煤矸石山不同植物根際土壤AM真菌群落多樣性分析
2022-08-31 03:22:42甄莉娜劉麗珍王潤梅
草地學報 2022年8期
關鍵詞:植物
甄莉娜, 劉麗珍, 牛 艷, 李 俠, 李 朕, 吳 娜, 王潤梅
(山西大同大學生命科學學院應用生物技術研究所, 山西 大同 037009)
煤矸石是煤礦開采過程中產生的固體廢棄物,矸石的長期堆積,不僅占用耕地,甚至會發生自燃,產生CO,SO2等酸性氣體,污染礦區空氣、土壤和水源[1],使礦區生態環境被嚴重破壞[2]。
AM真菌屬球囊菌門專性營養共生真菌,存在于絕大多數生態系統中[3]。AM真菌能與90%以上的陸生和水生植物根系形成共生體,參與植物的生理代謝過程[4]。在矸石山恢復初期,植物與AM真菌存在良好的共生關系[5],AM真菌以其龐大的菌絲網絡系統將植物與土壤有機地聯合起來,能夠促進宿主植物對土壤氮、磷、鉀等礦質元素及水分的吸收,改善植物的營養狀況[6]。不僅能提高植物的生物量,還能夠改良礦區土壤的理化性狀、提高土壤生產力[7]。對煤矸石山礦區植被演替、生態系統穩定以及修復重金屬污染土壤具有重要意義。
AM真菌群落組成受到多種因素的影響,如海拔、降水量、植物類型、土壤pH值、鹽度、粒徑、養分含量、矸石山高度等[8-11]。AM真菌群落組成、豐富度及多樣性指數等在不同植物根際土壤中存在差異[9]。植物、土壤和菌根真菌之間的相互關系,對礦區等受擾動土壤生態系統功能的恢復具有重要意義。煤矸石山較高的AM真菌多樣性,可能會緩解外界干擾對矸石山植物多樣性造成的影響。
環境和共生植物種類影響了不同類群AM真菌群落的多樣性。球囊霉屬(Glomus)在不同生態環境中均是優勢屬,在所有采樣點的出現頻率和相對豐度最大[12],有研究發現根內球囊霉、球囊霉和隱球囊霉是半干旱礦區常見的AM真菌[13]。禾本科植物廣泛存在于各個生態系統中,在草地群落組成的出現率和豐富度均居首位[14]。有研究發現煤矸石礦區周邊禾本科植物最多,早期修復植被均能與AM真菌形成良好的共生關系[6]。菊科植物種類多,植株富含脂肪,可形成菌根[15],其蓋度的增加,對土壤AM真菌的群落結構和多樣性有重要的影響力[16];藜科植物是常見的非菌根植物,主要分布在荒漠和半荒漠草地,于宏清研究表明大部分藜科植物在鹽堿地都存在菌根侵染現象,AM真菌可以顯著提高藜科植物生物量,提高抗鹽脅迫和養分吸收的能力[17]。
AM真菌對煤礦區植物群落的恢復重建有重要的促進作用[18],AM真菌群落多樣性的增加可以提高植物群落多樣性,進而提高生態系統的穩定性[19],以往研究主要集中在菌根真菌對礦區植物生長狀況、養分吸收、重金屬脅迫、細菌和真菌多樣性等方面[20-21],而關于矸石山植物根際AM真菌多樣性的研究較少。本文以煤矸石山不同植物類型土壤根際AM真菌為研究對象,檢測不同恢復階段優勢植物AM真菌群落的多樣性,比較不同植物根際土壤理化性質、AM真菌群落組成、相對豐度及多樣性指數間的差異,為煤矸石山的生態恢復提供理論和實踐依據。
1 材料與方法
