2種體型禾花鯉形態性狀對體質量的影響
2022-08-09 08:32:56樊佳佳馬冬梅朱華平蘇換換李婭婭黃樟翰李輝安黃劍華
南方農業學報 2022年5期
樊佳佳,馬冬梅*,朱華平,蘇換換,李婭婭,黃樟翰,李輝安,黃劍華
(1中國水產科學研究院珠江水產研究所/農業農村部熱帶亞熱帶水產資源利用與養殖重點實驗室,廣東廣州 510380;2廣東省水產動物免疫技術重點實驗室,廣東廣州 510380;3上海海洋大學水產與生命學院,上海 201306;4陽江市中睿農業發展有限公司,廣東陽江 529827;5乳源瑤族自治縣一峰農業發展有限公司,廣東韶關 512700)
0 引言
【研究意義】鯉(Cyprinus carpioLinnaeus)作為重要的經濟魚類,廣泛養殖于歐亞大陸(Biermann and Geist,2019)。由于地理隔離和人工選擇等因素影響,鯉魚地方種群較多(Casal,2006;明俊超等,2009),目前經過人工培育的鯉魚品種也很多,如紅鯉、團鯉、草鯉、芙蓉鯉、荷包鯉和錦鯉等。廣東北部山區的稻田禾花鯉養殖歷史悠久,該養殖模式具有“以漁促稻、穩糧增效、質量安全和生態環保”的優點,已成為發展農村經濟的重要途徑(Xie et al.,2011;馬達文等,2016)。經調查研究發現,因未經過人工選育,野生稻田養殖的禾花鯉種群存在較大的遺傳變異,按體型可分為偏短圓型和偏修長型兩類(馬冬梅等,2019)。北部山區乳源瑤族自治縣養殖戶也一直將稻田養殖短圓體型的禾花鯉稱為“石鯉”,稻田養殖長體型的禾花鯉稱為“柴鯉”。為增加稻田養殖禾花鯉的經濟效益,提高禾花鯉的產量,需對其進行選育。在選育禾花鯉新品種過程中,因其生長性狀很多,若均選作為選育指標,工作量會很大,因此通過分析兩種體型禾花鯉形態性狀對體質量的影響,找出最重要的關鍵性狀,將對后續育種工作具有重要意義。【前人研究進展】研究待選育物種的數量性狀遺傳規律是制定科學選育策略的基礎,而形態性狀和體質量是魚類遺傳改良和育種的主要目標性狀(覃寶利等,2020)。對魚體質量的測定往往會受到腸道內殘留物、體表水分等因素影響,導致測量數據有一定的偏差,而形態性狀的測量不受上述因素的影響,更易準確測量。分析魚類形態性狀和體質量關系的研究方法較多,最常見的是通徑分析(董浚鍵等,2018;陳紅林等,2019;張新明和程順峰,2020)和灰色關聯度分析(劉峰等,2017;劉陽等,2019),通過獲得對體質量影響重要的形態性狀,為研究對象生長性狀評估、良種改良或選育提供重要依據。通徑分析是指利用通徑系數分析多個自變量對因變量相互影響大小,分為直接通徑系數(直接影響)和間接通徑系數(間接影響),直接通徑系數為標準化回歸方程中的偏回歸系數,而間接通徑系數是在直接通徑系數基礎上計算獲得。該分析方法因可詳細獲得自變量與因變量、自變量與自變量之間的影響大小,所以結果準確性較高,但該分析方法需要樣本量足夠大,且數據符合一定的分布規律,目前被廣泛應用在魚類,如大口黑鱸(Micropterus salmoides)(何小燕等,2009)、斜帶石斑魚(Epinephe‐lus coioides)(趙旺等,2017)、翹嘴鱖(Siniperca chuatsi)(董浚鍵等,2018)、甌江彩鯉(Cyprinus carpiovar.Color)(陳紅林等,2019)和星康吉鰻(Conger myriaster)(張新明等,2020)等。