環改變成為科學界的共識[16],影響地區包括熱帶地區[17—18],并且有研究學者指出未來氣候變化可能導致極端干旱事件發生的頻度、強度增加[16, 19]。熱帶雨林在水分脅迫條件下,會變得非常脆弱,地上生物量減少[20—21]、樹木死亡率升高[22—23]。有研究學者在亞馬遜地區開展減雨試驗,連續4a時間減少了60%的雨水,得出樹木死亡率增加[24]、冠層含水量減少[25]、地上凈初級生產量減少[26]的結論。
氣候變化從許多方面對凋落物分解產生影響,其中水分條件的變化是影響凋落物分解的重要方面之一。因為降水量減少,會削弱雨水對凋落物的破碎、淋溶作用[27];其次,土壤生物的豐富度與活性受土壤含水量調控,間接使凋落物分解與營養元素釋放受影響[28];此外,降水量的減少在一定范圍內會使植被生產效率降低,進而導致凋落物生產量降低[29]。干旱脅迫降低凋落物的分解速率這一觀點被許多研究支持,比如Willcock等人[30]研究表明,降雨量減少會顯著抑制土壤生物活性,從而抑制凋落物分解;有大量研究表明凋落物分解速率和降雨量正相關,水分不足會降低凋落物分解速率[30—31];干旱處理減慢了凋落物分解速率,并且改變了混合凋落物間的相互作用[32]。所以,氣候變化可能通過降雨量減少、干旱脅迫影響凋落物分解[33]。甚至,氣候變化導致的干旱事件會改變森林凋落物的分解過程,增加對森林生態系統C收支預測的不確定性[34]。
熱帶、亞熱帶地區,土壤顯酸性,土壤中的P元素易與鐵鋁氧化物發生結合反應,并且P主要來源于礦石的淋溶,所以土壤中P元素有限;同時土壤中的P元素極易隨著雨水的淋溶、沖刷作用流失,因此普遍認為熱帶地區受P限制[35]。并且有研究證實,P的可獲得性確實限制了熱帶地區的凈初級生產力[35]、凋落物分解[36—39]。有研究學者指出,P添加可提高植物地下生物量的累積[40];也有研究學者指出,在土壤中添加P,對于凋落物的分解速率沒有顯著影響,但是抑制了N損失[41];在土壤N含量高、P含量低的熱帶森林生態系統中,P添加可促進葉凋落物分解[37—38],但也有研究得出相反結論,在Costa Rich、Brazil的熱帶生態系統實驗中,認為P添加對葉凋落物分解過程中的質量損失沒有較大作用[42—43]。目前,土壤中P元素可用性是否調控干旱脅迫對熱帶森林凋落物分解的影響并不清楚。
基于以上研究背景,本文提出如下研究假設:模擬干旱處理使試驗樣地的土壤含水量降低,減慢了葉凋落物分解過程中生物、化學反應速率,抑制了參與凋落物分解的分解者的活性,抑制了凋落物分解;P添加處理增加了土壤中P元素的可獲得性,可能改變土壤微生物群落結構,尤其是真菌群落,進而可能影響凋落物分解速率;基于上述原因,本文推測土壤中水分含量降低與P可利用性提高可能存在交互作用,土壤中P可利用性增加可在一定程度上緩解土壤水分含量下降對葉凋落物分解的抑制作用。