1.1 采樣地概況
大同市晉華宮礦是大同煤礦集團下屬的國有大型礦井之一,坐落于山西省北部,113°13′E,40°10′N,距大同12.5 km,礦區內有豐富的礦產資源,可采儲量1.5億t。地處低山丘陵黃土地貌景觀,井田內部為黃土覆蓋,屬中溫帶大陸性氣候,干燥少雨,降水量集中在7,8,9三個月,年平均最大降水量為628.3 mm,年溫差大,平均氣溫為5.1℃。
1.2 樣品采集
在大同市晉華宮礦煤矸石山區隨機采樣,于7月份進行矸石山不同坡向植物調查,設置草本植物樣方為1 m×1 m,共18個,在每個樣方內選取矸石山常見植物取樣,主要包括禾本科狗尾草(Setariaviridis)、虎尾草(Chlorisvirgata)、草地早熟禾(Poapratensis),豆科植物紫花苜蓿(Medicagosativa)、黃花草木樨(Melilotusofficinalis)、山野豌豆(Viciaamoena),藜科植物灰綠藜(Chenopodiumglaucum)、豬毛菜(Salsolacollina)、地膚(Kochiascoparia),菊科植物蒼耳(Xanthiumsibiricum)、黃花蒿(Artemisiaannua)和大籽蒿(Artemisiasieversiana)。利用抖根法采集根際土壤,抖動植物根系,去除根表面附著的大部分土壤,只保留根周圍1 mm左右的土,利用無菌生理鹽水洗下,離心,收集根際土。幼嫩根系保存于FAA(福爾馬林5 mL、冰醋酸5 mL、70%酒精90 mL)固定液中,在實驗室中進行根系侵染狀況檢測。另根際土用冰盒儲存帶回實驗室,在-80℃保存。
1.3 AMF Illumina MiSeq測序
土壤叢枝菌根真菌群落多樣性委托“美吉生物科技有限公司”檢測,根據 E.Z.N.A.?soil DNA kit (Omega Bio-tek,Norcross,GA,U.S.) 說明書進行微生物群落總DNA抽提,利用瓊脂糖凝膠電泳及NanoDrop2000技術,分別檢測提取DNA的質量、濃度與純度;利用引物擴增叢枝菌根真菌基因V4-V5高變區,測序區ITSrRNA的擴增引物序列為:AMV4.5 NF (F)-AAGCTCGTAGTTGAAT-TTCG、AMDGR(R)-CCCAACTATCCCTATT AATCAT,使用PCR(GeneAmp 9700)熱循環系統在95℃下循環30 s,在55℃下循環30 s,在72℃下循環45 s,共進行32個循環;在72℃下進行10 min的延伸,每個樣本3個重復。
同一樣本的PCR產物混合后,通過瓊脂糖凝膠檢測PCR產物,利用AxyPrep DNA凝膠回收試劑盒純化回收產物,并用QuantusTMFluorometer定量檢測回收產物。使用NEXTflexTMDNA快速建庫試劑盒進行建庫,包括接頭鏈接、篩選去除接頭自連片段、PCR擴增富集文庫模板、磁珠回收PCR產物、獲得文庫。利用Illumina公司的Miseq PE300平臺進行測序。土壤叢枝菌根真菌群落多樣性委托“美吉生物科技有限公司”檢測。
