雅魯藏布江中上游白草主要分布區(qū)植物群落特征
2022-08-31 06:02:08張力天楊時(shí)海王曉麗秦金萍董瑞珍邊巴普尺米瑪頓珠索朗次仁張樹淼仁增曲扎馬玉壽
生態(tài)學(xué)報(bào) 2022年15期
張力天,楊時(shí)海,劉 煒,王曉麗,秦金萍,董瑞珍,邊巴普尺,米瑪頓珠,索朗次仁,張樹淼,張 敏,仁增曲扎,馬玉壽,*
1 青海大學(xué)畜牧獸醫(yī)科學(xué)院, 西寧 810016 2 日喀則市百亞成農(nóng)牧產(chǎn)品加工有限公司, 日喀則 857800 3 西藏云旺實(shí)業(yè)有限公司, 日喀則 857000 4 蘭州大學(xué)生命科學(xué)學(xué)院, 蘭州 730000
種群結(jié)構(gòu)反映了種群內(nèi)不同年齡個(gè)體數(shù)量的組成情況、種群數(shù)量動(dòng)態(tài)及其發(fā)展趨勢,并在很大程度上體現(xiàn)種群與環(huán)境之間的相互關(guān)系,以及種群在群落中的作用和地位[1]。植物群落是植物種群與環(huán)境因子共同構(gòu)成的一個(gè)有機(jī)整體,不同物種種群在群落中所處空間分布格局取決于物種的生物學(xué)特性、生境條件及其交互作用[2—3]。探究植物群落多樣性及其維持機(jī)制,是了解植物群落生態(tài)功能和穩(wěn)定的基礎(chǔ),也一直是群落生態(tài)學(xué)研究的中心議題[3]。海拔梯度下,水、熱因子可在相對較小范圍內(nèi)急劇變化[4—5],成為物種豐富度梯度格局研究的重要方面[6],特別是在陸地生態(tài)系統(tǒng)中江河流域的河岸區(qū)是生物種群和各種環(huán)境因子相互影響的潛在敏感地區(qū)[1],掌握河岸生物多樣性分布格局與各種環(huán)境因子的關(guān)系,對于河岸生態(tài)系統(tǒng)保護(hù)特別是流域生態(tài)安全具有十分重要的科學(xué)意義[7—10]。
白草(Pennisetumcentrasiaticum)是禾本科(Gramineae)黍族(Trib. Paniceae)狼尾草屬(Pennisetum)多年生草種,具橫走根莖,稈直立,單生或叢生,產(chǎn)我國黑龍江、吉林、遼寧、內(nèi)蒙古、河北、山西、陜西、甘肅、青海、四川(西北部)、云南(北部)、西藏等省區(qū),多生于海拔800—4600 m山坡和較干燥之處[11]。白草是一種根莖型禾本科牧草,地下部可蔓延形成網(wǎng)狀地下根莖系統(tǒng)[12]而具有強(qiáng)大的擴(kuò)展能力[13]。目前,對于白草的研究有核型分析[14—15]、構(gòu)件生物量分配[16—17]、耐旱研究[13, 18—19]、營養(yǎng)價(jià)值[20—21]、栽培研究[22—23]以及生態(tài)修復(fù)[24—25]等。
雅魯藏布江是青藏高原上的最重要的河流,同時(shí)也是我國重要的國際河流之一[26],其發(fā)源于西藏自治區(qū)日喀則市仲巴縣海拔5590 m的杰馬央宗冰川[27],由西向東橫貫西藏南部,繞過喜馬拉雅山脈最東端的南迦巴瓦峰轉(zhuǎn)向南流,經(jīng)巴昔卡出中國境,而后進(jìn)入印度、孟加拉國,最后注入印度洋[28]。在雅魯藏布江流域,隨著經(jīng)度的增加,植被類型逐漸由荒漠草原向高山草原演變,進(jìn)而發(fā)展為灌木草原[29]。上游地區(qū)以蒿屬、針茅屬、狼尾草屬、嵩草屬等為主,是典型的荒漠草原群落[30]。中游地區(qū)除上述種類外,還包括薔薇科和豆科灌木,以高寒草地群落和灌木群落為主[31—32]。在雅魯藏布江中上游這一生態(tài)敏感區(qū)域開展野外樣方調(diào)查,可明確白草在該區(qū)域分布范圍、白草群落特征及其與環(huán)境因子的關(guān)系,為該流域植物多樣性評估和保護(hù)利用提供科學(xué)依據(jù),對確保青藏高原主體作為我國重要生態(tài)屏障有著十分重要的科學(xué)意義。
