基于LC-MS/MS分析馬纓杜鵑花代謝物的變化
2022-08-11 13:53:28武紹龍張習敏
廣西植物 2022年7期
武紹龍, 唐 明, 張習敏, 唐 婧*
( 1. 貴州師范大學 生命科學學院, 貴陽 550025; 2. 貴州師范大學, 貴州省植物生理與發育調控重點實驗室, 貴陽 550025; 3. 貴州師范大學, 國家林業和草原局西南喀斯特山地生物多樣性保護重點實驗室, 貴陽 550025 )
馬纓杜鵑()屬于常綠灌木或小喬木,因其花色鮮艷,形狀酷似馬頭飾的瓔珞而得名,主要分布于西南地區,包括四川西南部、云南全省和貴州西北部等(張長芹等,2015),其中貴州西北部以普底和金坡景區分布最為廣泛,為主要的建群種和優勢種(李葦潔和陳訓,2005)。每年3月中旬前完成初花期(花瓣開始開張到10%花序開放);3月下旬到4月初進入為盛開期(10%花序開放到10%花序出現凋謝);4月中旬進入凋謝期(10%花序出現凋謝到70%花序出現凋謝),馬纓杜鵑整個花期為25~30 d。研究馬纓杜鵑花開花至凋謝進程中的代謝組分變化,對延長和調節馬纓杜鵑花期,充分發揮馬纓杜鵑的觀賞價值和經濟價值,對貴州省地方經濟以及鄉村振興具有極其重要的研究意義。目前,關于馬纓杜鵑花主要包括花特征和花粉性狀的研究,以百里杜鵑保護區馬纓杜鵑為實驗材料研究花特征分化和資源分布情況(胡德美 等,2021);用顯微鏡觀察馬纓杜鵑花粉形態大小、外壁紋飾、孔溝形態等性狀(毛加寧,2000)。同時,在馬纓杜鵑花花期調控的研究則主要集中于營養元素、植物激素和溫度等處理方法,包括用不同濃度的N、P、K、Ca等營養元素處理馬纓杜鵑觀察對花芽發育和花期的影響(洪鯤等,2010);在馬纓杜鵑花芽發育初期和開花前期噴施外源激素對始花期、盛開期、末花期和開花數的影響(宋慶發等,2010);通過不同濃度的生長調節物質及礦物質營養處理馬纓杜鵑的花蕾,探索馬纓杜鵑花花期調控方法(岳保超,2015)。目前,僅有少量關于馬纓杜鵑莖和根代謝物的研究報道,利用各種色譜技術從馬纓杜鵑的莖分離鑒定得到10種化合物(徐金金等,2012),然而馬纓杜鵑花的發育進程中代謝物和代謝物途徑研究尚未見相關報道。
近年來,植物代謝組學研究已經被廣泛地應用于分析果樹生長、發育、果實成熟過程中的代謝物變化(Aaron et al., 2008;Zhang et al., 2011;Li et al., 2020),目前也開始應用于研究植物花形成、發育的代謝機制研究(賈巖等,2017;程少禹 等,2020);薛水玉等(2012)采用氣相色譜-質譜法技術分析款冬花不同發育階段代謝組,發現款冬花蕾發育過程中部分特征代謝物呈現顯著變化;He等(2020)利用轉錄組和代謝組研究鐵皮石斛花的發育機制,發現次生代謝和碳水化合物的變化與花的發育有關;徐紅霞等(2020)采用GC-MS技術檢測枇杷花發育進程中的代謝物質,揭示影響花發育進程的生理與代謝機制。代謝組學對植物花的發育代謝產物及代謝通路研究提供了方法。