1 材料與方法
1.1 研究區概況
本實驗研究地位于海南省三亞市甘什嶺省級自然保護區,國家林草局海南三亞竹藤伴生林生態系統定位觀測研究站。該區屬于熱帶低地雨林,地理坐標是109°34′E—109°42′E,18°20′N—18°21′N。地貌是低山丘陵,年平均日照時數大約為2563.0h,年均溫25.4℃,每年有明顯干濕兩季,旱季時間為當年10月至明年5月,除此之外均為雨季時間,平均年降水量約為1800mm,屬熱帶海洋性季風氣候。土壤母質為花崗巖,屬粗砂粘紅壤,砂礫含量大,部分區域土層貧瘠,雨水沖刷導致巖層裸露率高,部分區域達到70%以上,平均林冠高度為9.7—12.1m,部分區域至15m以上,差異較大,郁閉度0.6—0.9,區域內群落結構復雜,植被種類豐富[44]。喬木層主要有鐵凌(Hopeareticulate)、青皮(Vaticamangachapoi)、阿芳(Alphonseamonogyna)、瓊楠柿(Diospyroshowii)、崖柿(Diospyroschunii)等;草本植物主要有高稈珍珠茅(Scleriaelata)和益智(Alpiniaoxyphylla);藤本植物主要有多葉花椒(Zanthoxylummultijugum)、清香藤(Jasminumlanceolarium)、錫葉藤(Tetraceraasiatica)等[45]。
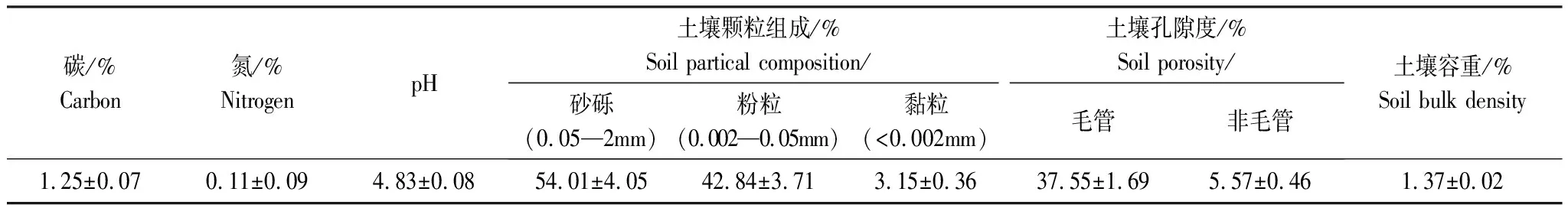
表1 實驗樣地土壤理化性質
數據為平均值±標準誤
1.2 實驗設計
依托海南三亞竹藤伴生林生態系統定位觀測研究站的長期穿透雨減少與P添加雙因素交互控制實驗平臺,共3個區組,每個區組由4個40m×40m的樣地組成,并采取4種不同的處理,分別為穿透雨減少(模擬干旱)(D,50%穿透雨減少)處理、P添加(P,50Kg P hm-2a-1)處理、模擬干旱×P添加(DP,50%穿透雨減少×50Kg P hm-2a-1)處理、對照(CK)樣地。每塊樣地四周鋪設PVC軟板,阻止樣地內外的水分交換。穿透雨減少采用不銹鋼管結合PEP透光膜在林冠下離地表約2m處建立攔截架,攔截50%的林內穿透雨、導入排水槽排出樣地;P添加采取人工撒施Ca(H2PO4)2·H2O(TSP)于地表,施P強度為50Kg P hm-2a-1,從2019年7月開始,起初施P頻率為1次/2月,2020年后穩定在1次/4月(施P頻率降低,總量不變)。本實驗根據樣地內各樹種多度、碳固持類型、葉質地,選取4個相對密度較高樹種的葉凋落物為研究對象,分別為鐵凌(Hopeaexalata)、白茶樹(Koilodepasbainanense)、黑葉谷木(Memecylonnigrescens)、山油柑(Acronychiapedunculata),2019年10月收集鐵凌、白茶樹、黑葉谷木、山油柑葉凋落物,將收集的葉凋落物于70℃下烘干24h,用于測定4種葉凋落物初始化學成分含量與野外分解試驗布設。