灰色關聯度分析是一種通過計算性狀間關聯度,以確定性狀間的重要性,因其可以對較少樣本量的數據進行分析,目前在魚類也有很多報道,如鯉魚(蘇勝彥等,2011)、牙鲆(Paralichthys olivaceus)(劉永新等,2014)、小黃魚(Pseudosciaena polyactisBleeker)(劉峰等,2017)和許氏平鮋(Sebastes schlegelii)(劉陽等,2019)等。【本研究切入點】同時選擇通徑分析和灰色關聯度分析方法進行分析,可增加結果可信度,但目前鮮見同時利用這兩種分析方法對稻田養殖不同體型的禾花鯉形態性狀與體質量相關性的研究報道。【擬解決的關鍵問題】按照體型將廣東北部地區傳統養殖的稻田禾花鯉分為短圓體型和長體型,同時利用通徑分析和灰色關聯度分析方法分析長體型和短圓體型禾花鯉的形態特征及其形態性狀與體質量的相關性,為按照體型確定選育禾花鯉測量指標提供參考數據。
1 材料與方法
1.1 試驗材料
供試禾花鯉養殖在廣東省韶關市乳源縣的稻田綜合種養基地。2020年5月25日取體長約5 cm的禾花鯉苗種投放在稻田中進行養殖,投放密度為7500尾/ha,稻田養殖期間僅以稻田中的昆蟲、田螺、水草和稻花等天然餌料為食,不投喂人工配合飼料。同年11月25日隨機取146尾禾花鯉轉運至實驗室,暫養在水泥池中24 h后進行生長性狀測量,按馬冬梅等(2019)的聚類分類法將146尾禾花鯉分為短圓體型(93尾)和長體型(53尾)。
1.2 測量方法
采用60 mg/L魚安定(MS-222)對待測禾花鯉進行淺度麻醉,電子天平稱活體質量(精確至0.01 g)。利用數碼相機對擺有刻度尺的禾花鯉進行拍照,照片采用魚類外部形態測量V 1.0軟件對全長、體長、頭長、體高、尾柄長、尾柄高和尾長進行測量(精確至0.01 cm)。頭高和體寬采用游標卡尺測量(精確至0.01 cm)。
1.3 統計分析
1.3.1 通徑分析 將測量的禾花鯉生長數據進行整理和分析,將體質量作為因變量Y,全長、體長、頭長、體高、尾柄長、尾柄高、尾長、頭高和體寬作為自變量,分別用X1、X2、X3、X4、X5、X6、X7、X8和X9表示,利用Excel 2007統計上述10個生長性狀數據的平均值、標準差等表型參數。采用SPSS 19.0的Analyze-Non‐parametric Tests—1-Sample K-S模塊對數據進行正態性檢驗。利用其Analyze—Correlate—Bivariate模塊對表型數據進行相關分析。利用其Analyze—Re‐gression—Linear模塊對數據進行回歸分析和構建多元回歸方程。參照杜家菊和陳志偉(2010)的方法計算直接通徑系數、間接通徑系數、單性狀決定系數和兩兩性狀間共同決定系數。
1.3.2 灰色關聯度分析 參照于萍和李克(2011)的灰色關聯分析系統,將禾花鯉體質量和9個形態性狀視為一個灰色關聯系統。首先將體質量作為參考序列x0,9個形態性狀作為比較序列x1~x9,利用標準差對數據進行無量綱化處理,然后計算9個形態性狀的灰色關聯系數和關聯度,最后根據9個形態性狀關聯度的大小確定形態性狀對體質量的關聯程度,按照從大到小排出關聯序,關聯序越小其對體質量越重要,計算公式如下:
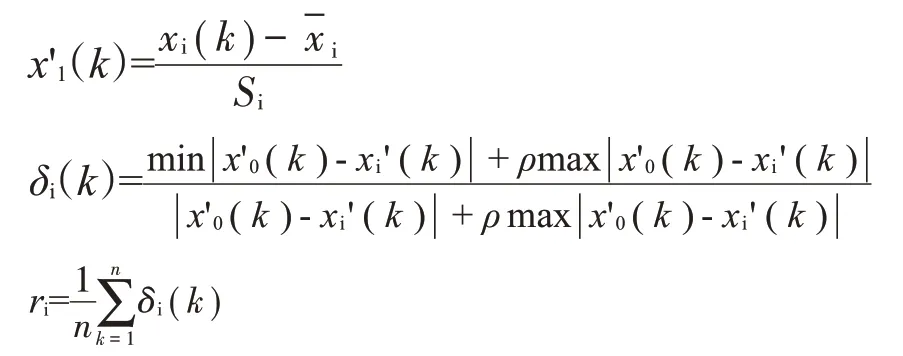
式中,x'(1k)為無量綱處理后的數據,x(ik)為表型性狀原始測量值為表型性狀的平均值,Si為表型性狀的標準差。δ(ik)為形態性狀的灰色關聯系數,ρ為分辨系數,本文設定ρ=0.5,min |x'0(k)-xi'(k)|表示形態性狀絕對差值中的最小值,max |x'0(k)-xi'(k)|表示形態性狀絕對差值中的最大值,ri表示形態性狀xi對體質量xo的關聯度,最后根據關聯度數值編排關聯序,分別用1~9表示。
2 結果與分析
2.1 不同體型的禾花鯉生長性狀數據描述統計
146尾禾花鯉分為短圓體型群體(93尾)和長體型群體(53尾)。對這2個群體的全長、體長、頭長、體高、尾柄長、尾柄高、尾長、頭高、體寬和體質量的平均值、標準差和變異系數進行描述統計,結果如表1所示。短圓體型群體和長體型群體的體質量變異系數分別為44.51%和43.90%,表明禾花鯉體質量的選擇潛力較大。對短圓體型群體和長體型群體的生長性狀進行顯著性檢驗,結果顯示除了尾柄高、體寬和體質量在兩群體間差異未達顯著性水平(P>0.05,下同),全長、體長、頭長、體高、尾柄長、尾長和頭高在兩群體間差異均達極顯著性水平(P<0.01,下同)。對生長性狀進行正態性檢驗,結果顯示所有性狀均符合正態分布(P>0.05),可進行通徑分析。

表1 禾花鯉不同體型群體的生長性狀參數Table 1 Growth trait parameters of rice flower carp populations of different body shapes
2.2 禾花鯉各生長性狀間的相關分析結果
對短圓體型群體和長體型禾花鯉群體的各生長性狀進行相關分析,結果如表2所示。各群體內各性狀之間的相關系數均為極顯著相關,其中2個群體的全長與體長的相關系數(R)均最大,分別為0.991和0.995,表明這兩個性狀相關性最大。短圓體型群體的尾柄長與體寬的相關系數最小,為0.490;長體型群體的頭長與尾長的相關系數最小,為0.343。從體質量和形態性狀的相關系數來看,短圓體型和長體型群體中,與體質量相關系數最大的分別為體長(0.934)和全長(0.931),而相關系數最小的分別為尾柄長(0.670)和頭長(0.486)。
2.3 禾花鯉各形態性狀對體質量的通徑分析結果
由表3和表4可知,短圓體型群體起主要作用的形態性狀有4個,分別為體長、尾柄高、體高和頭高,長體型群體起主要作用的形態性狀有5個,分別為全長、體寬、體高、頭長和頭高。短圓體型群體對體質量直接通徑系數最大的形態性狀為尾柄高,直接通徑系數為0.537;長體型群體對體質量直接通徑系數最大的形態性狀為全長,直接通徑系數為0.408。從間接通徑系數總和來看,短圓體型對體質量間接作用最大的形態性狀為體長,間接通徑系數總和為0.804;長體型對體質量間接作用最大的形態性狀為頭長,間接通徑系數總和為0.692。

表3 短圓體型禾花鯉形態性狀對體質量的通徑分析Table 3 Path analysis of morphological traits to body mass in round-shape rice flower carps