1.4 土壤理化性質檢測
使用pH計測定土壤pH值,利用重鉻酸鉀-濃硫酸氧化比色法測定土壤中有機質含量(Organic matter,OM),堿解擴散法測定土壤速效氮含量(Averlable nitrogen,AN),碳酸氫鈉浸提-鉬銻抗比色法測定土壤速效磷(Available phosphorus,AP),乙酸銨浸提-火焰光度計法測定速效鉀含量(Available potassium,AK)[22-23]。
1.5 數據處理
采用SPSS 22.0(New York,USA)軟件對不同植物類型數據進行單因素方差分析,比較檢驗煤矸石不同植物類型AM真菌土壤理化性質和群落多樣性的差異,對Sobs指數、Chao1指數、Ace指數、Shannon指數、Simpson多樣性指數進行分析,以上統計分析均在SPSS 22.0(New York,USA)軟件中完成。用R軟件(version2.15.3)分析和生成相對豐度柱形圖、NMDS圖和差異熱圖,分析不同植物類型AM真菌群落相對豐度,對土壤理化性質、多樣性指數、豐富度指數等進行冗余分析(RDA),利用Gephi軟件繪制網絡關系圖。
2 結果與分析
2.1 AM真菌群落稀釋性曲線分析
基于97%的序列相似度水平上,共得到4個科植物28個根系樣品的AM真菌稀釋曲線。由圖1可知,豆科和菊科的土壤真菌樣品的稀釋曲線都趨于平緩,OTU數量隨探測深度的增加不再增加,說明樣品OTU覆蓋度已接近飽和,能夠反映土壤中的真菌群落結構組成。藜科、禾本科土壤樣品中叢枝菌根真菌OTU數目相對較多,藜科的稀釋曲線仍有上升趨勢,未達到飽和狀態,表明如果繼續增加探測深度,OTU數目可能還會上升。
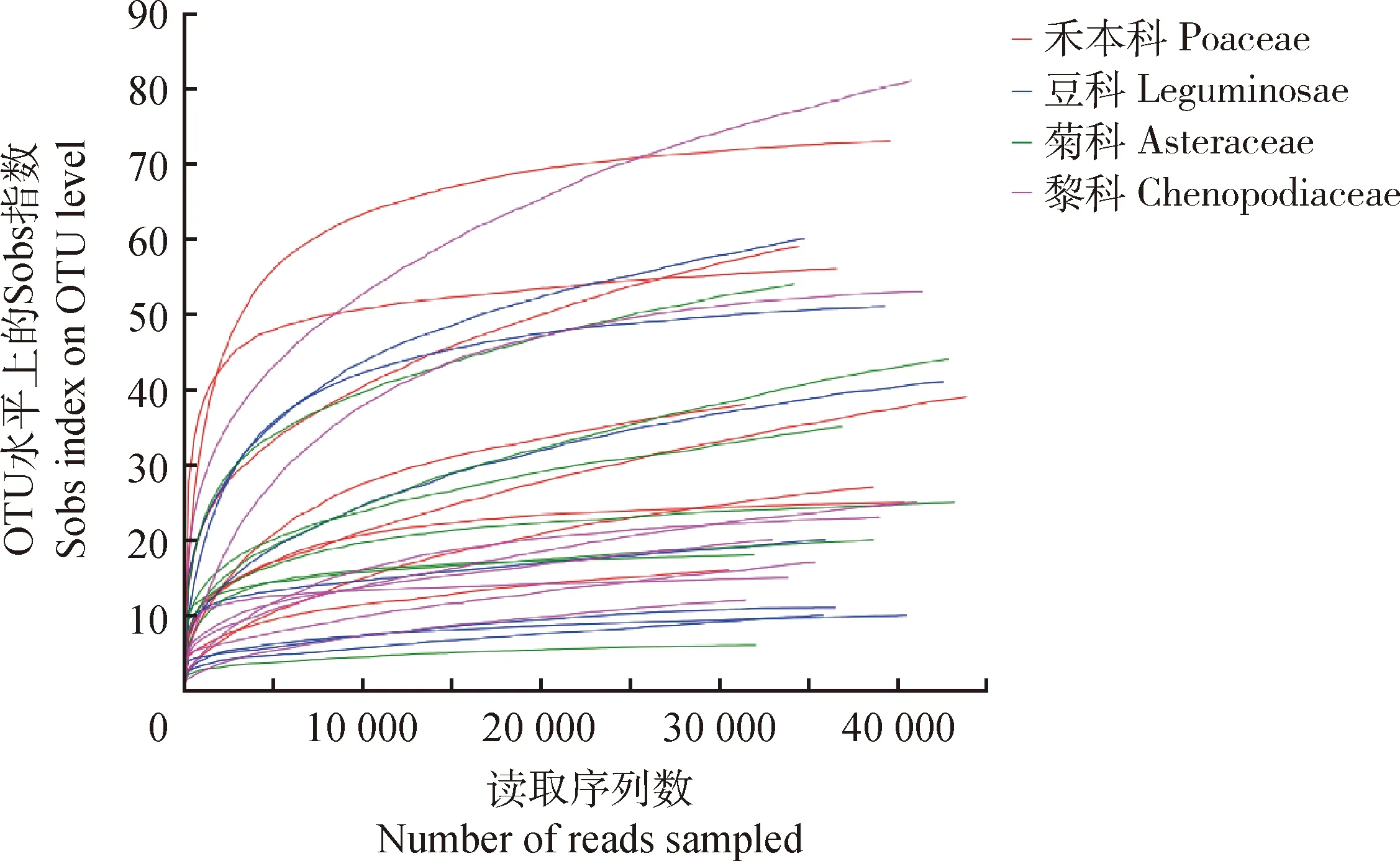
圖1 不同植物類型根際土壤AM真菌群落的稀釋曲線Fig.1 Rarefaction curve of AMF community in rhizosphere soil of different vegetation types
2.2 根際土壤AMF群落結構組成分析
經檢測禾本科、黎科、豆科和菊科植物的平均侵染率分別為:62%,38%,46%,50%。由根際土壤樣品AMF屬水平分析柱狀圖可知(圖2A),共得到5個AMF類群屬(≥不包括相對豐度小于0.5%的類群),球囊霉屬Glomus-f-Glomeraceae相對豐度最高,球囊霉綱未分類屬unclassified-c-Glomeromycetes為優勢屬。球囊霉綱未分類屬在豆科中相對豐度值低于其它植物,僅達到8.6%,而盾巨孢囊霉屬Scutellospora在豆科植物中的相對豐度高于其它處理。類球囊霉屬Paraglomus在禾本科植物中豐度較高。