本研究采用尼龍網分解袋,大小為20cm×30cm,網孔1mm×1mm,每個分解袋中裝8g葉凋落物(4個樹種×4個處理×6個重復×4次回收=384袋)。于2019年11月,將全部葉凋落物分解袋隨機布置于樣地地表,分解袋回收時間分別為2020年1月、5月、6月、8月(取樣時間受疫情影響),對應分解時間分別為57d、178d、230d、273d。每次回收,每個樹種葉凋落物4個處理下分別取6袋(每個樣地分別取2袋),取樣后將分解袋中的雜物泥沙處理干凈,于70℃烘干24h,稱量、粉碎、測定C、N含量。

表2 4種凋落物樹種性質
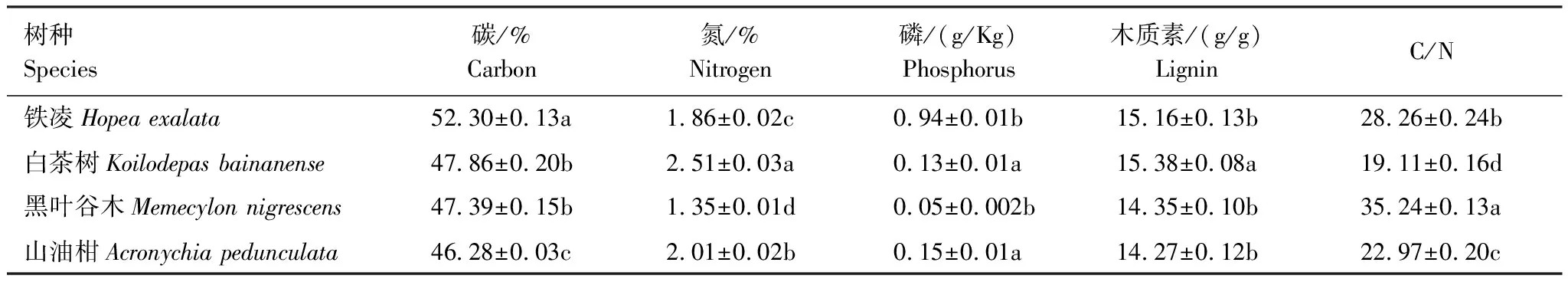
表3 4種葉凋落物化學成分含量
1.3 凋落物化學成分測定
凋落物總C含量的測定:元素分析儀(ECS 4010 CHNSO, Costech, Italy)。
凋落物總N含量的測定:元素分析儀(ECS 4010 CHNSO, Costech, Italy)。
凋落物初始全P含量的測定:硫酸-高氯酸法消煮(300℃,3h),采用鉬銻鈧比色法通過化學元素分析儀(Smartchem 300, AMS-Alliance, Italy)。
凋落物初始木質素含量的測定:重量法(GB/T 35818/2018)。
1.4 葉凋落物質量殘存率、分解系數以及養分損失率計算


C、N損失率CL、NL(%):CL=[(M0×C0)-MT×CT]/(M0×C0)×100%;
NL=[(M0×N0)-MT×NT]/(M0×N0)×100%
式中,Mt表示分解t時間后凋落物殘余質量占初始質量的比例(%);MT表示分解t時間后凋落物殘余質量(g);M0表示凋落物初始質量(g);t表示分解時間(a);k表示分解系數(a-1);C0、CT表示凋落物初始C含量、回收時剩余C含量(%);N0、NT表示凋落物初始N含量、回收時剩余N含量(%)。
1.5 數據分析
使用SPSS 22.0軟件進行統計分析,用單因素方差分析(One-way ANOVA),比較4種葉凋落物初始化學性質之間的差異是否顯著;用一般線性模型(General Linear model),分析凋落物基質質量、分解時間、模擬干旱、磷添加對凋落物分解系數、C、N損失的影響;用線性回歸分析(Linear regression analysis),分析葉凋落物分解系數、C、N損失與初始化學組分含量的關系;用雙因素方差分析(Two-way ANOVA),檢驗模擬干旱、P添加處理分別對4個樹種葉凋落物分解系數、C、N損失的影響。作圖軟件使用Origin 2018。
2 結果與分析