1 材料與方法
1.1 研究區(qū)概況
研究區(qū)位于西藏自治區(qū)雅魯藏布江上游日喀則市仲巴縣帕羊鎮(zhèn)至中游林芝市米林縣派鎮(zhèn),研究樣點(diǎn)經(jīng)度(83°44′16″—94°51′44″E),緯度(28°11′24″—29°6′1″N),平均海拔3890.65 m。
1.2 研究方法1.2.1 樣地設(shè)置
根據(jù)白草在西藏的分布范圍和生境特征,在雅魯藏布江中上游地區(qū)確定26個(gè)典型分布區(qū)域(圖1)。
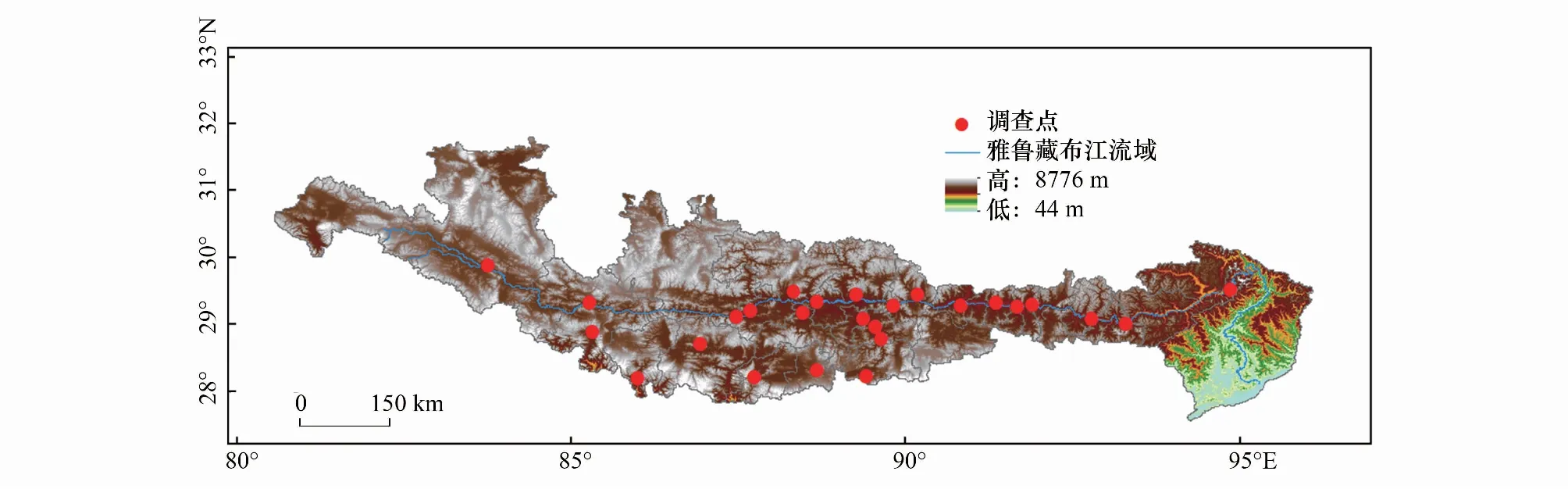
圖1 雅魯藏布江中上游白草主要分布區(qū)樣地Fig.1 Sampling sites of P. centrasiaticum communities in the upper and middle reaches of the Yarlung Zangbo River
通過路線踏察在研究區(qū)域選取白草分布較集中的樣地,利用手持GPS儀(eTrex Venture, Garmin, USA)記錄采樣點(diǎn)的經(jīng)度、緯度、海拔等信息,并根據(jù)經(jīng)緯度信息在International Water Management Institute(IWMI)在線氣候服務(wù)平臺(tái)(http://wcatlas.iwmi.org/Default.asp)獲取降雨量和平均溫度數(shù)據(jù)(表1)。
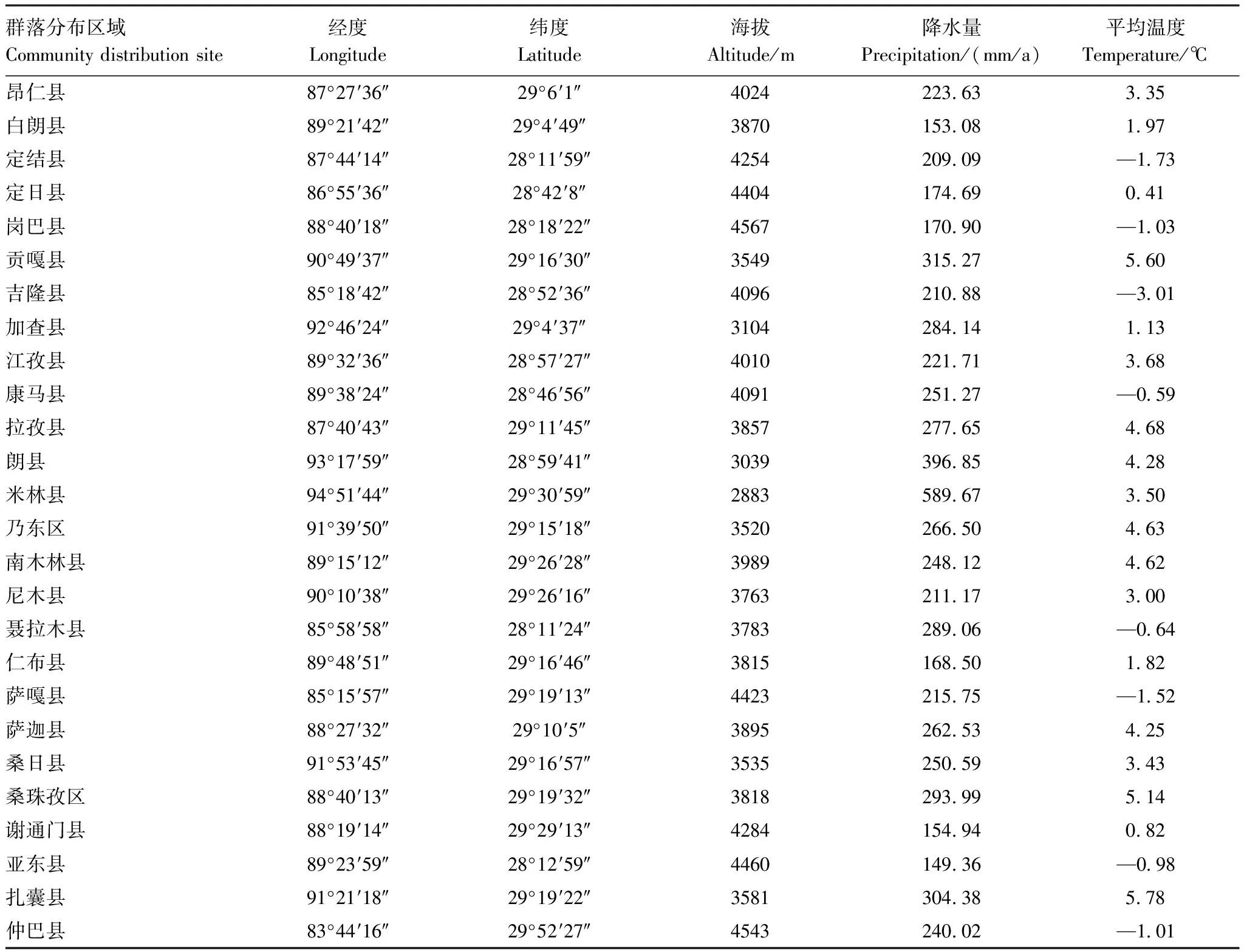
表1 各取樣點(diǎn)基本信息及氣候因子
1.2.2植物群落特征調(diào)查
2020年8—9月,在植物生長旺盛期,于每個(gè)樣地選擇群落結(jié)構(gòu)和組成分布均勻的區(qū)域,隨機(jī)選取3個(gè)50 cm×50 cm的調(diào)查研究樣方,調(diào)查群落內(nèi)各物種的高度、蓋度(對于樣方內(nèi)存在的矮小灌木,如砂生槐等,在本實(shí)驗(yàn)中,按照草本植物的調(diào)查方法處理)。分類齊地面剪取樣方內(nèi)各物種地上部分,帶回室內(nèi)于65℃下烘干至恒重,記錄干重,作為各物種的地上生物量。地上總生物量采用各物種地上生物量干重之和計(jì)算。
1.2.3植物功能群的劃分
植物功能群參照王長庭[33]及李里[34]的標(biāo)準(zhǔn)進(jìn)行劃分。根據(jù)拉瓊[35]等對雅魯藏布江上游源頭區(qū)至下游對河岸植物物種豐富度分布格局的研究結(jié)果將本研究中的植物功能群劃分為禾本科(Gramineae)、豆科(Leguminosae)、菊科(Asteraceae)和雜類草(Weeds)。
1.2.4地下生物量測定
地下生物量的測定采用土柱法,用特制的圓形鋼筒取樣。鋼筒直徑5 cm,高30 cm,端口削尖。地下生物量的取樣與地上生物量同步進(jìn)行,每個(gè)地上的生物量樣方內(nèi)設(shè)一個(gè)地下生物量樣方。每個(gè)樣方取3層,分0—10 cm,10—20 cm,20—30 cm。樣品按層裝入塑封袋中,標(biāo)記好帶回室內(nèi)。用80目篩沖洗、分離,將根系按層裝入紙袋中,在65℃烘箱內(nèi)烘干至恒重,稱其干重。根據(jù)土柱內(nèi)各層地下生物量測定值,按比例換算成單位面積地下生物量。