如圖2B所示,共得到41個AM真菌菌種(不包括相對豐度小于0.5%的類群),AM真菌種水平豐度分別為:禾本科(26)>藜科(22)>菊科(16)>豆科(15),其中球囊霉綱未分類菌種unclassified-c-Glomeromycetes相對豐度最高,在禾本科、藜科和菊科中均達到50%以上,而在豆科中相對豐度較低(8.8%)。球囊霉屬未分類菌種unclassified-g-Glomus-f-Glomeraceae和Glomus-Yamato08-A1-VTX00100分別是禾本科、藜科和菊科植物根際土壤AM真菌的次優勢種和第3優勢種。
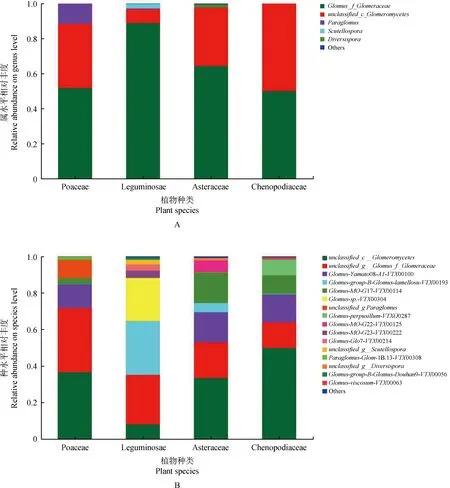
圖2 不同植物根際土壤AM真菌群落結構組成Fig.2 Composition of horizontal community structure of AMF in the rhizosphere soil of different vegetation types注:A,屬水平;B,種水平Note:A,Genus level;B,Species level
豆科植物Glomus-lamellosu-VTX00193的相對豐度高于藜科與菊科。豆科中相對豐度較高的菌種包括球囊霉屬未分類菌種unclassified-g-Glomus-f-Glomeraceae、Glomus-lamellosu-VTX00193和Glomus-sp-VTX00304,其中后2種在另外3科中比例極低。
2.3 根際土壤AMF群落物種組成差異比較
統計不同科植物根際土壤中共有和特有的OTU數量,選擇相似水平97%的OTU樣品表做出維恩圖(圖3)。4科植物共獲得454個OTU,根際土壤AM真菌OTU數量表現為:禾本科(230)>藜科(172)>菊科(125)>豆科(121),各自獨有的OTU數量分別是142個、75個、51個和63個。兩兩共有OTU數量比較,菊科與豆科、菊科與禾本科、藜科與豆科分別共有OTU為40個、48個、40個。其中禾本科與藜科同有OTU數量最多,為總OTU數量的16.3%,禾本科與豆科同有OTU數量最少,為總OTU數量的7.3%,說明禾本科與藜科土壤中AMF群落組成較相似,與豆科土壤中AMF群落組成差異較大。
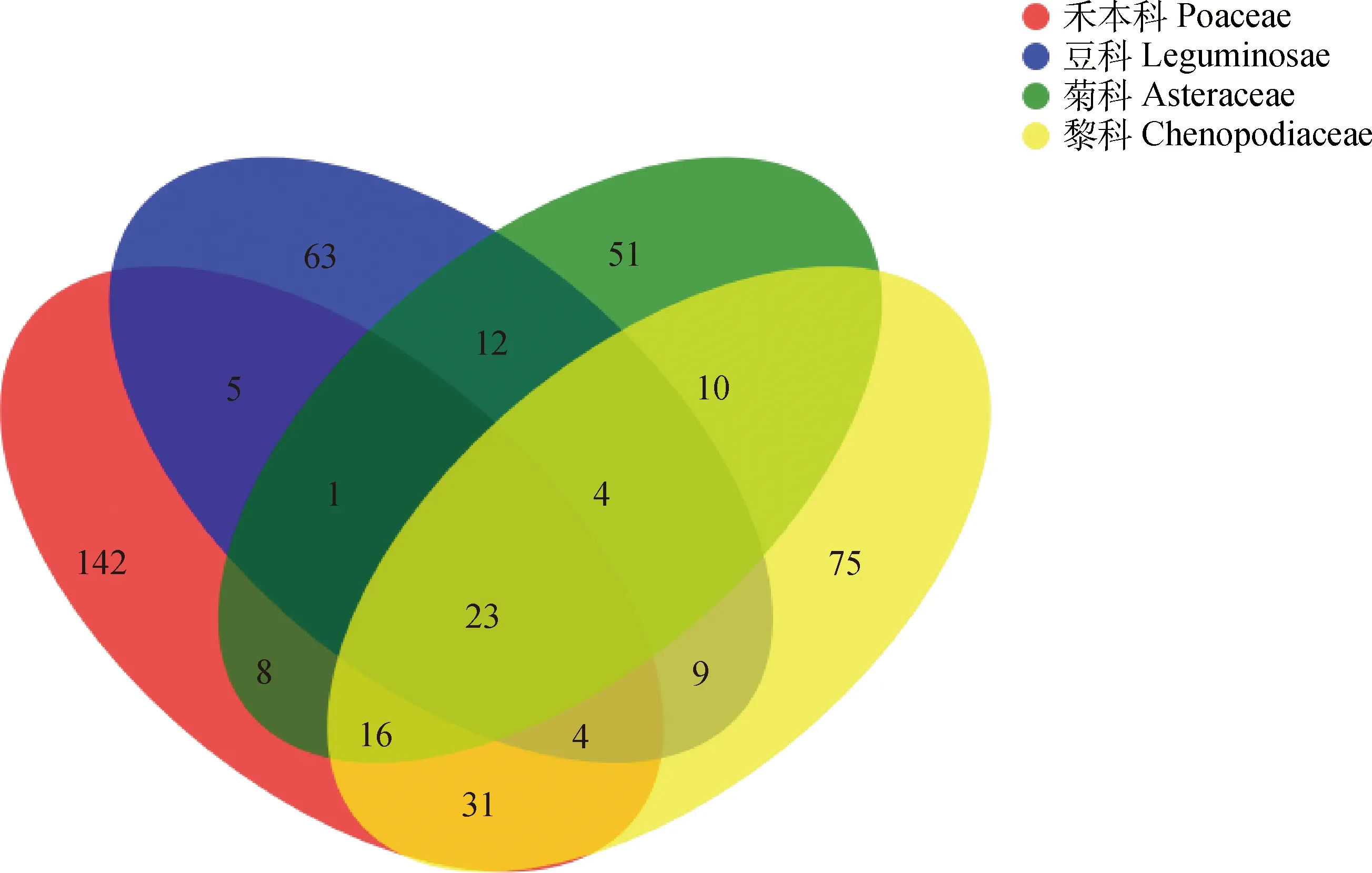
圖3 不同植物根際土壤AM真菌群落的OTU維恩圖Fig.3 OTU Venn diagram of the AMF community in the rhizosphere soil of different vegetation types
2.4 不同科植物AM真菌多樣性及豐富度
由不同植物根際土壤AM真菌多樣性的比較結果可知(表1),禾本植物的平均Sobs指數(53.25)和Chao1指數(57.33)最高,顯著高于豆科和菊科植物(P<0.05),說明禾本科植物根際土壤AM真菌數量最多。菊科的Ace指數低于其它3科植物,顯著低于禾本科植物(P<0.05),根際AM真菌種數最低。豆科的平均Simpson指數(0.49)和藜科的Shannon指數(0.77)最低,表明物種多樣性較低,但均與其它科植物差異不顯著。4科植物的Shannon,Simpson指數沒有顯著差異,禾本科豐富度和多樣性較高,豆科、菊科的豐富度較低。