本研究以貴州省畢節市大方縣百里杜鵑風景區為研究區域,以馬纓杜鵑花為研究對象。選取花苞期、開裂期、傳粉期、盛開期、衰老期和凋謝期的花為材料,采用非靶向代謝組學(LC-MS/MS)分析方法,研究馬纓杜鵑花開花至凋謝進程中6個階段的代謝產物及代謝通路,擬探討以下問題:(1)馬纓杜鵑花開花至凋謝進程中不同階段代謝物如何變化;(2)分析馬纓杜鵑花差異顯著的代謝物和相關的代謝通路;(3)篩選出與馬纓杜鵑花發育相關的重要代謝物。本研究結果為馬纓杜鵑花花期調控提供理論基礎和科學依據。
1 材料與方法
1.1 實驗材料
實驗材料為貴州省畢節市大方縣百里杜鵑風景區馬纓杜鵑。
2020年選取長勢基本一致馬纓杜鵑,分別在花苞期(3月20日)、開裂期(3月24日)、傳粉期(3月30日)、盛開期(4月3日)、衰老期(4月10日)、凋謝期(4月15日),從6株樹上采集適量馬纓杜鵑花樣本,經液氮冷凍后運回實驗室進行代謝組學檢測,每個時期取6個生物學重復樣本。
1.2 馬纓杜鵑花代謝物提取
基于液質聯用(LC-MS/MS)技術(Warwick et al., 2011;Want et al., 2010)進行非靶向代謝組學研究。取100 mg液氮研磨的馬纓杜鵑花樣本,置于EP管中,加入500 μL含0.1%甲酸的80%甲醇水溶液,渦旋震蕩,冰浴靜置5 min,15 000 r·min、4 ℃離心10 min,取100 μL的上清加超純去離子水稀釋至甲醇含量為53%,并置于離心管中15 000、4 ℃離心10 min,收集上清液,進樣LC-MS/MS進行檢測分析。
從每個實驗樣本中取等體積樣本混勻作為(quality control)QC樣本。空白樣本為含0.1%甲酸的53%甲醇水溶液代替實驗樣本,前處理過程與實驗樣本相同。
1.3 非靶向代謝組學(LC-MS/MS)檢測
采用Thermo Hyperil Gold column色譜柱(C18),柱溫40 ℃,流速0.2 mL·min。正離子模式:流動相A為0.1%甲酸,流動相B為甲醇;負離子模式:流動相A為5mmol醋酸銨(pH 9.0),流動相B為甲醇;梯度洗脫(0 ~ 1.5 min,98%A,2%B;1.5 ~ 12 min,100%B;12 ~ 14 min,100%B;14 ~ 14.1 min,98%A,2%B;14.1 ~ 17.0 min,98%A,2%B)。進樣量為10 μL。利用Thermo Vanquish UHPLC液相色譜Thermo QE系列質譜分別進行正、負離子模式采集。