表4 長體型禾花鯉形態性狀對體質量的通徑分析Table 4 Path analysis of morphological traits to body mass in long-shape rice flower carps
2.4 禾花鯉各形態性狀對體質量的決定系數
由表5可知,短圓體型群體的尾柄高對體質量的直接決定系數最大,為0.289,尾柄高和體高對群體的共同決定系數最大為0.127;長體型群體的全長對體質量的直接決定系數最大,為0.166,全長和體寬對群體的共同決定系數最大為0.224。由表6可知,短圓體型和長體型群體中對體質量起主要作用的形態性狀組合與體質量相關系數(R)分別為0.975和0.976,復相關系數(R2)分別為0.951和0.953,校正R2分別為0.949和0.948。

表5 形態性狀對體質量的決定系數Table 5 Determination coefficient of morphological traits to body mass

表6 形態性狀與體質量的復相關分析Table 6 Multiple correlation analysis for morphometric traits to body mass
2.5 形態性狀對體質量的回歸方程
利用逐步多元回歸方法建立以9個形態性狀為自變量,體質量為因變量的回歸方程如下:
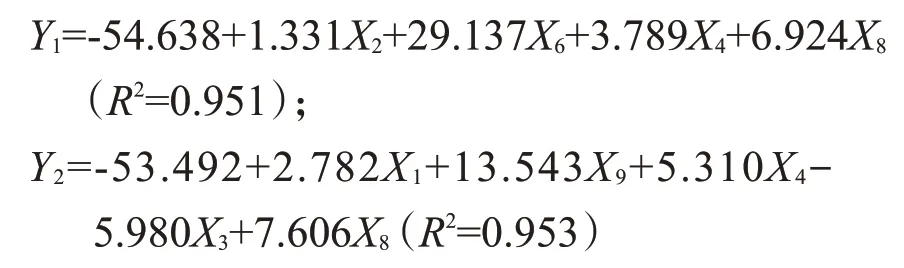
式中,Y1、Y2分別代表短圓體型和長體型群體的體質量,X1、X2、X3、X4、X6、X8、X9分別代表全長、體長、頭長、體高、尾柄高、頭高和體寬。
經統計分析,短圓體型和長體型群體回歸方程的F值分別為427.056和188.558,顯著性檢驗均呈極顯著差異(表7)。通徑分析和多元逐步回歸分析所得結果與偏回歸分析一致,表明建立的回歸方程可靠性強。
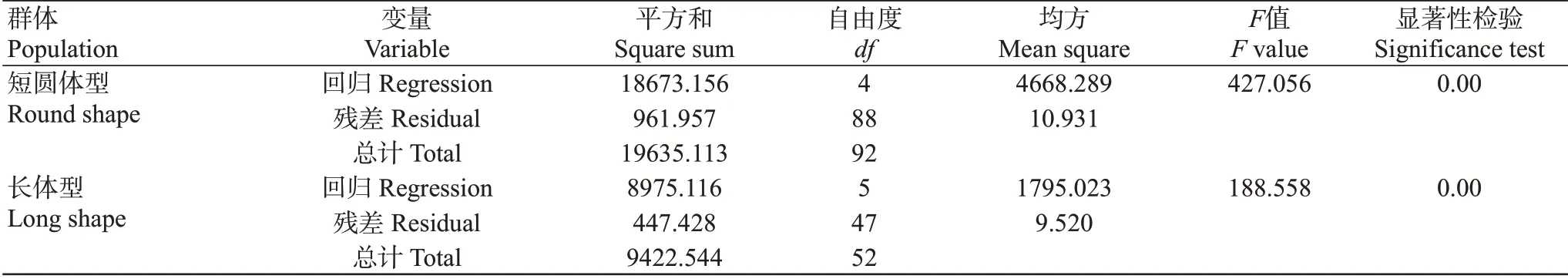
表7 形態性狀與體質量回歸關系的方差分析Table 7 ANOVA of regression relationship between morphometric traits and body mass
2.6 各形態性狀與體質量的灰色關聯度分析結果
稻田養殖禾花鯉形態性狀與體質量的灰色關聯度分析結果如表8所示。短圓體型和長體型灰色關聯系數平均值分別為0.827~0.860和0.760~0.803。從同一形態性狀來看,短圓體型群體與體質量的關聯系數平均值均大于長體型群體。分析2個群體的體質量與形態性狀的關聯度,結果如表9所示。短圓體型群體9個形態性狀的關聯序為體高>頭長>體長>尾柄高>頭高>全長>體寬>尾長>尾柄長;長體型群體9個形態性狀的關聯序為尾柄長>體高>尾柄高>全長>體寬>尾長>體長>頭高>頭長。

表8 9個形態性狀與體質量的灰色關聯系數分析Table 8 Gray relation coefficient analysis of nine morphologi‐cal traits and body mass
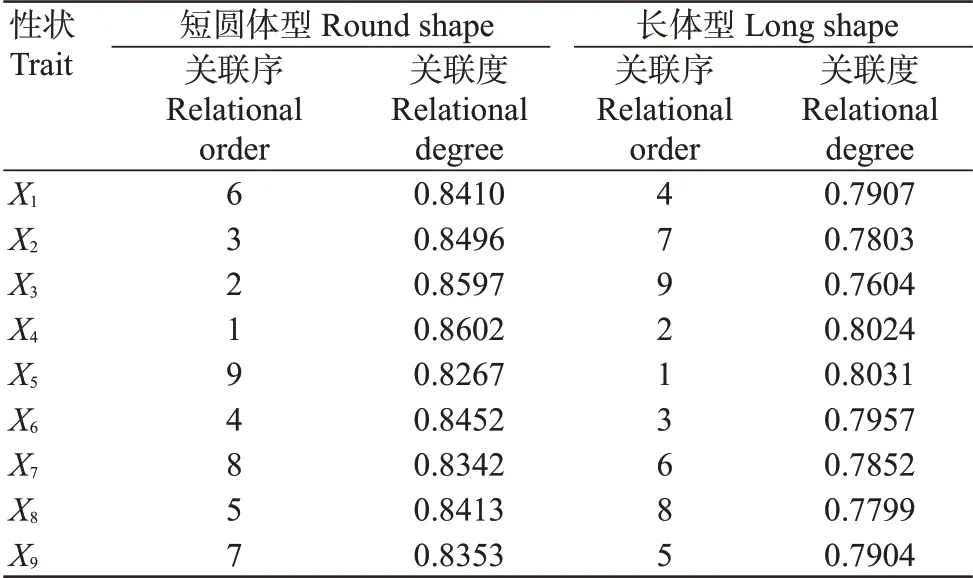