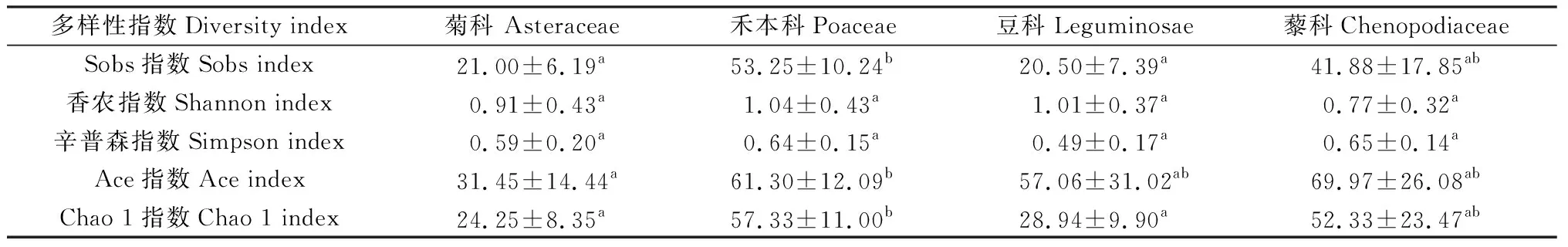
表1 不同植物類型AM真菌多樣性及豐富度Table 1 Diversity and richness of arbuscular mycorrhizal fungi in the rhizosphere soil of different vegetation types
2.5 根際土壤AMF群落NMDS圖分析
根據Bray-Curtis相似性系數,對不同植物根際土壤中的AM真菌群落數據進行多維尺度分析(NMDS),置換多元方差分析(PERMANOVA)表明豆科、菊科、禾本科和藜科植物之間的根際土樣中AM真菌群落種類差異顯著(P<0.05)(圖4),由于禾本科土樣各樣點間的距離極為接近,說明禾本科土樣中AMF群落種類的相似性系數高并且物種最為豐富。菊科、藜科的樣點空間分布零散,其物種數與相似性系數均低于禾本科。
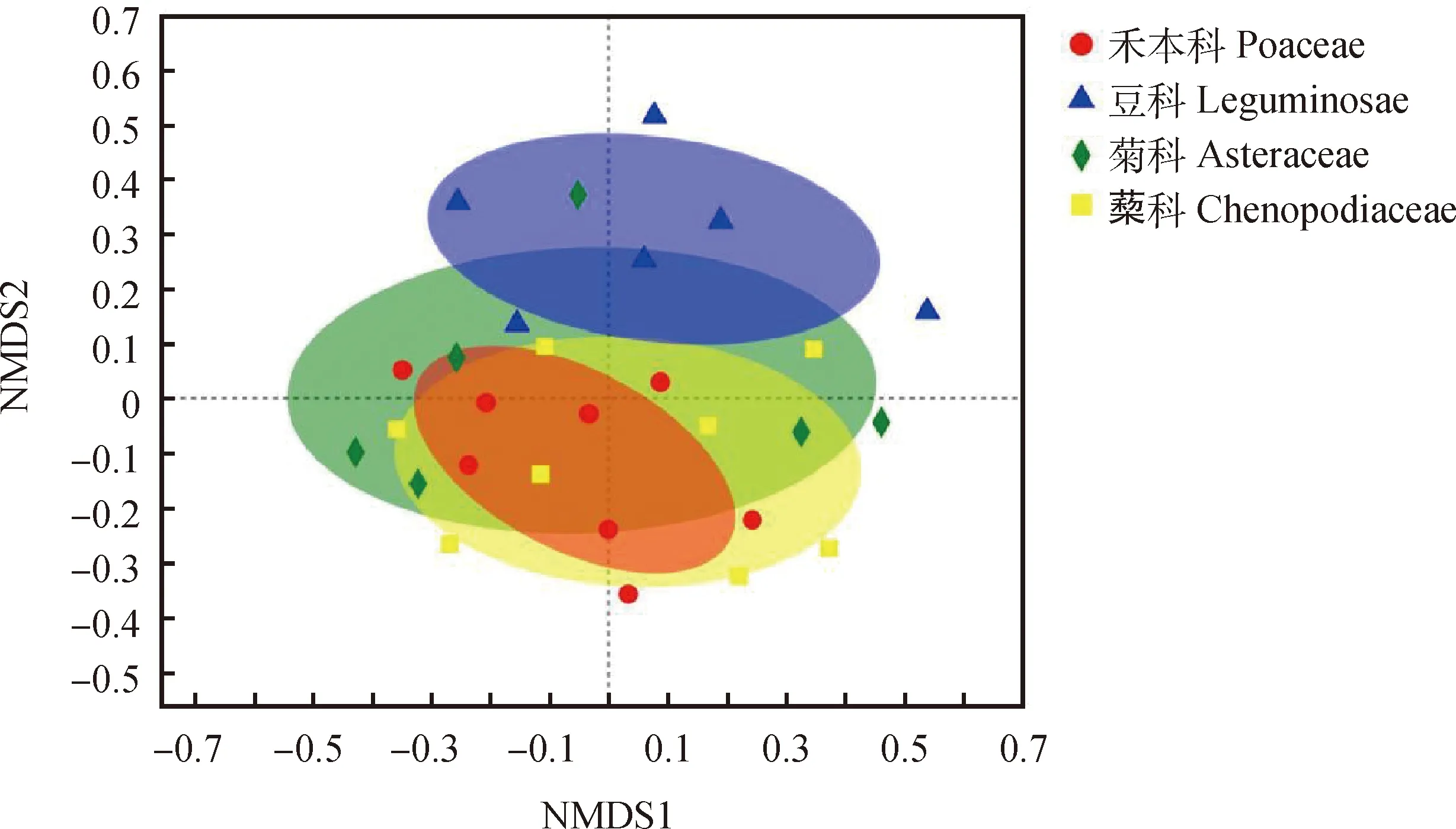
圖4 不同植物的根際土壤AM真菌群落的NMDS圖Fig.4 NMDS map of AMF community in the rhizosphere soil of different vegetation types注:不同顏色橢圓表示不同科植物根際AM真菌群落95%的置信區間,置換多元方差分析顯示Stress=0.178,R=0.151,P=0.017Note:Different color ellipses represent 95% confidence intervals of AM fungal communities in rhizosphere of different plants families. The displacement multivariate analysis of variance shows Stress=0.178,R=0.151,P=0.017
2.6 土壤理化性質分析
由表2可知,煤矸石植物根際土壤偏堿性,同禾本科和菊科植物相比藜科植物顯著降低了根際土壤pH值;禾本科植物根際土壤有機質和速效氮含量最高,顯著高于其它植物(P<0.05),其有機質含量分別是豆科、菊科和藜科植物的5.52,3.72,2.7倍;不同植物根際土壤速效磷和速效鉀含量差異顯著(P<0.05),藜科植物二者含量最高,其速效鉀含量分別是禾本科和菊科的2.13和1.87倍,差異顯著(P<0.05),豆科速效磷含量顯著低于禾本科和藜科植物(P<0.05)。
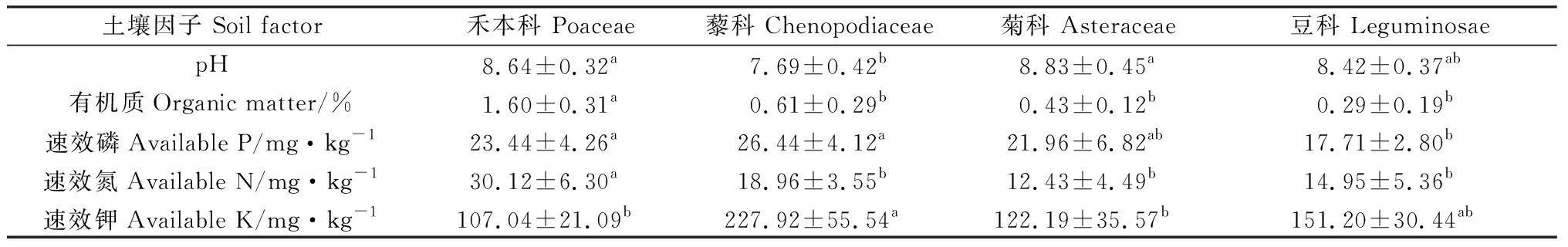
表2 不同植物類型土壤理化性質分析Table 2 Soil physical and chemical properties of different vegetation types
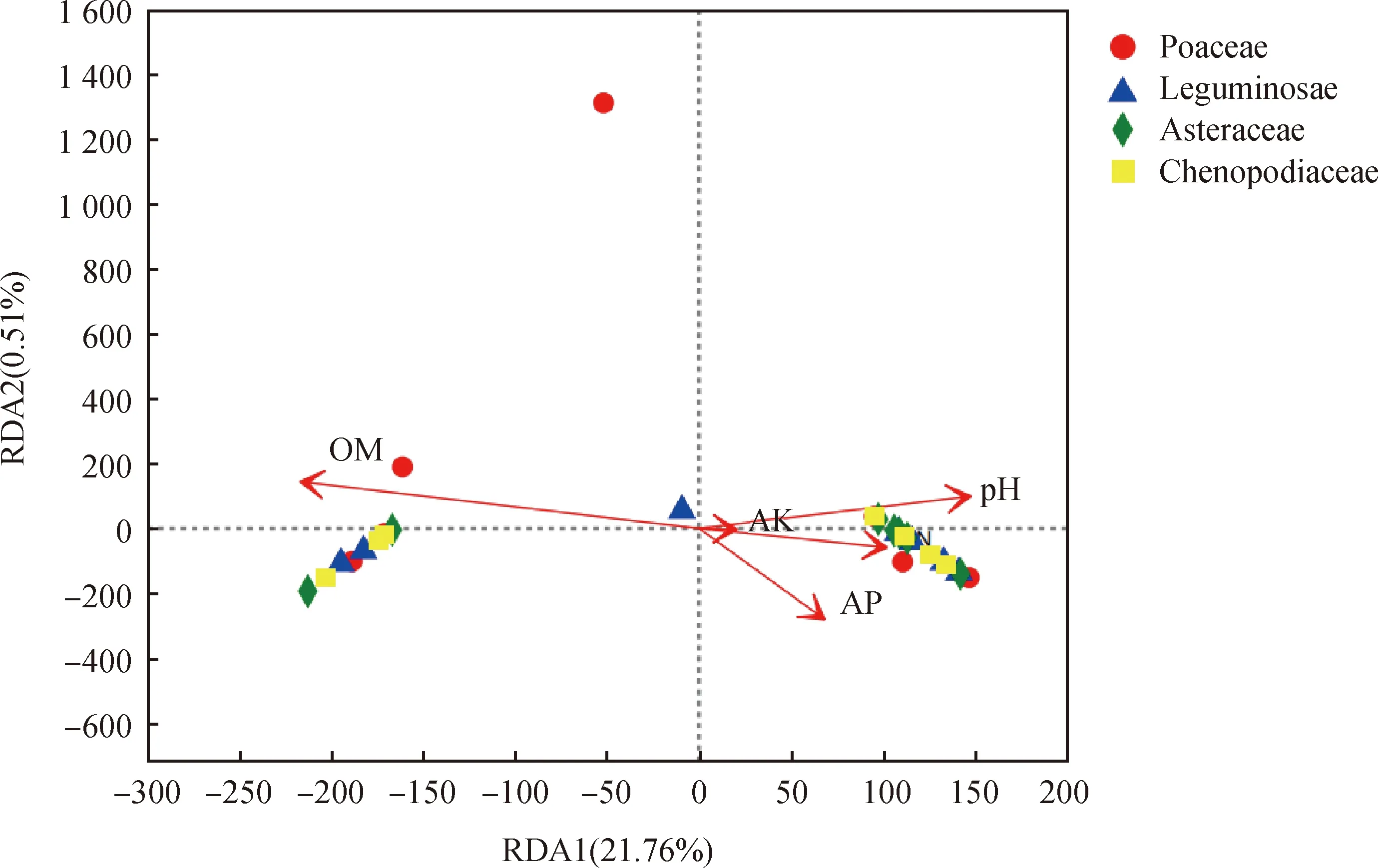