1.2.5土壤養(yǎng)分測定
用上述圓形鋼筒進(jìn)行土壤樣品取樣,每個(gè)樣方采集3鉆0—10 cm表層土,每個(gè)樣點(diǎn)的土樣混合裝入密封袋帶回實(shí)驗(yàn)室,自然風(fēng)干后去除雜草、根系、石塊等,過篩備用。土壤養(yǎng)分調(diào)查參考鮑士旦《土壤農(nóng)化分析》[36],土壤pH測定采用pH計(jì)測定,土壤含水量采用稱重法測定,土壤有機(jī)質(zhì)(SOM)采用重鉻酸鉀氧化法測定,土壤全氮(TNC)采用重鉻酸鉀—硫酸消化法測定,全磷(TPC)采用高氯酸—硫酸法測定,速效磷(APC)采用碳酸氫鈉浸提—鉬銻抗比色法測定,速效氮(ANC)采用擴(kuò)散法測定。
1.2.6數(shù)據(jù)分析
豐富度:
r=S
植物物種相對重要值計(jì)算:
Shannon-Weiner指數(shù)計(jì)算:
Simpson指數(shù)計(jì)算:
物種均勻度指數(shù)計(jì)算:
J=H/lnS
式中:S為所調(diào)查樣方內(nèi)物種總數(shù),RC為相對蓋度,RH為相對高度,RB為相對生物量。
用IBM SPSS Statistics 20 進(jìn)行單因素方差分析(One-way ANOVA)和Duncan多重比較(顯著水平0.05),并用于計(jì)算指標(biāo)間的Pearson線性相關(guān)系數(shù);采用Canoco 4.5軟件進(jìn)行白草群落特征與環(huán)境因子之間的冗余分析(Redundancy analysis, RDA)。用Microsoft Excel 2016對原始數(shù)據(jù)進(jìn)行匯總并進(jìn)行圖表的繪制,數(shù)據(jù)以平均值±標(biāo)準(zhǔn)誤差(Mean ± SE)來表示。
2 結(jié)果與分析
2.1 雅魯藏布江中上游地區(qū)白草主要分布區(qū)植物群落結(jié)構(gòu)組成和植物多樣性特征
從雅魯藏布江上游仲巴縣帕羊鎮(zhèn)至米林縣派鎮(zhèn)白草主要分布區(qū)26個(gè)樣地中,各白草群落共記錄到植物有35科96屬共119個(gè)物種。其中含種數(shù)較多的有禾本科(Gramineae)15屬16種,代表物種:白草、固沙草、垂穗披堿草(Elymusnutans)等;菊科(Asteraceae)14屬20種,代表物種:藏沙蒿、白蓮蒿、狗娃花(Heteropappushispidus)等;豆科(Leguminosae)9屬12種,代表物種:砂生槐、筆直黃耆(Astragalusstrictus)、黃華(Thermopsislupinoides)等。
根據(jù)樣地群落中物種重要值大小劃分為白草群落、藏沙蒿群落、砂生槐群落、白蓮蒿群落和固沙草群落5種群落類型,群落類型所在樣地和次優(yōu)勢種統(tǒng)計(jì)見表2。
經(jīng)過對社區(qū)家長教育的考察和梳理發(fā)現(xiàn),在社區(qū)教育發(fā)展的成長、成熟、壯大過程中,有幾個(gè)關(guān)鍵因素始終伴隨,即社會(huì)發(fā)展的歷史背景、政府的多方面支持、社區(qū)成員堅(jiān)持不懈的本真追求及多重社會(huì)力量的參與等[1]。這些干擾社區(qū)教育實(shí)施發(fā)展的因素進(jìn)一步反襯出社區(qū)家長教育的必要性。