掃描范圍選擇70 ~ 1 050;ESI源的設置如下:Spray voltage為3.2 kV;Sheath gas flow rate為35 arb;Aux gas flow rate為10 arb;Capillary temp為320 ℃。Polarity:positive;negative;MS/MS二級掃描為data-dependent scans。每10個分析樣本進行一次QC樣本,評價整個實驗過程中儀器狀態的穩定性和檢測數據結果的準確性。
1.4 數據分析
將采集到的原始數據導入CompoundDiscoverer 3.1(CD)軟件中,通過保留時間、質荷比對數據進行過濾,然后根據保留時間偏差0.2 min和質量偏差5 μg·mL進行峰對齊,隨后根據質量偏差5 μg·mL、信號強度偏差30%(Dai et al., 2017)等信息進行峰提取,同時對峰面積進行定量,再整合目標離子,然后通過分子離子峰和碎片離子進行分子式的預測并與mzCloud、mzVault和MassList數據庫進行比對,用blank樣本去除背景離子,并對定量結果進行歸一化,最后得到數據的鑒定和定量結果。
使用MetaX(Wen et al., 2017)軟件對數據進行對數轉換處理,將所有實驗樣本和QC樣本導入SIMCA14.1軟件進行多元統計分析(阿基業,2010),采用主成分分析(PCA)觀察各樣本的整個過程的穩定性,然后進一步使用有監督的正交偏最小二乘判別分析(OPLS-DA)來區分各組間代謝物的差異性。根據多變量OPLS-DA模型分析中變量權重(>1)、結合檢驗的值(<0.05)和單變量分析差異倍數(fold-change,>2或<0.5)篩選差異代謝物。維恩圖由TB tools 0.6軟件完成,代謝通路富集分析基于KEGG數據庫。
2 結果與分析
2.1 馬纓杜鵑花開花至凋謝過程的形態變化
將馬纓杜鵑花開花至凋謝過程分為6個時期,如圖1所示。花苞期(A):花未開,表現為花苞狀態,內部花藥還未成熟,雄蕊未開裂;開裂期(B):花苞已經初步打開,雄蕊剛開裂,雌蕊柱頭上未有花粉;傳粉期(C):花苞較為開發,雄蕊完全裂開,雌蕊柱頭上已經結合覆蓋花粉;盛開期(D):花朵呈現完全盛開,顏色艷麗,雄蕊已經完全開裂,且花粉已經全部裂出,雌蕊柱頭上花粉已經不多,傳粉受精已完成;衰老期(E):花朵開始凋謝,出現萎蔫狀態;凋謝期(F):花朵完全枯萎、凋謝和萎蔫。