圖5 不同植物AM真菌群落與土壤理化性質的RDA圖分析Fig.5 RDA analysis of AM fungal communities of different plants and soil physical and chemical properties
2.7 RDA圖分析
種水平RDA圖分析結果表明,第一軸(RDA1)對AMF群落分布的方差的解釋率為21.76%,植物根際土壤AMF群落與AN,pH值,AK相關密切,但差異不顯著。第二軸(RDA2)對AMF群落分布的方差解釋率為0.51%,主要影響因子為OM。兩軸共同解釋率為22.27%。
通過構建土壤理化性質與不同科植物AM真菌屬水平的Heatmap圖,發現類球囊霉屬(Paraglomus)與土壤有機質含量極顯著正相關(P<0.001),與土壤速效氮含量顯著正相關(P<0.01),含量較高的未分類屬真菌與土壤有機質含量顯著正相關(P<0.01)(圖6)。
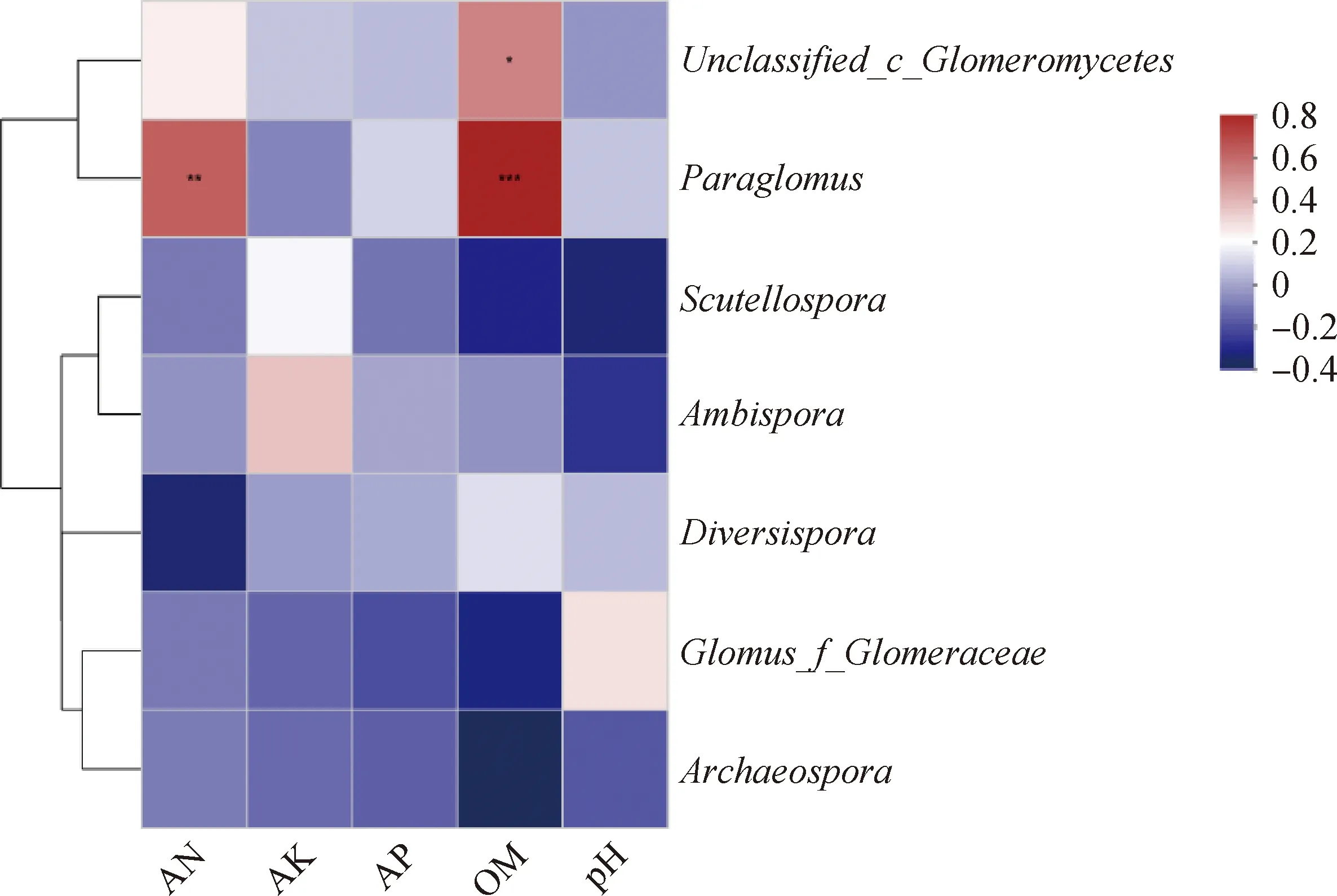
圖6 土壤理化性質與AM真菌優勢屬相關性分析Fig.6 Correlation analysis between soil physical and chemical properties and dominant genus of AM fungi注:*表示P<0.05,**表示P<0.01,***表示P<0.001,紅色代表正相關,藍色代表負相關Note:* means significant differences at the 0.05 level,** means significant differences at the 0.01 level,and ***means significant differences at the 0.001 level. Red is a positive correlation. Blue is a negative correlation
2.8 AM真菌群落種間共生網絡關系圖
通過構建種間共生網絡關系圖,比較不同植物根際AM真菌菌群間的網絡相關性,反映AM真菌菌群的共存格局,探究煤矸石山恢復初期常見的植物類群與土壤菌根真菌群落的共生關系。不同節點代表不同物種,不同顏色代表該物種的種屬,紅色和綠色連線分別表示正負相關,粗細代表相關性大小,節點連線的多少代表該菌種與其它菌種聯系的密切程度。
如圖7所示,煤矸石山禾本科和黎科植物根際土壤菌根真菌種類、共生網絡的邊數量均高于豆科和菊科植物,二者AM真菌網絡規模大于菊科和豆科植物,種間互作關系復雜。4科植物均以球囊霉屬菌種占優勢,禾本科和黎科根際AM真菌核心物種間正相關關系比例更高,禾本科植物菌根真菌菌種正相關關系130個,負相關21,以Unclassified-c-Glomeromycetes,Glomus-M0-G4-VTX00166,Glomus-perpusillum-VTX00 287,Glomus-group-B-Glomus-lamellosn-DVTX00193等27個物種為核心物種;黎科正相關關系152個,負相關32個,以Unclassified-c-Glomeromycetes,Glomus-Wirsel-OTU16-VTX 00156,Glomus-perpusillum-VTX00287等29個菌種為核心物種。