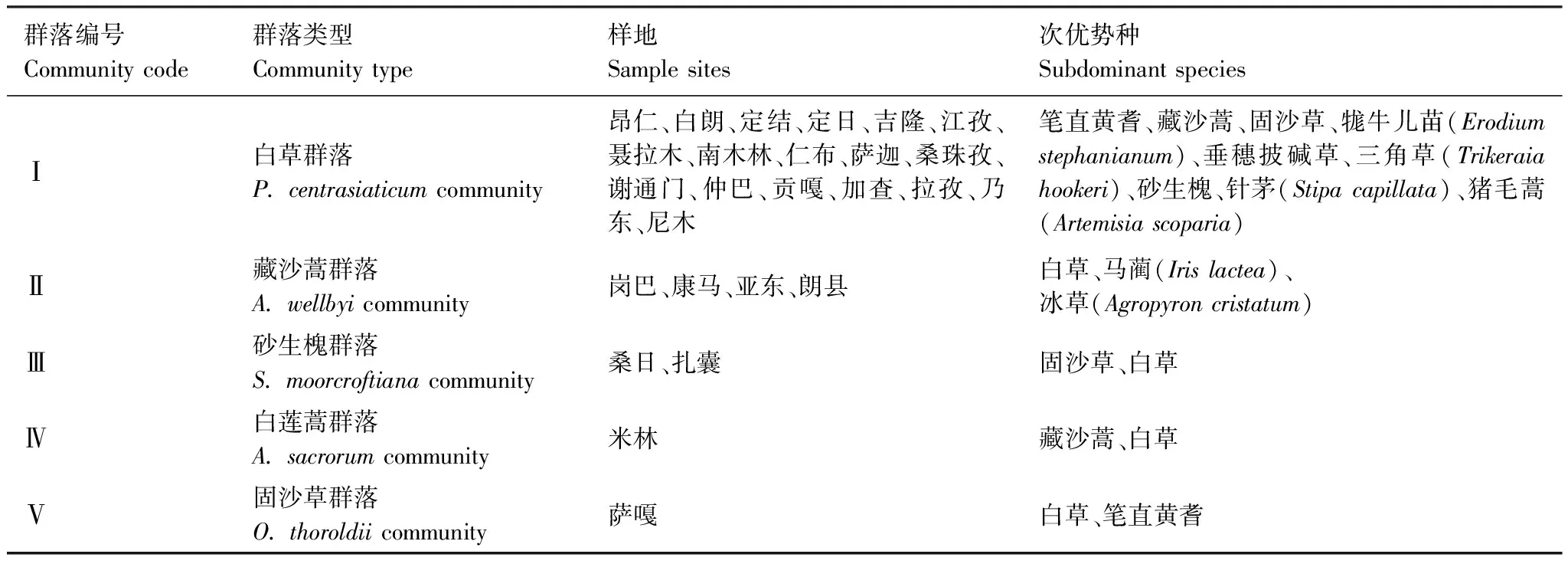
表2 白草主要分布區(qū)植物群落類型及其結(jié)構(gòu)組成
雅魯藏布江中上游白草主要分布區(qū)植物群落多樣性特征統(tǒng)計(jì)結(jié)果見表3。各白草群落物種豐富度變幅較大,桑日縣最低(S=12),江孜縣最高(S=37),植物物種數(shù)量相差25個(gè)。Shannon-Weiner指數(shù)(H)變幅為1.28(仁布縣)—2.30(米林縣);Simpson指數(shù)(D)變幅為0.52(拉孜縣)—0.86(米林縣);物種均勻度指數(shù)(J)變幅為0.54(昂仁縣)—0.80(朗縣)。
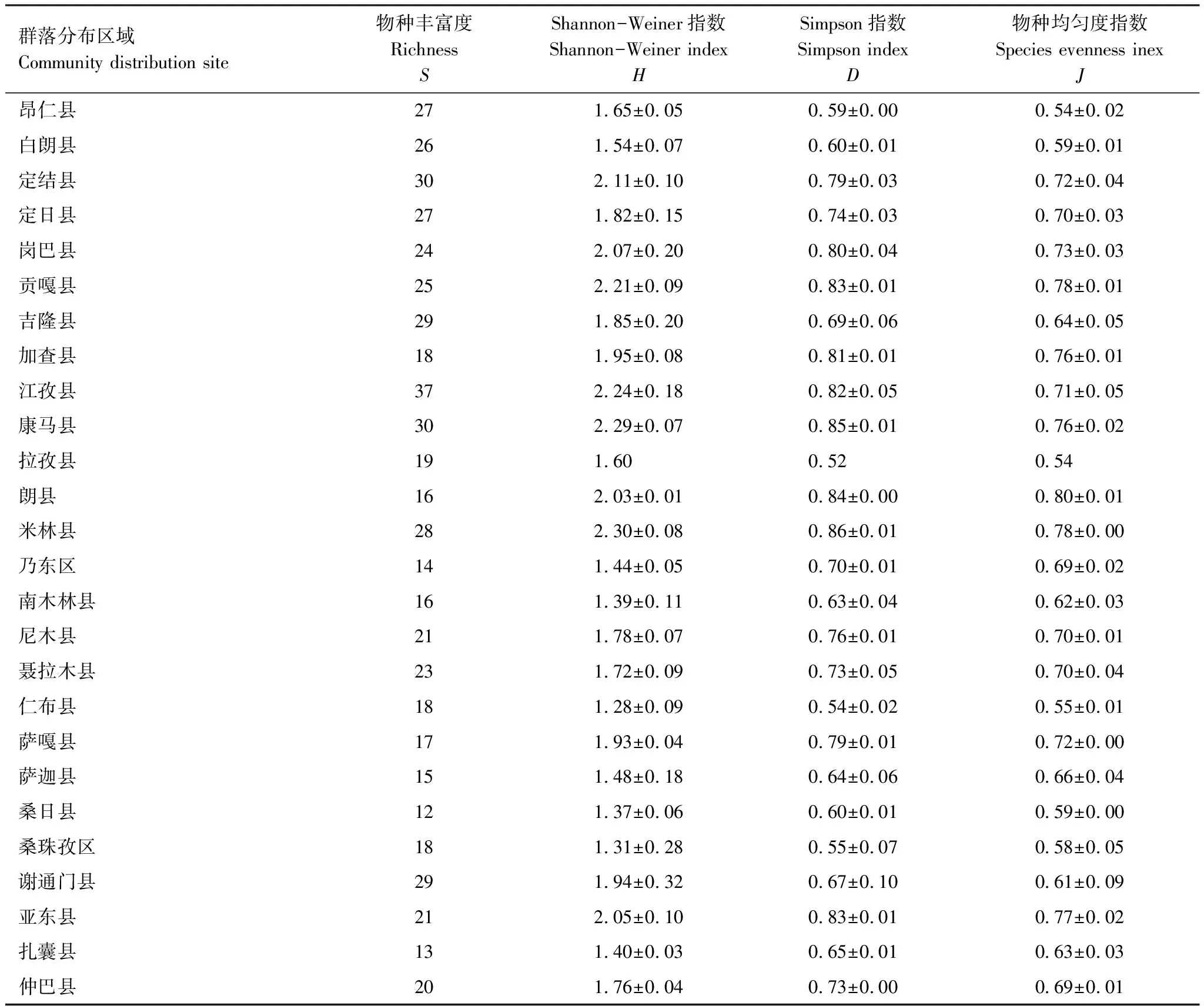
表3 白草主要分布區(qū)植物群落物種多樣性特征
對各群落內(nèi)白草高度、蓋度、生物量和重要值進(jìn)行統(tǒng)計(jì)分析,結(jié)果見表4。白草平均高度為64.22 cm,變幅為26.97 cm(仲巴縣)—96.38 cm(米林縣);白草平均蓋度為56.57%,變幅為10.33%(桑日縣)—90.67%(薩迦縣);白草平均生物量為250.87 g/m2,變幅為48.77 g/m2(桑日縣)—507.84 g/m2(尼木縣);白草平均重要值為39.38%,變幅為11.88%(桑日縣)—67.52%(拉孜縣)。
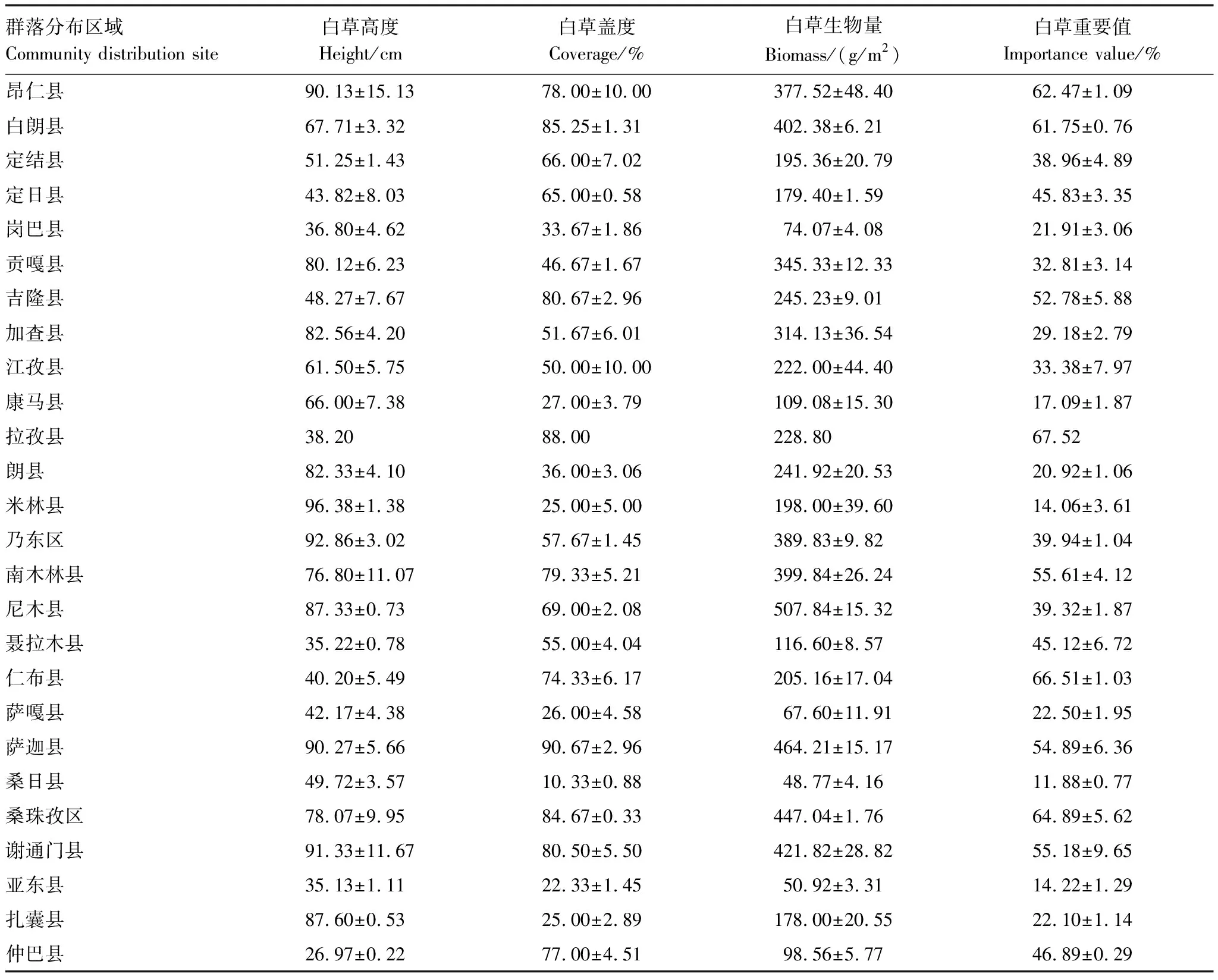
表4 白草主要分布區(qū)植物群落物種多樣性特征
2.2 群落地下生物量分析
將群落地下0—10 cm,10—20 cm,20—30 cm土層地下生物量分別統(tǒng)計(jì),結(jié)果如表5。各群落0—10 cm土層中,地下生物量均為最大,隨土層的加深,地下生物量降低。各群落中,0—10 cm、10—20 cm、20—30 cm土層和總的地下生物量平均為1969.30 g/m2,921.77 g/m2,483.16 g/m2和3374.23 g/m2;各土層地下生物量占比分別為60.38%,26.36%和13.26%。
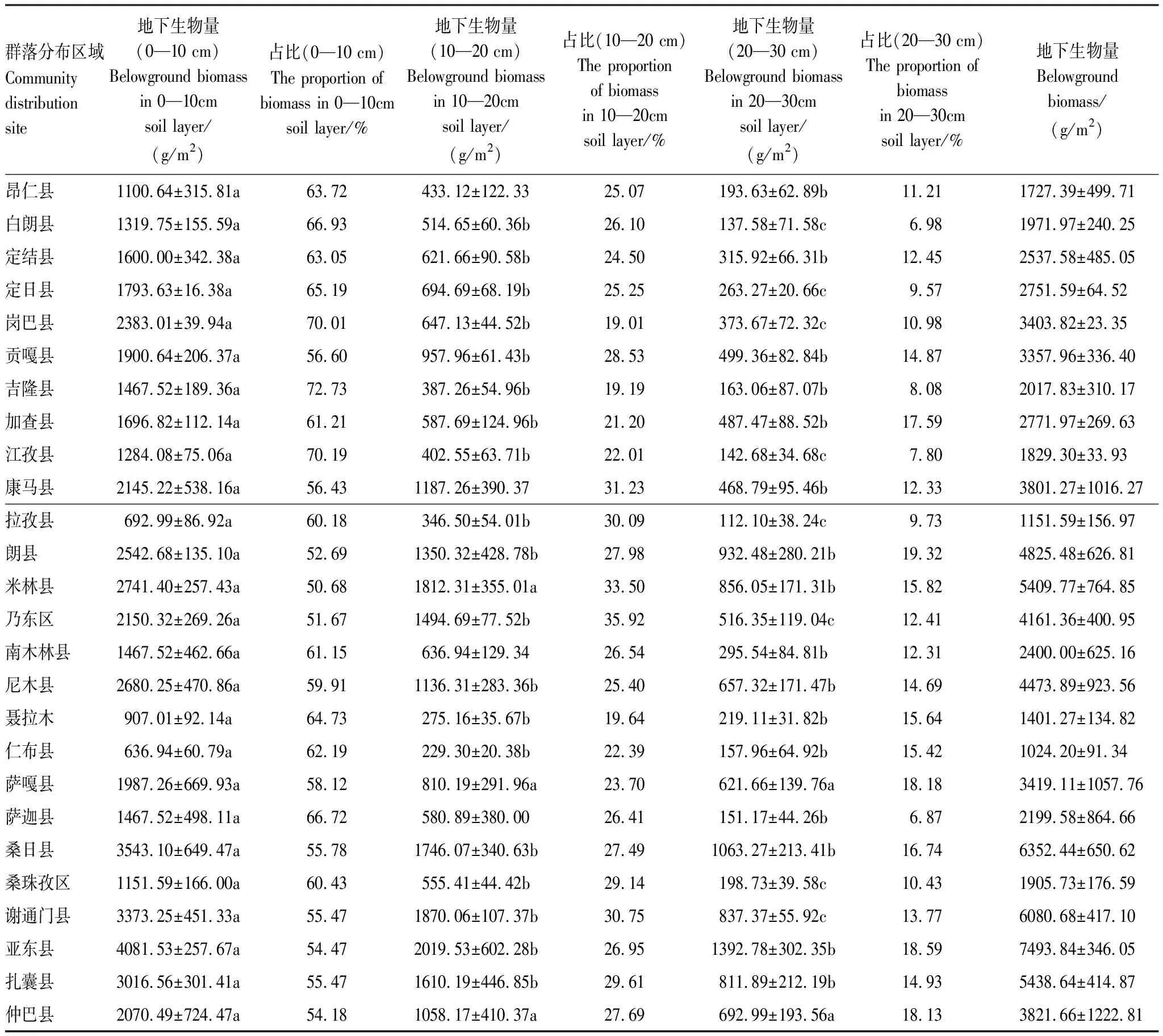
表5 白草主要分布區(qū)不同土層地下生物量及各層生物量占比
各樣地中,亞東縣地下生物量最高,達(dá)到7493.84 g/m2,仁布縣地下生物量最低,為1024.20 g/m2。在0—10 cm土層,亞東縣生物量最高,為4081.53 g/m2,吉隆縣生物量占比最高,為72.73%;在10—20 cm土層中,亞東縣生物量最高,為2019.53 g/m2,乃東區(qū)生物量占比最高,為35.92%;在20—30 cm土層中,亞東縣生物量最高,為1392.78 g/m2,朗縣生物量占比最高,為19.32%。
2.3 白草種群特征及群落特征與主要環(huán)境因子間的相關(guān)性
白草種群特征與主要環(huán)境因子間相關(guān)關(guān)系如表6所示。白草高度與海拔(r=-0.60),白草高度與土壤pH(r=-0.53),白草生物量與土壤pH(r=-0.55)呈極顯著負(fù)相關(guān)關(guān)系(P<0.01)。白草高度與全磷含量(r=0.42),白草蓋度與速效氮含量(r=0.43),白草蓋度與有機(jī)質(zhì)含量(r=0.41),白草生物量與全磷含量(r=0.40),白草重要值與速效氮含量(r=0.40),白草重要值與有機(jī)質(zhì)含量(r=0.46)呈顯著正相關(guān)關(guān)系。