A. 花苞期; B. 開裂期; C. 傳粉期; D. 盛開期; E. 衰老期; F. 凋謝期。A. Bud stage; B. Dehiscence stage; C. Pollination stage; D. Blooming stage; E. Senescence stage; F. Withering stage.圖 1 馬纓杜鵑花開花至凋謝過程的形態變化Fig. 1 Morphological changes of Rhododendron delavayi from flowering to withering
2.2 馬纓杜鵑花樣本主成分分析
為分析馬纓杜鵑花各個時期間代謝組的相互關系,采用主成分分析(PCA)通過建立數學模型進行正交變換簡化降維分析樣本間代謝譜的內部結構關系。馬纓杜鵑花主成分分析(圖2)表明,不同時期代謝物各組間存在明顯的分離趨勢,每個樣本的數據處理結果可信,每個樣本之間存在明顯差異,且組內生物學重復重疊效果好,說明在實驗過程中數據組的重現性好,可用于后續差異代謝物分析。花苞期和開裂期、傳粉期、盛開期代謝物的變化相對較大,衰老期與凋謝期的代謝物相對最大。
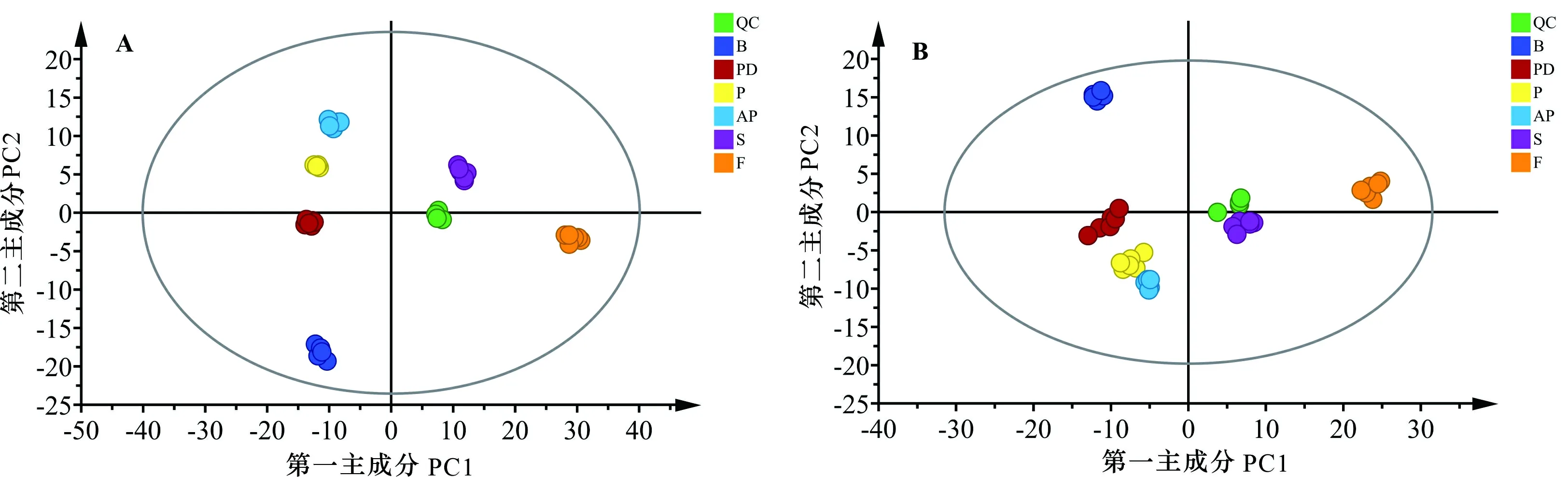
A. 正離子模式; B. 負離子模式。 QC. 質控樣本; B. 花苞期; PD. 開裂期; P. 傳粉期; AP. 盛開期; S. 衰老期; F. 凋謝期。下同。A. Positive ion mode; B. Negative ion mode. QC. Quality control sample; B. Bud stage; PD. Dehiscence stage; P. Pollination stage; AP. Blooming stage; S. Senescence stage; F. Withering stage. The same below.圖 2 馬纓杜鵑花開花至凋謝進程中代謝物PCA模型得分圖Fig. 2 PCA model scores of metabolites from flowering to withering of Rhododendron delavayi
為篩選出馬纓杜鵑開花至凋謝進程中的差異代謝物,采用正交偏最小二乘判別分析(OPLS-DA)區分6個時期代謝物的總體差異,結果如圖3所示。馬纓杜鵑開花至凋謝進程中,花苞期、開裂期、傳粉期、盛開期、衰老期、凋謝期6個時期的代謝物明顯分離,表明各組樣本代謝物存在顯著性的差異變化。該模型質量參數和均大于0.9,說明當前OPLS-DA分析模型穩定可靠,具有良好的預測能力,有利于尋找潛在的馬纓杜鵑開花至凋謝進程中的差異代謝物。
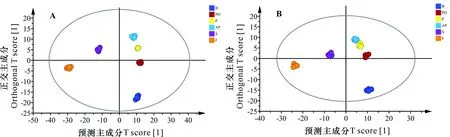
圖 3 馬纓杜鵑花開花至凋謝進程中代謝物OPLS-DA得分圖Fig. 3 OPLS-DA scores of metabolites from flowering to withering of Rhododendron delavayi