2.1 基質質量對葉凋落物分解的影響
凋落物基質質量對葉凋落物分解系數、C、N損失率均有顯著影響(P<0.001;P<0.001;P<0.001)(表4),表現為白茶樹葉凋落物的分解系數大于其它3個樹種葉凋落物的分解系數,白茶樹葉凋落物的C損失率大于
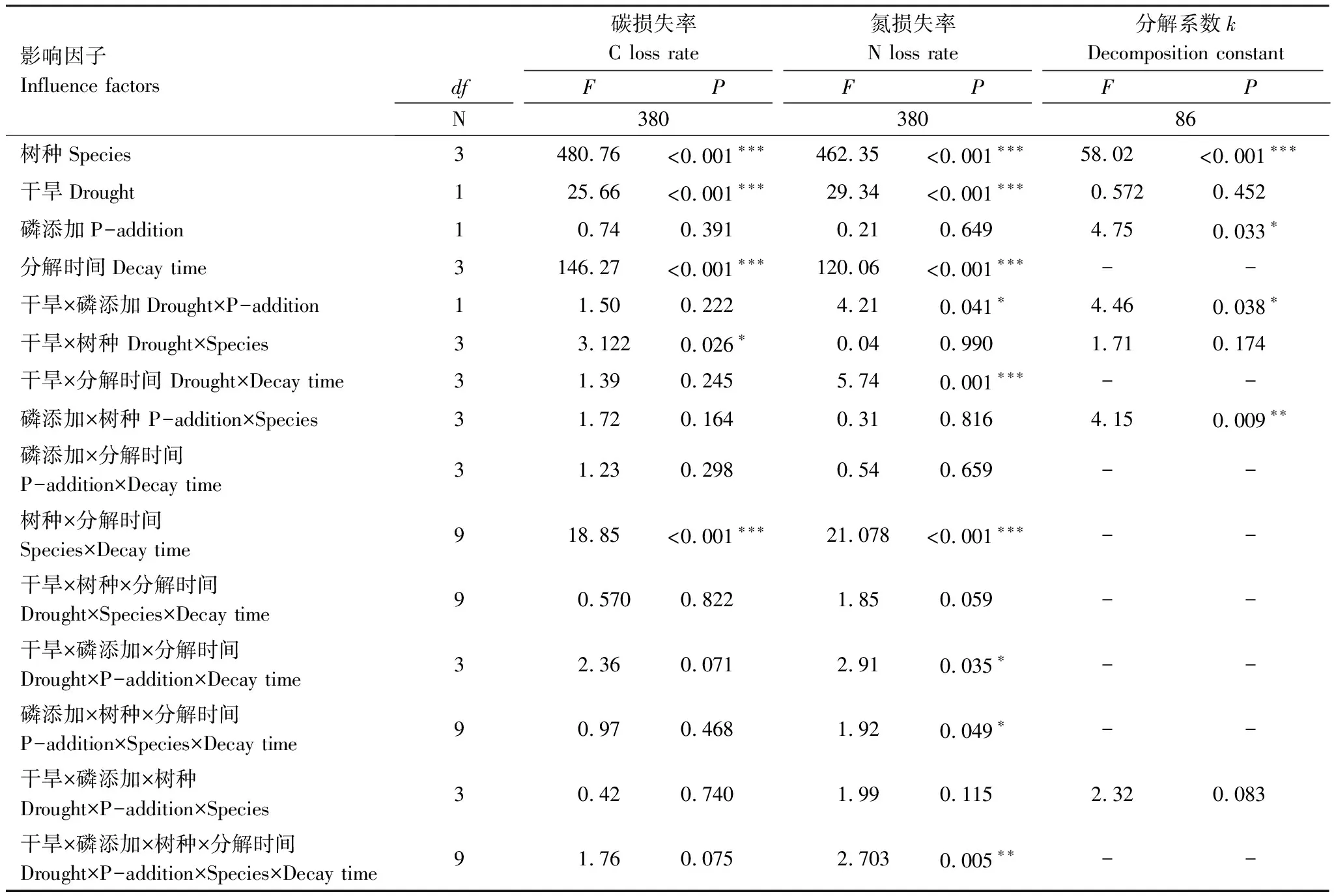
表4 樹種、模擬干旱、磷添加、分解時間及交互作用對葉凋落物分解的影響
其它3個樹種葉凋落物的C損失率,N損失率由大到小依次為白茶樹、山油柑、鐵凌、黑葉谷木(圖1)。并且,葉凋落物分解系數、C、N損失率與凋落物初始N含量、C/N比存在顯著正線性相關、負線性相關關系,比如,初始N含量較高、C/N比較低的白茶樹葉凋落物分解較快,C、N損失率較高(圖2)。

圖1 4個樹種葉凋落物碳、氮損失率、分解系數Fig.1 Decomposition constants, C, N loss rates of four-tree species leaf litter
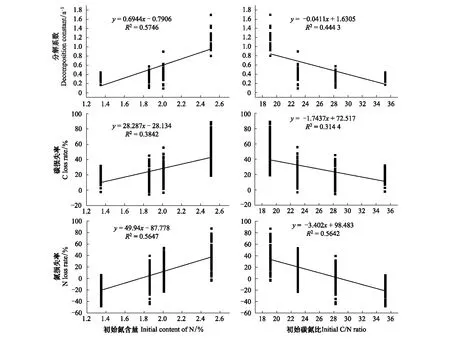
圖2 葉凋落物初始氮含量、碳氮比與碳、氮損失率、分解系數的關系Fig.2 The relationship between initial N content, C/N ratio and C, N loss rate, decomposition constant
2.2 模擬干旱處理對葉凋落物分解的影響
4個樹種葉凋落物分解過程中的分解系數,模擬干旱處理使鐵凌葉凋落物分解系數顯著降低了11%(P=0.012),但對其它樹種葉凋落物無顯著影響(圖3)。
4個樹種葉凋落物的C含量,分解57d后,模擬干旱處理使白茶樹葉凋落物的C損失率下降了14.46%(P=0.042)(圖4);分解178d后,模擬干旱處理使白茶樹葉凋落物C損失率顯著下降了12.24%(P=0.044)(圖5);分解230d后,模擬干旱處理使白茶樹葉凋落物C損失率顯著上升了0.6%(P=0.033)(圖6);分解273d后,模擬干旱處理對鐵凌、白茶樹葉凋落物C損失率有顯著影響,分別使鐵凌葉凋落物C損失率顯著下降了21.65%(P=0.050),使白茶樹葉凋落物C損失率顯著下降了13.72%(P=0.010)(圖7)。并且模擬干旱處理對葉凋落物C損失率的影響同時還與葉凋落物的樹種有關(P=0.026)(表4)。