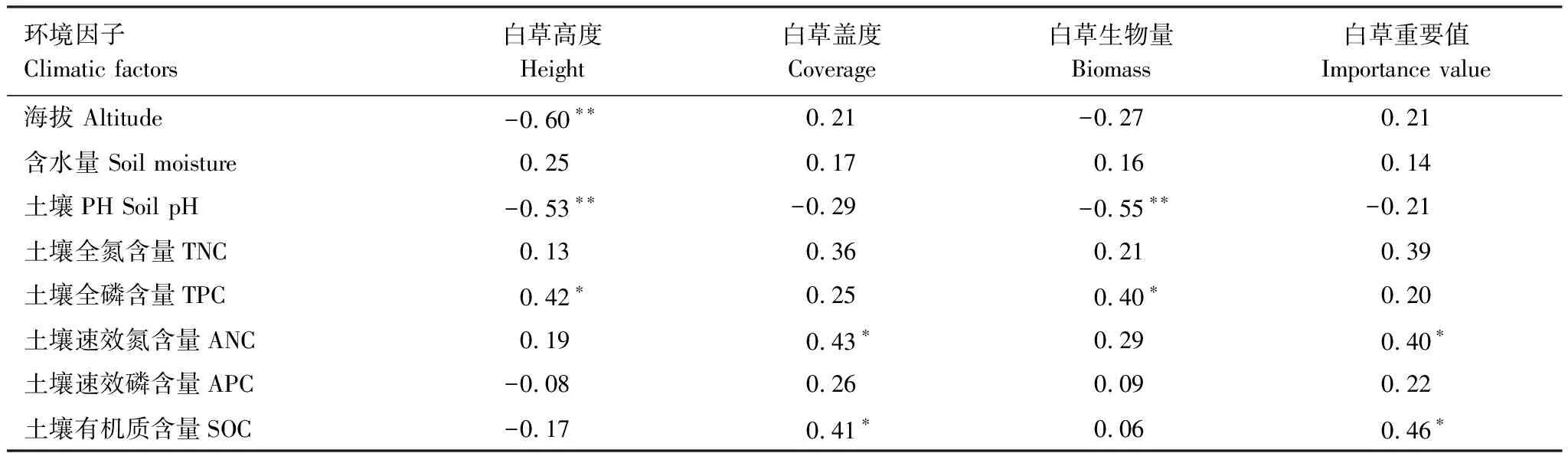
表6 白草種群特征與環(huán)境因子相關(guān)分析
冗余分析(RDA)是一種回歸分析與主成分分析相結(jié)合的排序方法,是多響應(yīng)變量回歸分析的拓展[37],能夠整體反映環(huán)境因子與白草群落常見物種組成和個(gè)體分布之間的關(guān)系。將白草和群落中常見的12種植物(白草、砂生槐、筆直黃耆、豬毛蒿、菊葉香藜Chenopodiumfoetidum、狗娃花、風(fēng)毛菊Saussureajaponica、固沙草、蒲公英Taraxacummongolicum、平車前Plantagodepressa、毛瓣棘豆Oxytropissericopetala、垂穗披堿草、藏沙蒿)的重要值為基礎(chǔ)數(shù)據(jù)建立響應(yīng)變量矩陣,以9個(gè)環(huán)境因子(7個(gè)土壤養(yǎng)分含量、年平均溫度、年平均降雨量)作為解釋變量進(jìn)行RDA分析,結(jié)果見圖2。
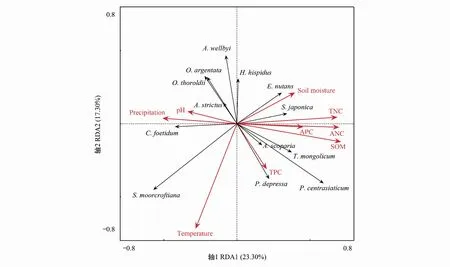
圖2 白草主要分布區(qū)常見物種重要值與環(huán)境因子的RDA排序Fig.2 RDA ordination of environmental factors and importance value of common species in P. centrasiaticum distribution areas
軸1和軸2共解釋全部方差的40.60%,在環(huán)境—群落的解釋中起主導(dǎo)作用。其中RDA1軸與土壤pH、年平均降雨量、全氮含量、速效氮含量、速效磷含量、有機(jī)質(zhì)含量、土壤含水量有較強(qiáng)的關(guān)系性,單獨(dú)解釋了方差的23.30%,說明土壤養(yǎng)分因子及降雨量對白草群落結(jié)構(gòu)產(chǎn)生影響。RDA2軸與年平均溫度、全磷含量有較強(qiáng)相關(guān)性,單獨(dú)解釋變異的17.30%。總體上,白草主要分布區(qū)土壤pH與其它土壤理化特征之間呈負(fù)相關(guān),白草重要值和土壤含水量、全氮、全磷、速效氮、速效磷、有機(jī)質(zhì)含量有較強(qiáng)的相關(guān)性。
2.4 白草主要分布區(qū)聚類分析
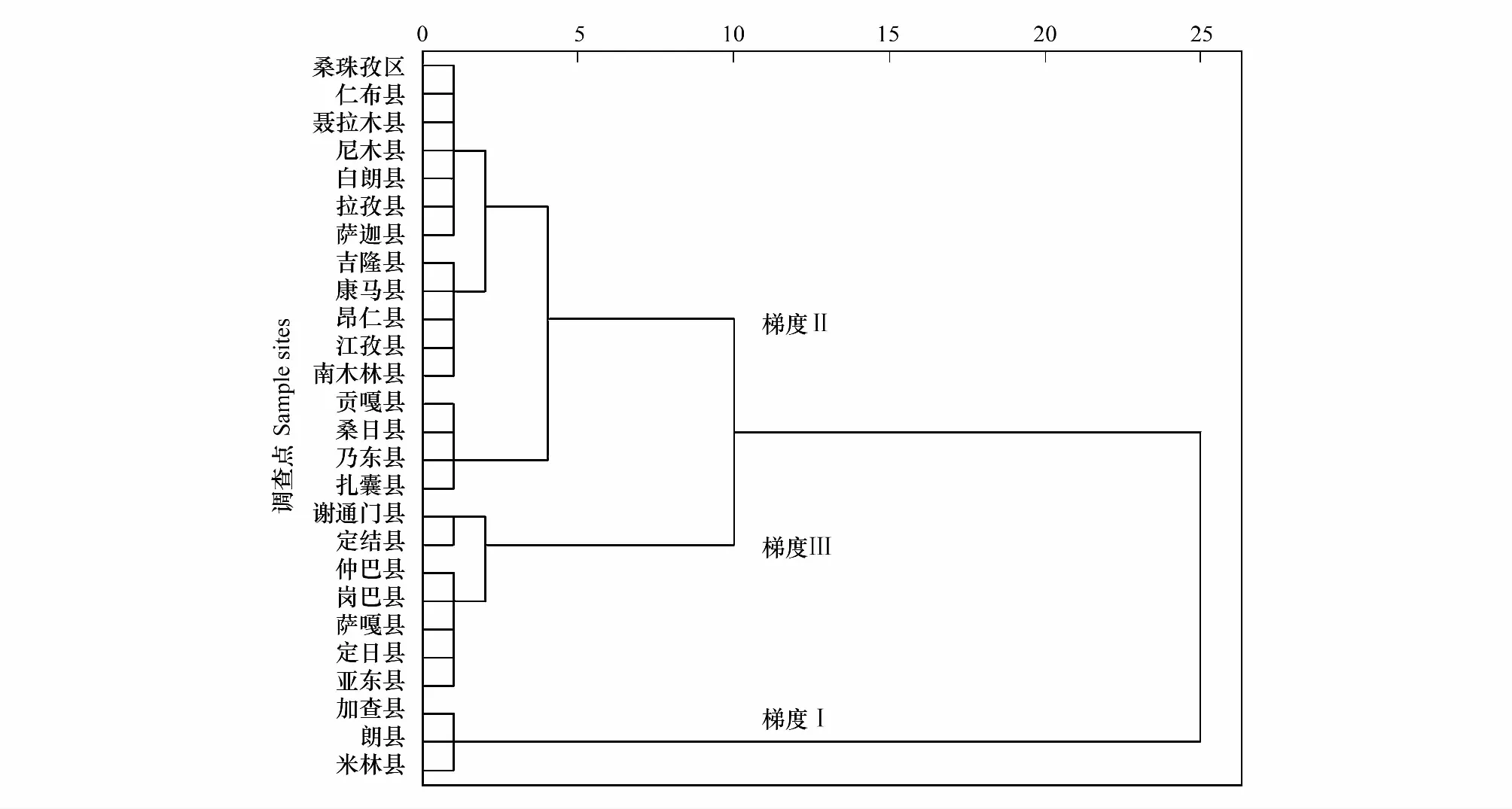
圖3 雅魯藏布江中上游白草主要分布區(qū)聚類圖Fig.3 Clusters of P. centrasiaticum distribution areas in the middle and upper reaches of the Yarlung Zangbo River
梯度Ⅰ(2883—3104 m,平均海拔3008.67 m),包含加查縣、朗縣和米林縣3個(gè)樣點(diǎn);梯度Ⅱ(3520 m—4096 m,平均海拔3824.75 m),包含薩迦縣、桑珠孜區(qū)、南木林縣等16個(gè)樣點(diǎn);梯度Ⅲ(4254 m—4567 m,平均海拔4419.29 m),包含謝通門縣、定日縣、定結(jié)縣等7個(gè)樣點(diǎn)。
2.4.1不同海拔植株功能群特征