2.3 馬纓杜鵑花代謝物分析
研究馬纓杜鵑花開花至凋謝進程中代謝物的變化,采用LC-MS/MS檢測馬纓杜鵑花6個時期的代謝物多峰圖,花苞期代謝物色譜圖見圖4。共檢測到973種代謝物,正離子模式587種,負離子模式386種,其中黃酮類114種;有機酸114種;酚酸類112種;氨基酸及其衍生物75種;脂類61種;生物堿57種;萜類45種;核苷酸及其衍生物43種;糖類及其衍生物36種;苯丙素類33種;醇和多元醇33種;鞣質11種;甾體3種;其他類236種。
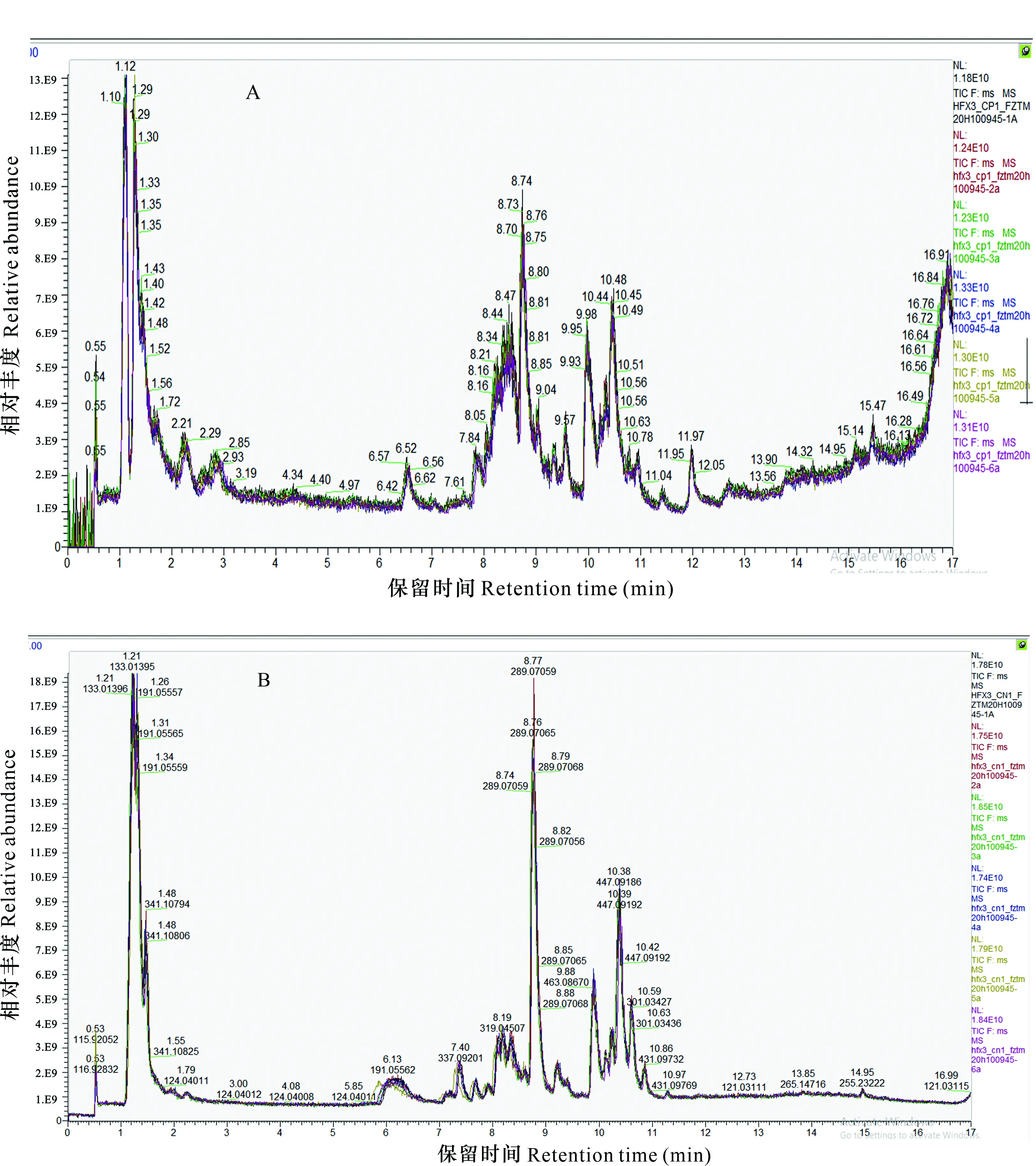
圖 4 LC-MS/MS代謝物檢測多峰圖Fig. 4 LC-MS/MS metabolite detection multimodal graph
2.4 差異代謝物分析
根據OPLS-DA模型的變量重要性投影值篩選馬纓杜鵑花6個時期的差異代謝物(>1、<0.05和>2 或<0.5)。從花苞期、開裂期、傳粉期、盛開期、衰老期到凋謝期進行相鄰兩個時期的代謝組比較,結果發現:馬纓杜鵑花開花到凋謝進程中共有591種差異代謝物。其中,花苞期與開裂期有170種差異代謝物(正離子模式87,負離子模式83),48種上調,122種下調;開裂期與傳粉期有144種差異代謝物(正離子模式89,負離子模式55),68種上調,76種下調;傳粉期與盛開期有111種差異代謝物(正離子模式67,負離子模式44),75種上調,36種下調;盛開期與衰老期有284種差異代謝物(正離子模式184,負離子模式100),177種上調,107種下調;衰老期與凋謝期有314種差異代謝物(正離子模式180,負離子模式134),260種上調,54種下調。在馬纓杜鵑花期進入衰老期和凋謝期后差異代謝物數量明顯增多于開花過程,且大部分是表達量顯著上升的差異代謝物。而在馬纓杜鵑花苞期至開裂期過程中,差異代謝物中大部分的表達量顯著下調。差異代謝物維恩圖分析表明(圖5),絕大部分(≥46.8%)的差異代謝物都只出現在一個相鄰變化時期,沒有任何差異代謝物出現在整個開發至凋謝過程。
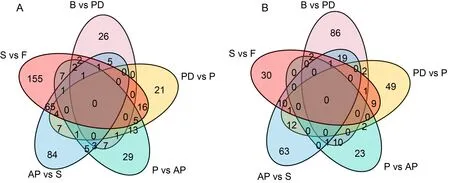
A. 上調代謝物; B. 下調代謝物。A. Up-regulated metabolites; B. Down-regulated metabolites.圖 5 馬纓杜鵑花開花至凋謝進程中差異代謝物維恩圖Fig. 5 Venn diagram of differential metabolites from flowering to withering of Rhododendron delavayi