模擬干旱處理對葉凋落物N損失率均產生顯著影響(P<0.001)(表4)。4個樹種葉凋落物的N含量,分解57d后,模擬干旱處理對4個樹種葉凋落物N損失均產生顯著影響,分別使鐵凌葉凋落物N損失率下降了806.62%(P=0.003);使白茶樹葉凋落物N損失率下降了64.93%(P=0.022);使黑葉谷木葉凋落物N損失率上升了15.13%(P=0.046);使山油柑葉凋落物N損失率下降了195.34%(P=0.015)(圖4);分解178d后,模擬干旱處理使山油柑葉凋落物N損失率顯著下降了108.69%(P=0.024)(圖5);分解230d后,模擬干旱處理使黑葉谷木葉凋落物N損失率顯著下降245.74%(P=0.001)(圖6);分解273d后,模擬干旱處理使白茶樹葉凋落物N損失率顯著下降了11.95%(P=0.005)(圖7)。
2.3 磷添加處理對葉凋落物分解的影響
P添加處理對葉凋落物分解系數存在顯著影響,使葉凋落物分解系數升高0.64%(表4;P=0.033),但該影響與葉凋落物樹種無關(表4;圖3)。
4個樹種的葉凋落物C含量,分解57d后,P添加處理使鐵凌葉凋落物C損失率增加183.42%(P=0.004)(圖4);分解178d后,P添加處理對4個樹種葉凋落物C損失率均不產生顯著影響(圖5);分解230d后,P添加處理對4個樹種葉凋落物C損失率均不產生顯著影響(圖6);分解273d后,P添加處理對4個樹種葉凋落物C損失率均無顯著影響(圖7)。
4個樹種的葉凋落物N含量,分解57d后,P添加處理對4個樹種葉凋落物N損失率均無顯著影響(圖4);分解178d后,P添加處理使黑葉谷木葉凋落物N損失率顯著下降了58.62%(P=0.039)(圖5);分解230d后,P添加處理對4個樹種葉凋落物N損失率均不產生顯著影響(圖6);分解273d后,P添加處理對4個樹種葉凋落物N損失率均無顯著影響(圖7)。
2.4 模擬干旱與磷添加處理對葉凋落物分解的交互效應
模擬干旱與P添加處理葉凋落物分解系數產生顯著交互效應,在僅模擬干旱處理下未對葉凋落物分解系數產生顯著影響,但當模擬干旱、P添加處理同時存在時,使葉凋落物分解系數降低了18.74%(表4,P=0.038),且交互效應與葉凋落物樹種無關(表4;圖3)。
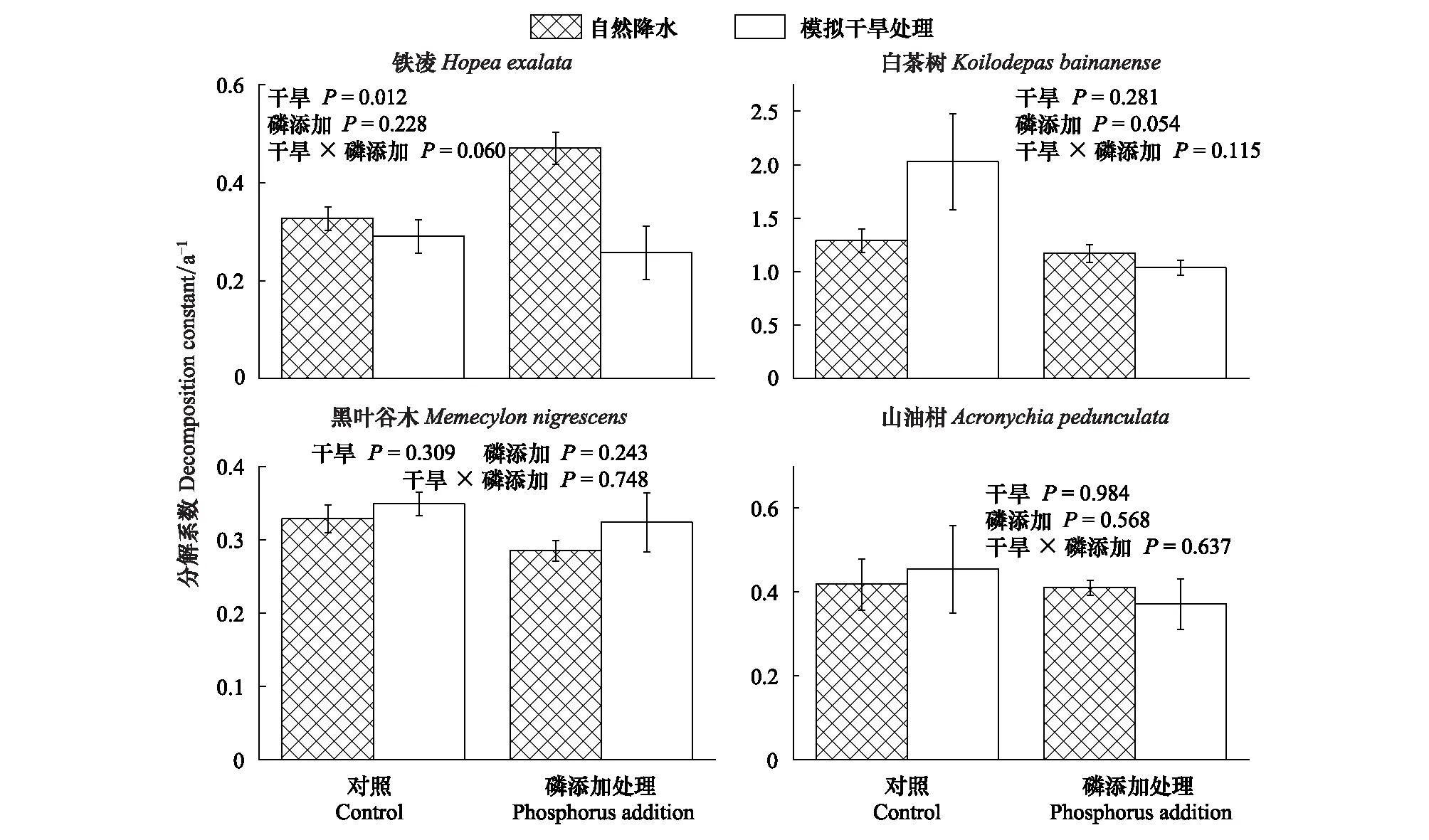
圖3 4種葉凋落物的分解系數Fig.3 Decomposition constants of four-tree species leaf litter
4個樹種的葉凋落物C含量,分解57d后,模擬干旱與P添加處理對4個樹種均未產生顯著交互效應(圖4);分解178d后,模擬干旱與P添加處理對4個樹種葉凋落物C損失率均不產生顯著交互效應(圖5);分解230d后,模擬干旱與P添加處理對白茶樹葉凋落物C損失率產生顯著交互效應,在僅模擬干旱處理下,白茶樹葉凋落物C損失率增加0.06%(P=0.033),但當模擬干旱與P添加同時作用時,C損失率顯著下降了31.95%(P=0.028)(圖6);分解273d后,模擬干旱與P添加處理對4個樹種葉凋落物C損失率均無顯著交互效應(圖7)。