表9 9個形態性狀對體質量的灰色關聯度Table 9 Gray relation degree of nine morphological trait to body mass
3 討論
3.1 稻田養殖禾花鯉形態選育目標的確定
由于我國幅員遼闊,歷史悠久,養殖的鯉魚已形成多個群體,在形態學上也存在明顯差異。如體型細長的黑龍江野鯉(劉明華,1999)、體型修長的黃河鯉(秦改曉等,2019)、紡錘形的興國紅鯉及體型粗短的荷包紅鯉(明俊超等,2009)等,且體型性狀均可穩定遺傳(明俊超等,2009)。廣東北部山區稻田養殖的禾花鯉體長/體高值為1.9~3.0,體型變化較大,說明雖然地處山區,但該地區養殖的鯉魚未經過人工選擇,加之可能有不同的鯉魚種質引進,所以種質非常混雜。如若作為養殖品種,急需進行品種純化和改良。本研究隨機取146尾稻田養殖的禾花鯉,其中短圓體型禾花鯉占群體樣本數的63.70%。這與廣東北部地區養殖戶反映的短圓體型禾花鯉與長體型禾花鯉相比在養殖過程中逃逸率低、成活率高的現象相匹配,說明短圓體型可成為稻田養殖用禾花鯉選育的目標之一。
在水產動物遺傳育種中,表型形態性狀的變異程度通常可作為人工選育時選育目標性狀確定的重要依據。某個性狀的變異程度越大,那么該性狀的選擇潛力也越大(張成松等,2013)。本研究結果表明,廣東北部地區傳統稻田養殖的禾花鯉短圓體型和長體型禾花鯉群體體質量變異系數分別為44.51%和43.90%。而黃河鯉F1代的體質量變異系數僅為7.827%(劉曉敏和石英,2015);同為鯉科的全州禾花鯉和融水金邊禾花鯉(Procypris carpio)體質量變異系數為23.76%和9.42%(周康奇等,2020),與上述鯉相比,廣東北部地區的禾花鯉體質量選育潛力較高。而對4種體色甌江彩鯉體質量的變異系數超過68.03%以上(陳紅林等,2019);青海湖裸鯉(Gymno‐cypris przewalskii)體質量變異系數為45%(王朝溪等,2015),說明廣東北部地區的禾花鯉體質量變異程度仍處于鯉科魚類的體質量變異范圍之內。另一方面,本研究發現短圓體型和長體型群體在體質量上差異不顯著,說明2種體型的禾花鯉只有體型差異,生長速度無顯著差異。綜上所述,短圓體型禾花鯉因生長速度和長體型無顯著差異,但具有成活率高等優點,可作為下一步選育目標。
3.2 禾花鯉形態性狀對體質量的影響分析
本研究分別對2種體型的禾花鯉體質量與形態性狀間進行相關分析,結果顯示2個群體內所測量的10個生長性狀間均達極顯著相關。而通徑分析結果表明短圓體型群體中對體質量起主要作用的形態性狀是體長、尾柄高、體高和頭高;在長體型群體中對體質量起主要作用的是全長、體寬、體高、頭長和頭高。通過計算短圓體型和長體型群體形態性狀對體質量的R2分別為0.951和0.953,而校正R2分別為0.949和0.948,當R2≥0.850時,說明已找到影響依變量的主要自變量。對短圓體型禾花鯉群體體質量起主要作用的形態性狀進行通徑系數分析,結果表明除尾柄高外,體長、體高和頭高均為間接通徑系數大于直接通徑系數。而對長體型群體體質量起主要作用的形態性狀進行通徑系數分析,結果表明5個形態性狀均為間接通徑系數大于直接通徑系數。這與4種體色甌江彩鯉形態性狀與體質量的通徑分析結果類似,均為間接通徑系數大于直接通徑系數(陳紅林等,2019),表明對禾花鯉體質量起主要作用的形態性狀主要是間接作用為主導。