不同植物功能群在不同海拔梯度的高度、蓋度、地上生物量和重要值如圖4所示。26個(gè)樣地不同功能群平均高度順序?yàn)椋汉瘫究?32.78 cm)>豆科(20.11 cm)>菊科(17.00 cm)>雜類草(9.16 cm)。各功能群的平均蓋度順序?yàn)椋汉瘫究?65.50%)>菊科(14.25%)>豆科(11.01%)>雜類草(9.01%)。各功能群平均地上生物量順序?yàn)椋憾箍?327.78 g/m2)>禾本科(305.15 g/m2)>菊科(255.55 g/m2)>雜類草(67.23 g/m2)。各功能群平均重要值順序?yàn)椋汉瘫究?52.51%)>菊科(16.90%)>豆科(16.04%)>雜類草(14.55%)。
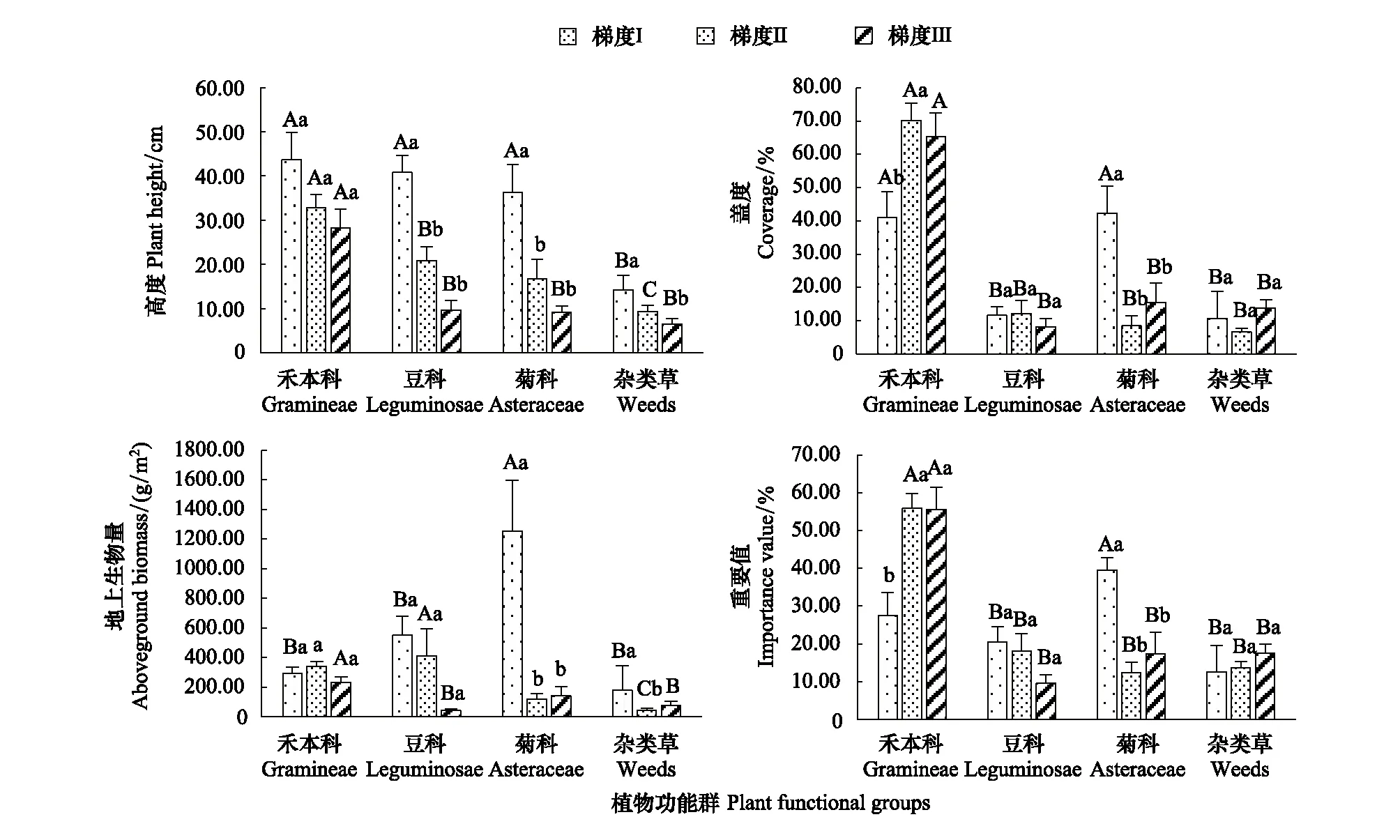
圖4 不同海拔梯度植株功能群特征Fig.4 The Characteristics of plant functional groups in different altitude gradients不同大寫字母表示相同海拔不同植物功能群特征差異顯著,不同小寫字母表示相同植物功能群特征在不同海拔差異顯著(P<0.05)
豆科、禾本科、菊科和雜類草功能群的高度均隨著海拔上升而顯著降低(P<0.05)。禾本科功能群的蓋度在梯度Ⅱ(70.22%)顯著高于梯度Ⅰ(40.93%)(P<0.05),與梯度Ⅲ(65.27%)差異不顯著;菊科功能群的蓋度(42.29%)在梯度Ⅰ最大,顯著高于梯度Ⅱ(8.44%)和梯度Ⅲ(15.52%)(P<0.05);豆科和雜類草功能群的蓋度在3個(gè)海拔下差異不顯著。菊科功能群的地上生物量在梯度Ⅰ(1255.58 g/m2)顯著高于梯度Ⅱ(118.42 g/m2)和梯度Ⅲ(140.38 g/m2)(P<0.05),雜類草功能群的地上生物量在梯度Ⅰ(178.01 g/m2)顯著大于梯度Ⅱ(42.45 g/m2)(P<0.05),與梯度Ⅲ(76.40 g/m2)差異不顯著;豆科和禾本科功能群地上生物量在3個(gè)海拔下差異不顯著。禾本科功能群重要值在梯度Ⅱ(55.85%)和梯度Ⅲ(55.58%)差異不顯著,顯著高于梯度Ⅰ(27.50%)(P<0.05);菊科功能群的重要值在梯度Ⅰ(39.53%)顯著高于梯度Ⅱ(12.47%)和梯度Ⅲ(17.32%)(P<0.05);豆科和雜類草功能群的重要值在3個(gè)海拔下差異不顯著。
2.4.2不同海拔植物群落地下生物量分配
不同海拔白草群落的地下生物量占總生物量比例的計(jì)算結(jié)果如圖5所示。隨著海拔的增大,地下生物量占比顯著增加(P<0.05)。在梯度Ⅲ,地下生物量占比為89.08%,顯著高于梯度Ⅱ(77.37%)和梯度Ⅰ(65.44%)(P<0.05),梯度Ⅱ地下生物量占比顯著高于梯度Ⅰ(P<0.05)。
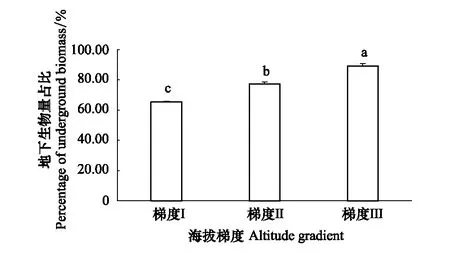
圖5 不同海拔地下生物量占比 Fig.5 Percentage of underground biomass in different altitude gradients 不同小寫字母表示地下生物量占比在不同海拔差異顯著(P<0.05)
2.4.3不同海拔植物群落多樣性特征
植物群落多樣性在各海拔梯度的變化見表7,群落物種豐富度在3個(gè)海拔梯度差異不顯著,其變化趨勢為:梯度Ⅲ(24.00)>梯度Ⅱ(21.44)>梯度Ⅰ(20.67)。群落多樣性指數(shù)(H、D、J)在不同海拔的變化趨勢為:梯度Ⅰ>梯度Ⅲ>梯度Ⅱ,梯度Ⅰ顯著大于梯度Ⅱ(P<0.05),梯度Ⅲ和另2個(gè)梯度的差異不顯著。

表7 不同海拔植物群落多樣性指數(shù)
3 討論
3.1 雅魯藏布江中上游白草主要分布區(qū)植物功能群特征