圖4 分解57d后,4種葉凋落物的碳、氮損失率Fig.4 C, and N loss of four-tree species leaf litter, after decaying 57 days

圖5 分解178d后,4種葉凋落物的碳、氮損失率Fig.5 C, and N loss of four-tree species leaf litter, after decaying 178 days
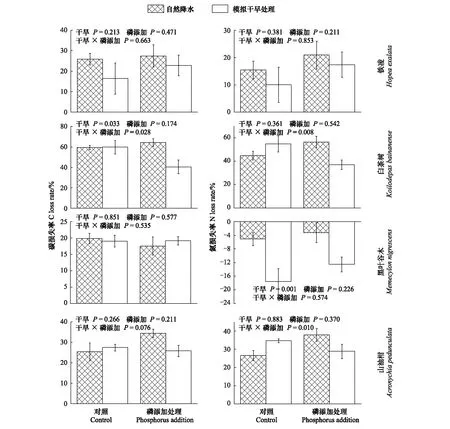
圖6 分解230d后,4種葉凋落物的碳、氮損失率Fig.6 C, and N loss of four-tree species leaf litter, after decaying 230 days
4個樹種的葉凋落物N含量,分解57d后,模擬干旱與P添加處理對黑葉谷木葉凋落物N損失率有顯著交互效應,在僅模擬干旱處理下,黑葉谷木葉凋落物N損失率增加15.13%(P=0.046),但當模擬干旱與P添加同時作用時,N損失率下降了138.6%(P=0.023)(圖4);分解178d后,模擬干旱與P添加處理對4個樹種葉凋落物均不產生顯著交互效應(圖5);分解230d后,模擬干旱與P添加處理對白茶樹、山油柑葉凋落物N損失率存在顯著交互效應,在僅模擬干旱處理下,未對白茶樹、山油柑葉凋落物產生顯著影響,但當模擬干旱與P添加處理同時作用時,白茶樹葉凋落物N損失率顯著下降了17.44%(P=0.008),山油柑葉凋落物N損失率顯著上升了8.35%(P=0.010)(圖6);分解273d后,模擬干旱與P添加處理對4個樹種葉凋落物均無顯著交互效應(圖7)。

圖7 分解273d后,4種葉凋落物的碳、氮損失率Fig.7 C, and N loss of four-tree species leaf litter, after decaying 273 days
3 討論
凋落物分解系數、C、N損失與凋落物基質質量緊密相關[41]。在本試驗中,分解結束時,白茶樹葉凋落物較其它三種葉凋落物分解剩余質量最少、C、N損失率最高,這可能因為白茶樹葉凋落物初始N含量最高、CN比最低。這與一些研究學者的結論相似,凋落物分解速率與初始N含量呈極顯著正相關,是控制分解速率最主要的因素[47];N含量高、CN比低的凋落物滿足土壤微生物養分需要,更易被微生物分解[48]。
本研究中,模擬干旱處理對葉凋落物分解的影響表現為抑制作用,使鐵凌葉凋落物分解系數下降11%,使鐵凌、白茶樹葉凋落物C損失率下降,使鐵凌、白茶樹、黑葉谷木、山油柑葉凋落物N損失率下降。與前人研究結果一致,Salamanca等人在溫帶森林開展有關降雨量變化對凋落物分解影響的研究中發現,降雨量對凋落物分解的控制主要是通過雨水的淋溶作用與微生物、動物群落活動,并且根據土壤呼吸、微生物生物量數據可知,降雨量減少降低微生物、動物群落豐富度與活性,間接抑制凋落物分解[49]。