2.5 代謝通路分析
通過KEGG Pathway富集分析顯著差異代謝物(Kanehisa & Goto, 2000;Lin et al., 2014),如圖6所示。注釋到68條代謝通路,顯著代謝通路(<0.05)要有6條,分別是苯丙素類生物合成,植物激素的生物合成,類黃酮生物合成,精氨酸和脯氨酸代謝,苯丙氨酸、酪氨酸和色氨酸生物合成鳥氨酸、賴氨酸和煙酸生物堿的生物合成。其中,極顯著的代謝通路(<0.01)包括苯丙素類生物合成、植物激素的生物合成和類黃酮生物合成。

圖 6 馬纓杜鵑花開花至凋謝進程中差異代謝物主要代謝通路富集分析Fig. 6 Enrichment analysis of main metabolic pathways of differential metabolites from flowering to withering of Rhododendron delavayi
本研究根據代謝組學分析差異代謝物與代謝通路的關系,挖掘與馬纓杜鵑花開花至凋謝相關的差異代謝物,推測代謝物可能的作用機制。結合代謝通路圖(圖7),共篩選到10種差異代謝物,包括苯丙氨酸(L-phenylalanine)、反式肉桂酸(-cinnamic acid)、 查耳酮 (chalcone)、 柚皮素(naringenin)、對香豆酰基莽草酸(-coumaroyl shikimic acid)、阿魏酸(ferulic acid)、松柏醇(coniferyl alcohol)、芥子酸(sinapic acid)、紫丁香苷(syringin)、槲皮素(quercetin)。在馬纓杜鵑花的6個時期,代謝物含量表達如圖6所示,苯丙素類生物合成相關的差異代謝物包括苯丙氨酸、反式肉桂酸、對香豆酰莽草酸、松柏醇、芥子酸、紫丁香苷在衰老期(S)和凋謝期(F)兩個時期均為上調表達,特別是在凋謝期(F)達到最高值;而黃酮類生物合成相關的代謝物柚皮素、槲皮素和查爾酮隨著花的生長發育含量呈現下調表達。在‘夏黑’葡萄二季果冬芽分化關鍵時期,CCC處理會誘導苯丙素合成和類黃酮合成通路中部分基因的上調或下調表達,通過改變與成花相關的基因表達和次級代謝產物的合成和代謝來實現促進成花(時曉芳等,2021)。轉錄組學分析不同發育階段款冬葉,發現葉片生長發育的過程中苯丙素類生物合成相關基因表達量不斷增多(聶佳慧等,2018)。馬纓杜鵑花開花至代謝進程中苯丙素類代謝物表達量逐漸升高,而黃酮類代謝物表達量逐漸降低,苯丙素類和黃酮類代謝物表達可能與馬纓杜鵑花發育有關。
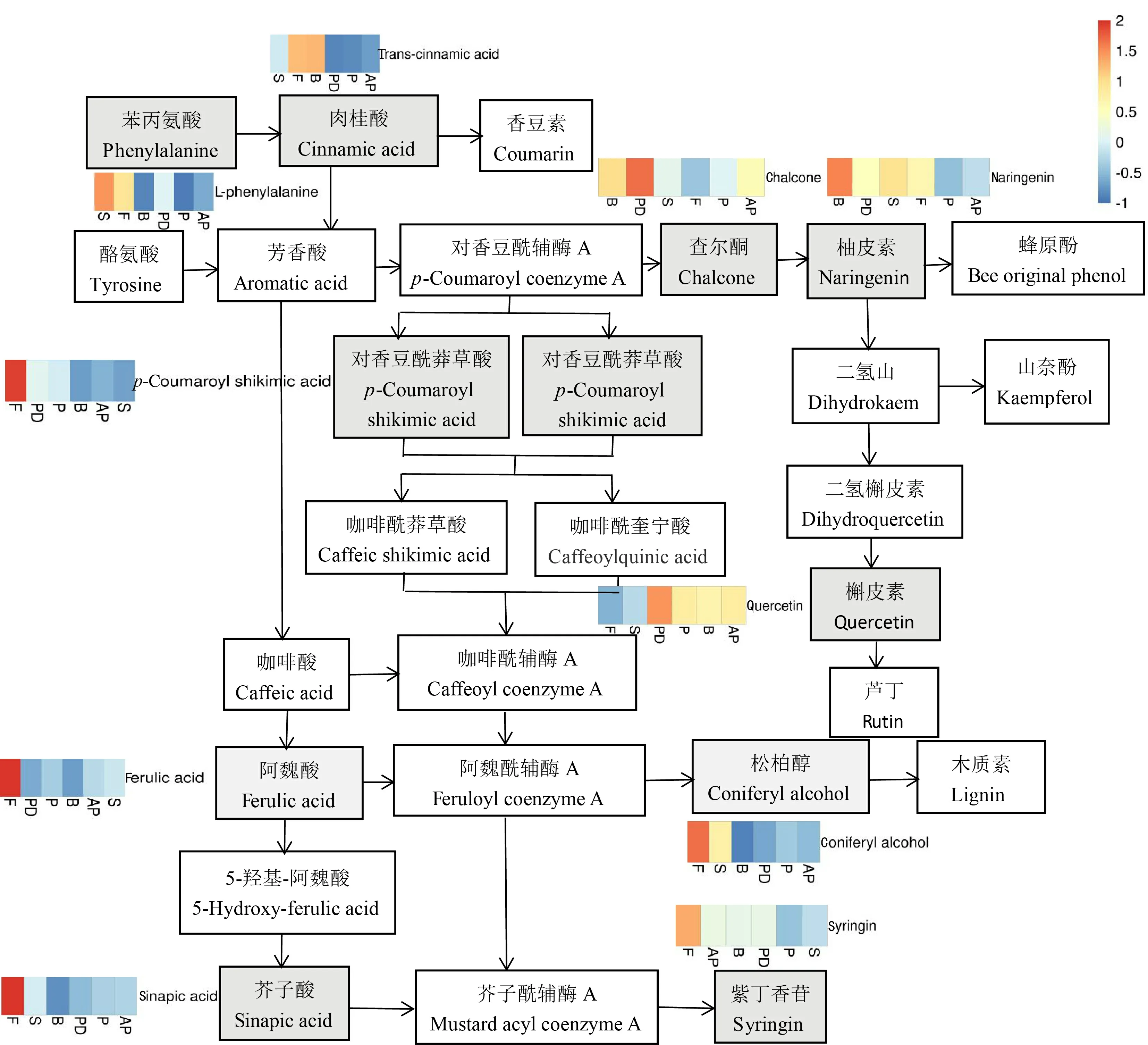