功能群被認(rèn)為是與系統(tǒng)的某種功能直接相關(guān)的物種群,功能群內(nèi)物種對系統(tǒng)的作用有很大相似性,因而用功能群數(shù)作為物種多樣性的指標(biāo)研究多樣性與生態(tài)系統(tǒng)功能的關(guān)系值得肯定[33, 38]。用功能群以解釋不同演替梯度或生境中物種多樣性格局的變化及與生態(tài)系統(tǒng)功能的關(guān)系,使復(fù)雜的問題較為簡化且更具說服力[34, 39]。因研究角度的不同,不同科學(xué)家對植物功能群的劃分也不同[34, 40—41]。拉瓊[35]研究表明:雅魯藏布江上游源頭區(qū)至下游出現(xiàn)頻度最高的前三大科分別是菊科、禾本科和豆科,這與本研究的結(jié)果相同。以白草為代表的禾本科功能群葉面積和葉位相對較大,在雅魯藏布江中上游26個(gè)樣地中有最大的高度、蓋度和重要值,在生長方面占據(jù)光照和養(yǎng)分的優(yōu)勢[40, 42—43],抑制了豆科、菊科和雜類草的生長。功能群之間具有補(bǔ)償作用,存在生態(tài)位重疊的現(xiàn)象,當(dāng)一種功能群的數(shù)量或生產(chǎn)力下降時(shí),會(huì)為另一些功能群提供更大的空間,使之占據(jù)更多的資源[34]。梯度Ⅰ的禾本科功能群蓋度低(40.93%),菊科功能群迅速進(jìn)行補(bǔ)償,提高其高度(36.34 cm)、蓋度(42.29%)、生物量(1255.58 g/m2)和重要值(39.53%)。其它研究也表明禾本科牧草蓋度的降低將導(dǎo)致毒害草和雜類草增多;禾本科牧草蓋度的提升,使得雜類草減少[40—44]。植物的這種生態(tài)補(bǔ)償作用是生命實(shí)體對環(huán)境變化和干擾的一種響應(yīng)對策,通過功能群之間的相互作用來實(shí)現(xiàn)植物功能群對群落穩(wěn)定性的影響[34]。
3.2 雅魯藏布江中上游白草主要分布區(qū)群落結(jié)構(gòu)特征與環(huán)境解釋
植物之間均存在著廣泛而復(fù)雜的相互作用[45],植物種群結(jié)構(gòu)可反映種群的數(shù)量動(dòng)態(tài)、發(fā)展趨勢與環(huán)境之間的相互關(guān)系[46],也可揭示植物種群與生境相適應(yīng)的結(jié)果,對預(yù)測群落未來發(fā)展趨勢具有重要意義[47—48]。植物種群多樣性特征及其空間分布格局能夠反映環(huán)境對種群內(nèi)個(gè)體生長和生存的影響,也能反映和指示植物種群的生態(tài)適應(yīng)對策[49]。在草地發(fā)展過程中,受到多種環(huán)境因素的影響,其中降水和溫度作為重要的氣候因子,成為草地生長發(fā)育因素研究中的重點(diǎn)[50—51]。受海拔和經(jīng)度的影響,雅魯藏布江流域自西向東形成濕熱增加的梯度,植物多樣性沿該梯度逐漸增加[29]。拉瓊[35]等沿海拔梯度從雅魯藏布江上游源頭區(qū)至下游記錄的維管束植物238種,物種分布格局呈現(xiàn)出物種豐富度在下游最高、中游偏低、上游略上升的趨勢。另外,在拉薩河上游的研究[52]發(fā)現(xiàn)物種豐富度在山腰低,在山腳和山頂高,其成因被認(rèn)為是半山腰高密度灌木使物種豐富度降低。在本實(shí)驗(yàn)中,白草主要分布區(qū)共記錄植物物種數(shù)119個(gè),隨海拔上升,群落豐富度增加。在白草主要分布區(qū),禾本科植物在不同的植物功能群中高度和蓋度最大,占據(jù)光照和養(yǎng)分的優(yōu)勢,導(dǎo)致雜草物種的排除[53],使本實(shí)驗(yàn)記錄的植物物種總數(shù)低于拉瓊的調(diào)查。