同時,在本試驗中,模擬干旱處理對4個樹種葉凋落物的抑制作用存在差異,可能因為不同樹種葉凋落物的基質質量不同。這與前人的研究結果一致,Sanaullah等人表明不同的凋落物分解中基質質量會調控干旱脅迫對分解的影響[50],原因可能是在分解過程中,微生物分解者更傾向于分解質量較高(初始N含量較高)的凋落物,所以當包括干旱在內的非生物條件限制微生物分解者的活性時,該種凋落物分解受抑制程度最大[50]。
本研究中,P添加處理對葉凋落物分解產生影響,但對C、N損失產生影響不同,促進C損失但抑制N損失,比如使鐵凌葉凋落物C損失率升高,使黑葉谷木N損失率下降。原因可能是土壤中P含量的升高,促進微生物分解高C物質的能力;N和P均是植物生長過程中重要的限制因素,相互獨立又相互影響[51—52],所以土壤中P含量的升高可能會影響凋落物分解中的N元素變化。首先,在本研究中,P添加處理對鐵凌葉凋落物的C損失有促進作用,這與一些研究結果相似。有研究表明提高土壤中的P含量對凋落物分解有促進作用,土壤中較高的P含量可提高微生物群落與胞外酶活性[53],進而提高微生物分解高C物質的能力,比如木質素、纖維素[53—56],所以促進凋落物中C釋放。其次,在本研究中,P添加處理對黑葉谷木葉凋落物N損失有抑制作用,曾有研究學者表明,比如P添加對凋落物分解產生抑制作用或無明顯作用[43, 54, 57—59],原因可能是P添加抑制了微生物的P礦化作用[60],當P添加時微生物就有了足夠的P,不必要通過分解凋落物等有機物來獲取P[58],所以抑制了微生物分解者對凋落物的分解,但是抑制微生物P礦化這一說法很少被承認[61];還有研究學者認為P添加對凋落物分解的抑制作用因為土壤中P含量的增加提高了微生物利用C的能力,但是微生物利用C完成自身生長代謝需要,而不是用于合成凋落物分解所需的酶[62—63]。同時也有研究學者認為P添加對凋落物分解N損失存在促進作用,土壤中P的可獲得性影響著土壤微生物的豐富度與活性,比如氨氧化古菌,進而增加了硝化反應速率,促進了微生物分解者從凋落物中獲取N[64]。不同研究關于P添加對凋落物分解的影響結論不同,原因可能是P添加處理時的藥品劑量不夠大、對樣地的處理時間不夠長或者P限制不是研究地區凋落物分解的主要影響因子[38],也有研究學者認為高濃度的P添加處理可能對P限制地區的凋落物分解有抑制作用[58]。試驗過程中,施P頻率發生變化,但施P總量沒有變化,并且P元素在土壤中屬于緩釋,所以本試驗不將施P頻率改變作為影響分解結果的主要因子。