豆科和菊科植物正相關分別是26,30個,負相關8,20個,核心物種分別為15和13個。
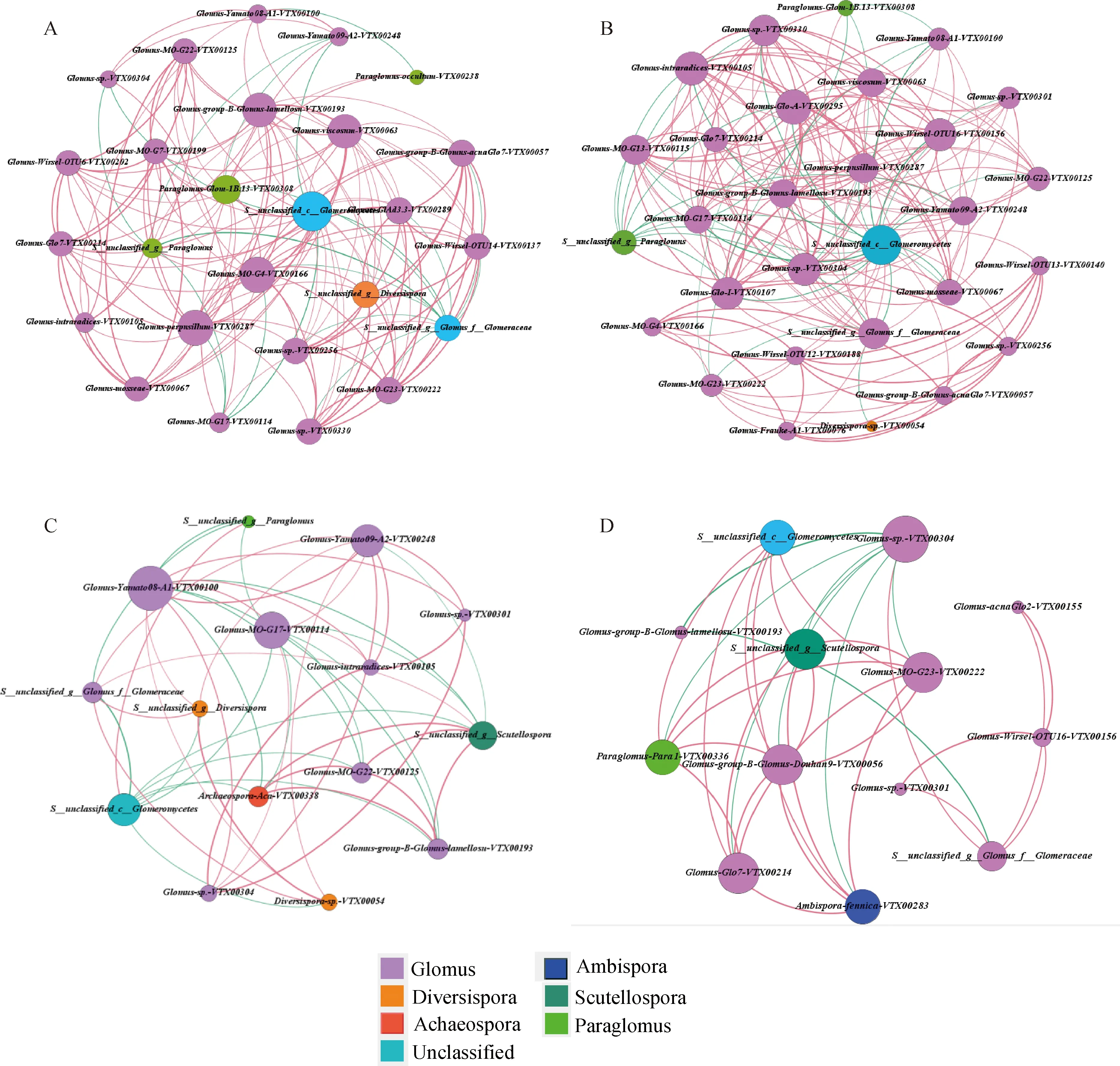
圖7 AM真菌群落相關性網絡分析Fig.7 Network analysis of the AM fungi community注:A,禾本科植物;B,黎科植物;C,豆科植物;D,菊科植物;邊代表兩節點間相關性顯著(P<0.05)Note:A,Gramineae;B,Chenopodiaceae;C,leguminous;D,Asteraceae;The correlation between the two nodes represented by the edge was significant at the 0.05 level
3 討論
真菌多樣性與地上植被多樣性間有很強的耦合性,除植被調查外,土壤真菌多樣性是衡量礦區土壤恢復進程的重要生物指標[24]。本試驗以煤矸石山植物根際土壤菌根真菌為研究對象,比較了不同科植物根際AM真菌群落種類和結構的差異,探究根際土壤AM真菌群落組成與不同植物類型的關系。盡管驅動AM真菌物種功能多樣性的機制尚不清楚,有研究發現與非生物因素相比,植物群落對地下菌根真菌多樣性具有重要的影響力[25],喬木和草本植物物種數量影響著AM真菌的豐度和多樣性[26]。本試驗結果表明不同植物根際土壤AM真菌菌群間差異較大,吳佳偉等[27]也得到了與本研究一致的結果。植物對與其共生的伙伴有一定的選擇作用,有研究發現與豆科和禾本科植物共生的AM真菌菌群不同[28]。AM真菌菌群的功能貢獻差異最有可能與植物特定的養分需求有關[29]。煤礦區寄主植物種類可能通過土壤性狀的變化對AM真菌群落產生強烈的影響[30]。
煤矸石山復墾地土壤OTU數量受到植物優勢類群的影響,調查發現禾本科和藜科OTU數量較多,豆科和菊科的OTU數較少。禾本科的Shannon指數和Chao1指數最高,多樣性和豐富度最高。張海波等發現AM真菌物種豐富度、Shannon多樣性指數受到土壤類型與植物種類交互作用的顯著影響[31];在矸石山恢復初期,禾本科、菊科和藜科植物是先鋒植物,而豆科植物隨著表層基質理化性質的改良出現較晚[9],這可能是導致豆科植物根際AM真菌群落結構與另外3科植物有較大差異的原因。隨著時間的推移,植物群落對AM真菌多樣性的影響,可能是由于在采礦后生態系統發展過程中,植物群落演替引起的植物特定養分需求和寄主特異性的變化引起的[9]。不同人工恢復植被根際AM真菌群落組成不同,增加植物種類可以增加根系和土壤中AM真菌群落的多樣性[32]。