梯度Ⅰ溫度高、降雨多,白蓮蒿和藏沙蒿等成為白草主要分布區(qū)的優(yōu)勢種或者次優(yōu)勢種,相對高大的菊科植物降低了樣方內(nèi)物種豐富度[54],使其物種豐富度低于梯度Ⅱ。同理,梯度Ⅱ的物種豐富度低于梯度Ⅲ,這是物種之間對光照競爭的結(jié)果[55]。白草主要分布區(qū)土壤pH與其它土壤理化特征呈負(fù)相關(guān),與羅巧玉[45]對發(fā)草適生地土壤以及王文穎[56]對高寒藏嵩草草甸土壤的研究結(jié)果相同。土壤pH與海拔呈正相關(guān)關(guān)系,禾本科和雜類草植物數(shù)量隨土壤pH值逐漸增加[57],因而白草群落豐富度隨海拔上升增加。白草主要分布區(qū)禾本科植物的重要值為梯度Ⅱ>梯度Ⅲ>梯度Ⅰ,而群落多樣性指數(shù)呈現(xiàn)相反趨勢。這與周選博[43]和張峰[58]研究得出禾本科植物重要值增加,將導(dǎo)致Shannon-Weiner指數(shù)、Simpson指數(shù)和物種均勻度指數(shù)變小這一結(jié)果相同。
白草主要分布區(qū)各群落中,地下生物量隨土層深度增加而遞減,在0—10 cm土層地下生物量占比達(dá)到60.38%。表層通氣條件較好,地下生物量大,隨著土壤深度的增加,土壤溫度、含水量和通氣條件變劣,地下生物量相應(yīng)減少[59]。雅魯藏布江中上游白草群落的地下生物量大都分布在表土層中以獲取更多的熱量、水分和礦質(zhì)營養(yǎng),表明高原植物對嚴(yán)酷環(huán)境有適應(yīng)性。青藏高原高寒地區(qū),強(qiáng)輻射、生長季節(jié)短等因素阻礙植物生長[60],寒冷天氣降低植物細(xì)胞分裂的速度,植物難以發(fā)育成大個(gè)體[61],導(dǎo)致高海拔地區(qū)植物個(gè)體矮小,地上生物量降低;同時(shí),以白草為代表的多年生植物常將更多生物量投資到根部[16, 62],使雅魯藏布江中上游流域白草主要分布區(qū)的地下生物量占比隨海拔上升顯著增加。
4結(jié) 論
以上研究對雅魯藏布江中上游地區(qū)白草主要分布區(qū)植物群落特征進(jìn)行調(diào)查,發(fā)現(xiàn)白草主要分布區(qū)有119個(gè)植物物種,屬35科96屬,表土層地下生物量占比最高。RDA分析表明白草重要值和土壤含水量、全氮、全磷、速效氮、速效磷、有機(jī)質(zhì)含量有較強(qiáng)的相關(guān)性。聚類分析揭示4個(gè)功能群的高度均隨海拔上升而下降,群落物種豐富度和群落地下生物量隨海拔上升而占比增加。以上結(jié)果反映出白草是雅魯藏布江中上游廣泛分布的鄉(xiāng)土草種,其群落特征多樣,與環(huán)境因子有較強(qiáng)的相關(guān)性。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18
中華詩詞(2022年6期)2022-12-31 06:41:24
科學(xué)大眾(中學(xué))(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
中國科技論壇(2017年7期)2017-07-25 08:49:53
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學(xué)周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學(xué)周刊·少年版(2015年1期)2015-07-07 17:15:12
少兒科學(xué)周刊·兒童版(2015年1期)2015-07-07 04:12:52