本試驗中,模擬干旱×P添加處理對葉凋落物分解產生了顯著的交互效應。P添加緩解甚至反轉了干旱處理對葉凋落物分解過程中C、N損失率的作用,說明了降水量與土壤P含量均是該地區的影響因子,以及提高土壤中P的可用性,可改變干旱脅迫的影響。前人的研究中也得到相似觀點,比如有研究學者關于水分條件、P限制哪個是溫帶農作物生產量的主要限制因子實驗中得出,在水分脅迫條件下,P添加使植物對干旱脅迫的耐受能力更強,產生更多的生物量[65]。有研究表明P添加與水分脅迫的交互作用與土地類型有關,在中度干旱的條件下,P添加有助于在黑土中生長的植物產生更多的生物量,但在砂土中不存在促進作用[66]。也有研究得到相反結論,比如有研究學者表明,干旱脅迫與P添加同時存在情況下,并未對土壤呼吸產生顯著交互效應[67]。原因可能在于研究地區環境因素不同,土壤中P的可用性對不同生態系統類型的限制程度不同,或者P添加處理的濃度與時間不同,所以處理產生的效果不同。有研究指出,干旱處理與營養元素添加均是影響土壤與微生物呼吸等活動、物種豐富度的影響因子,但是他們之間的交互作用機制仍不清晰;干旱對土壤及微生物的影響作用強于P添加的作用,干旱抑制了土壤的呼吸作用,使土壤微生物群落中的多樣性降低;但是,干旱與磷添加同時作用時,土壤呼吸未被抑制,土壤微生物群落中豐富度仍發生變化[67]。在本試驗中,干旱與P添加對凋落物分解的影響機制,本研究猜測可能是,干旱抑制了土壤中參加分解的微生物的活性與豐富度,但是土壤中P可利用性的提高,促進了一些微生物的活性與豐富度,所以二者存在交互作用。但是,干旱與P添加哪個影響因子的作用更強,在本試驗中,無法得出。
4 結論
本研究驗證了不同樹種葉凋落物因質量不同,分解存在差異;干旱脅迫抑制葉凋落物分解,但對不同樹種葉凋落物的影響存在差異;土壤中P含量的升高會促進葉凋落物的C損失、抑制葉凋落物的N損失;提高土壤中P的可獲得性在一定程度上可緩解干旱脅迫對凋落物分解的抑制。綜合本試驗與以往研究,模擬干旱、P添加處理對葉凋落物分解的影響結果,可能與研究地土壤理化性質、凋落物基質質量、施P濃度、時間相關。在本試驗中,也有一些不足希望在以后的試驗中有所改進,比如在實驗設計中,應對減雨處理與P添加處理的土壤水分含量與P含量進行監控,檢驗處理是否在試驗中產生效果;并且減雨處理與P添加處理可以設定一些梯度,更加細致的討論土壤水分與不同程度的P可利用性對葉凋落物分解等生態過程的影響程度;微生物分解是凋落物分解過程中的一個重要途徑,所以關于凋落物分解的研究離不開與微生物相關的實驗,在未來除關注凋落物分解過程中的養分元素變化外,還需要關注微生物的變化。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
當代陜西(2021年2期)2021-03-29 07:41:24
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國塑料(2016年3期)2016-06-15 20:30:00