本研究發現球囊霉屬真菌在煤矸石山不同植物根際土壤AM真菌菌群中占據優勢地位,這可能是由于球囊霉屬真菌養分吸收能力強、適應性強、繁殖快、且與草本植物之間能形成較強的共生關系,使其在植物根系和土壤中占優勢[33-35]。其次是球囊霉綱未分類屬豐度較高,該屬物種在其它生境中較少檢測到,疑似為礦區特有菌根真菌類群。類球囊霉屬常出現在煤矸石山恢復后期,而多樣囊霉屬(Diversispora)和盾孢囊霉屬(Scutellospora)出現在恢復初期[26]。本試驗中禾本科植物的類球囊霉屬豐度高于其它科植物,在前期植物調查中發現,禾本科植物在煤矸石山恢復的初期和中期均是矸石山優勢物種,說明禾本科植物能夠與矸石山不同恢復階段的AM真菌形成良好的共生關系,這也可能是其在煤矸石山恢復過程前期處于優勢地位的重要原因。多樣囊霉屬在菊科植物根際土壤中有更高的豐度,有研究發現該屬菌種能夠提高植物在低磷土壤中的適應能力,該屬真菌可能提高了菊科植物在煤矸石山恢復早期低磷土壤中的生存能力[11]。Allen等研究發現原囊霉屬(Archaeospora),多樣囊霉屬和類球囊霉屬隨著矸石山恢復時間的增加,數量變化較大,可能是由于這些物種在功能上與生態過程更相關,而對與其共生的植物資源的需求較低[35]。
禾本科植物根際土壤有機質和速效氮含量高于其他植物,藜科植物根際土壤pH值最低,而土壤速效鉀含量顯著高于其他植物。禾本科為須根系植物,根系發達,易于被AM真菌侵染,使其有較強的養分吸收能力,根際區域有較高的養分含量。藜科植物為非菌根植物,在煤矸石山采集的藜科植物根系中均有菌根侵染現象,并存在較高的豐度,說明在惡劣的矸石環境中,AM真菌與藜科植物形成了良好的共生關系,通過合作,雙方均獲得較好的收益。AM真菌能夠增加生長在礦區等重金屬污染脅迫土壤中植物的根系活力、生物量、根際土壤微生物菌群等,降低對植物的毒害作用[34]。研究發現不同植物根際AM真菌群落網絡關系差異顯著,且關系網中核心物種組成不同,但均以球囊霉屬為主。禾本科和黎科植物網絡連接密集程度更高且核心物種組成更豐富,黎科作為非菌根植物,可能通過形成密集的AM真菌網絡結構,促進了其在矸石山恢復早期的生長。有研究發現,添加AM真菌改善退化土壤微生物群落關系網,尤其是真菌的網絡結構,進而改良了土壤結構[36-37]。在煤矸石山土壤修復過程中,應充分考慮土壤類型及養分含量等因素,選擇適宜的修復植物,發揮AM真菌的生態功能。
4 結論
4科植物共分離到球囊霉綱AM真菌5目7科7屬 41種,禾本科和藜科植物的OTU數量較多。與禾本科植物共生的AM真菌群落多樣性最高,具有較高的α多樣性,且禾本科植物之間根際土壤AM真菌群落種類的相似性系數較高。藜科根際土壤AM真菌菌群組成較為單一,豆科植物根際OTU數量最低。不同科植物之間根際AM真菌群落差異顯著。不同植物根際土壤AM真菌菌種生態網絡關系差異顯著,禾本科和黎科植物網絡連接密集程度較高且核心物種組成更豐富。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