圖 7 苯丙素類、黃酮類等有效成分生物合成通路Fig. 7 Biosynthesis pathway of active components about phenylpropanes and flavones
3 討論與結論
3.1 馬纓杜鵑花差異代謝物
代謝物是生物體表型的基礎,能幫助我們更直觀有效地了解生物學過程及其機理(熊歡歡等,2019;霍冬敖等,2021)。采用GC-MS技術對枇杷花整個發育進程中代謝物質分析,C/N含量比是決定花發育進程的重要因素之一(徐紅霞等,2020)。長期以來馬纓杜鵑花花期調控一直都是研究熱點,馬纓杜鵑花的開花至凋謝進程是十分復雜的生理過程,該過程可能與代謝物種類和含量密切相關。本研究采用非靶向代謝組學方法,研究馬纓杜鵑花開花至凋謝的6個時期(花苞期、開裂期、傳粉期、盛開期、衰老期和凋謝期)代謝物變化。通過代謝組學分析,共篩選獲得591種差異表達代謝物,主要包括黃酮類、有機酸、酚酸類、氨基酸,及其衍生物、脂質和生物堿等,這些差異代謝物在花的生長發育不同時期表達情況不相同,隨著馬纓杜鵑花的生長發育差異代謝物在花苞期至盛開期逐漸減少,進入衰老期和凋謝期后顯著增加;而上調代謝物在整個開花至凋謝進程中逐漸增加,并且在凋謝期達到最大值;下調代謝物則在花苞期至盛開期逐漸減少,衰老期增加,凋謝期減少。本研究表明,馬纓杜鵑花的不同發育階段差異代謝物種類和表達可能與花的開花至凋謝進程密切相關。
3.2 馬纓杜鵑花差異代謝通路
基于KEGG PATHWAY數據庫對差異代謝物富集分析,注釋到68條代謝通路,其中3條極顯著的代謝通路(< 0.01)分別是苯丙素類生物合成、植物激素的生物合成和類黃酮生物合成。其中,苯丙素類生物合成是最顯著的代謝通路,苯丙氨酸為下游次生代謝產物提供原料。本研究基于苯丙素類、黃酮類等有效成分生物合成通路共篩選獲得10種有效成分生物合成有關的差異代謝物。進一步研究發現,苯丙素類生物合成相關的代謝物表達量同馬纓杜鵑花的其他時期相比,在衰老期和凋謝期代謝物表達量逐漸升高,而黃酮類生物合成相關的代謝物隨著花的生長發育含量呈現下調表達,這些代謝物可能對馬纓杜鵑花凋謝有重要調控作用。在馬纓杜鵑花的花苞期苯丙氨酸表達量相對較低,隨著花的生長發育代謝物表達量逐漸升高,代謝通路上相關代謝也發生相應變化,說明它對整個代謝通路的完成具有重要調控作用。
現有研究發現,苯丙素類生物合成是最重要的植物次生代謝途徑之一(Dong & Lin, 2021),植物的苯丙素類生物合成代謝途徑及其分支途徑會產生許多次生代謝產物,如色素、酚酸、黃酮、類黃酮、木質素等(文歡等,2017)。款冬花不同生長發育階段的次生代謝物代謝組成明顯不同,苯丙素類代謝物在發育初期至中后期表達量較高;黃酮類代謝物在發育的各個階段表達量均有波動,但總體變化不大,次生代謝物累積促進花的發育(賈巖等,2017)。