短圓體型群體的尾柄高對體質量的直接決定系數最大,為0.289,尾柄高和體高對群體的共同決定系數最大,為0.127;長體型群體的全長對體質量的直接決定系數最大,為0.166,全長和體寬對群體的共同決定系數最大,為0.224。分析結果表明傳統未經選育的禾花鯉群體中,與短圓體型禾花鯉體質量最相關的是魚體縱向高度的生長性狀(尾柄高和體高),而與長體型禾花鯉體質量最相關的是魚體橫向和深度的生長性狀(全長和體寬)。對全州禾花鯉群體全長對體質量的單獨決定系數最大,為0.314,體高和全長對體質量的共同決定系數最高,為0.275;融水金邊禾花鯉全長對體質量的單獨決定系數最大,為0.214,全長和頭長對體質量的共同決定系數最高,為0.262(周康奇等,2020)。巖原鯉群體全長對體質量的單獨決定系數最大,為0.426,全長和體寬對體質量的共同決定系數最大,為0.245(曲煥韜等,2018)。綜合對比分析發現,對鯉科魚類體質量產生影響的主要形態性狀的差異不僅與品種和生長環境等因素密切相關,還與群體中不同個體的體型差異相關,在分析同一群體的體質量與生長性狀的相關性之前要分析群體中個體體型的變異程度,可增加分析的準確程度。
本研究同時利用通徑分析法和灰色關聯分析法分析短圓體型和長體型禾花鯉形態性狀對體質量影響,選取通徑分析保留的形態性狀與灰色關聯分析關聯系數排名前5的形態性狀的交集,結果顯示短圓型體型群體與體質量最相關的形態性狀為體長、尾柄高、體高和頭高,而長體型群體與體質量最相關的形態性狀為全長、體寬和體高,兩種分析結果一致性較好。因灰色關聯分析不考慮自變量間相互影響,僅單純的分析各自變量與因變量之間關聯系數,所以分析結果可靠性不如通徑分析方法所分析的結果。但灰色關聯分析不受取樣數量限制,本研究結果也證實應用通徑分析方法保留的主要形態性狀與通過灰色關聯分析方法排名前5的生長性狀基本一致性,說明該方法準確性也相當高。考慮在良種選育過程中后備親本選留是按照選育目標性狀及育種值排名進行選擇,因選留親本存在人為選擇因素,導致群體很難符合正態分布且數量相對較少,這時候如果分析性狀間關聯程度,灰色關聯分析有很大應用優勢。
4 結論
兩種體型群體分別取通徑分析保留的形態性狀與灰色關聯分析關聯系數前5名形態性狀的交集,表明短圓型體型群體與體質量最相關的形態性狀為體長、尾柄高、體高和頭高,而長體型群體與體質量最相關的形態性狀為全長、體寬和體高,說明兩種分析結果一致性較好。
猜你喜歡
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
中國生殖健康(2019年2期)2019-08-23 08:12:08
電子制作(2018年18期)2018-11-14 01:48:24
產品可靠性報告(2017年7期)2017-09-05 09:49:12
山東工業技術(2016年15期)2016-12-01 05:31:22
汽車觀察(2016年3期)2016-02-28 13:16:26
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
中國質量與標準導報(2014年1期)2014-02-28 22:21:28