采用高通量測序技術對川佛手不同發育時期轉錄組學研究,發現苯丙素生物合成中7個相關基因隨著果實的發育及變色呈現顯著差異表達,提高其表達從而促進果實發育(潘媛等,2020)。苯丙素類生物合成途徑的前端苯丙氨酸轉化為反式肉桂酸,再由肉桂酸羥化酶(C4H)產生對羥基香豆酸,再經4-香豆酰-CoA 連接酶(4CL)催化生成4-香豆酸CoA,進一步向下游特異性合成途徑,轉化為不同的苯丙素類代謝產物,包括香豆素、黃酮類、萜類和木質素等(王玉等,2019;霍冬敖等,2021)。在馬纓杜鵑花開花至凋謝進程中苯丙素類生物合成相關的差異代謝物苯丙氨酸、反式肉桂酸、對香豆酰莽草酸、松柏醇、芥子酸、紫丁香苷代謝物隨著花的生長發育含量呈現上調表達,特別是在凋謝期(F)達到最高值;而黃酮類生物合成相關的代謝物柚皮素、槲皮素和查爾酮在花的生長發育過程中下調表達。對不同發育時期大白菜花的轉錄組研究,發現花香的主要成分是苯丙素,而黃酮類化合物是花、果實和種子的主要顯色物質,此外它還能影響花瓣的伸長,促進花粉的萌發(李改珍等,2017)。在苯丙素類生物合成中L-苯丙氨酸(侯思宇等,2015)、柚皮素(Tu et al., 2016)、阿魏酸(Louie et al., 2010)等作為中間體,它們的表達量高低直接限制到下游代謝途徑的進行,從而影響花的開花至凋謝過程。苯丙素類、黃酮類等代謝物作為重要次生代謝產物,推測其可能為馬纓杜鵑花開花至凋謝進程具有重要的調控作用。更好地了解代謝物的生物合成與馬纓杜鵑花開花至凋謝進程中的關系還需通過進一步的深入分子水平研究。
本研究通過非靶向代謝組學技術對馬纓杜鵑花開花至凋謝進程中不同階段的代謝成分進行分析,基于KEGG數據庫對差異代謝物進行注釋并獲得了相應通路,構建了馬纓杜鵑花中苯丙素類、黃酮類等有效成分生物合成途徑,為進一步研究代謝物的基因調控作用以及全面解析代謝途徑提供代參考,為延長馬纓杜鵑花花期調控機制提供了理論基礎和科學依據。
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
音樂探索(2022年2期)2022-05-30 21:01:37
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
航空世界(2020年10期)2020-01-19 14:36:20
中國外匯(2019年20期)2019-11-25 09:54:58
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
民主與科學(2014年3期)2014-02-28 11:23:03
