福建大頭蛙抗癌多肽FJ-2945抗腫瘤活性研究
2022-07-13 07:51:56林婷婷廖偉堅(jiān)林平發(fā)潘雪豐
林婷婷,廖偉堅(jiān),林平發(fā),潘雪豐
(福建衛(wèi)生職業(yè)技術(shù)學(xué)院藥學(xué)院,福建 福州 350101)
0 引言
惡性腫瘤是威脅人類健康的疾病之一,在全球范圍內(nèi)的發(fā)生率和死亡率呈逐年上升的趨勢(shì)[1]. 其中,肝細(xì)胞癌(hepatocellular carcinoma,HCC)是臨床中最常見(jiàn)的肝臟惡性腫瘤亞型,導(dǎo)致HCC發(fā)病的主要因素有酗酒、乙肝病毒感染以及代謝性疾病等[2]. 我國(guó)每年新增的HCC患者約占全球每年新增病例的50%,屬于我國(guó)高發(fā)性腫瘤[3],給我國(guó)公共衛(wèi)生系統(tǒng)帶來(lái)巨大威脅. 對(duì)于進(jìn)展期HCC患者,臨床上通常可采用手術(shù)切除病灶、肝移植、射頻消融及經(jīng)導(dǎo)管肝動(dòng)脈化療栓塞等治療方案. 然而早期HCC沒(méi)有特征性臨床表現(xiàn),難以被及時(shí)診斷并進(jìn)行治療,多數(shù)患者初診即為中晚期腫瘤并伴隨癌細(xì)胞轉(zhuǎn)移. 針對(duì)這類患者,分子靶向藥物例如酪氨酸激酶抑制劑(tyrosine kinase inhibitors,TKIs)能取得一定的治療效果,但長(zhǎng)期使用一、二代TKIs依然伴隨著獲得性耐藥等問(wèn)題[4-5],阻礙了HCC的治療效果. 因此探索療效顯著且毒副作用小的抗腫瘤藥物,是改善肝細(xì)胞癌患者生活質(zhì)量、延長(zhǎng)生存期的必然要求.
多肽通常由10~100個(gè)氨基酸分子脫水縮合,其廣泛存在于動(dòng)植物、真菌及細(xì)菌等生物體內(nèi). 在無(wú)尾兩棲類的皮膚分泌液中發(fā)現(xiàn)多種具有抗腫瘤活性的多肽,其均具有正電性、疏水性和兩親性等特性, 可與帶有陰離子的腫瘤細(xì)胞膜產(chǎn)生選擇性[6]. 同時(shí),生物活性肽具有抑制細(xì)胞增殖、抗微管蛋白聚合、細(xì)胞毒性等功能,呈現(xiàn)出抗癌潛能[7]. 國(guó)內(nèi)外學(xué)者已從兩棲類動(dòng)物的皮膚分泌物中分離獲得數(shù)百種生物活性分子,也陸續(xù)發(fā)現(xiàn)了一系列具有抗腫瘤活性的多肽. 研究發(fā)現(xiàn),蟾蜍皮膚提取液中的Bufadienolides、多肽和生物堿,具有抗癌的藥理活性[8],印度蟾蜍皮膚水提液中細(xì)胞毒蛋白(BMP1)對(duì)癌細(xì)胞具有抑制增殖和凋亡的活性[9],一些具有抗菌活性的天然肽(magainin II,蛙皮素II)也具有殺瘤的作用[10],福建大頭蛙皮膚分泌物中提取的抗菌多肽 LFB 同時(shí)具有抗癌的活性[11]. 臨床上使用的華蟾素注射液在治療原發(fā)性肝癌、中晚期肺癌等惡性腫瘤中取得良好的療效,也證實(shí)了蛙類分泌物可作為潛在治療癌癥的新藥物來(lái)源[12].
本課題對(duì)福建本土自然資源福建大頭蛙(Limnonectesfujianensis)皮膚分泌物中含有的多肽進(jìn)行分離分析,從中發(fā)現(xiàn)多肽FJ-2945具有抑制肝癌細(xì)胞增殖、遷移和促進(jìn)細(xì)胞凋亡的作用,通過(guò)系統(tǒng)研究其體外的抗腫瘤活性,初步探討其抗腫瘤作用機(jī)制,為抗腫瘤多肽FJ-2945的臨床前研究提供理論基礎(chǔ)以及實(shí)驗(yàn)依據(jù).
1 材料與方法
1.1 材料
Hep3b購(gòu)自上海細(xì)胞生物研究所; DMEM培養(yǎng)基購(gòu)自上海HyClone公司; FBS購(gòu)自鄭州博賽生物技術(shù)股份有限公司; RNA提取試劑盒,逆轉(zhuǎn)錄試劑盒購(gòu)自諾唯贊生物技術(shù)有限公司; 引物序列由上海生工公司合成; CCK8試劑盒、RIPA裂解液、PMSF蛋白酶抑制劑、BCA蛋白濃度測(cè)定試劑盒、SDS-PAGE蛋白上樣緩沖液、分子量標(biāo)準(zhǔn)彩色預(yù)染蛋白(15~120 ku)等購(gòu)自碧云天生物技術(shù)有限公司; Skp2、p21、p27、E-cadherin和N-cadherin蛋白抗體購(gòu)自上海Cell Signaling Technology公司; cyclin A2與二抗購(gòu)自上海Santa Cruz公司.
1.2 方法
1.2.1蛙皮多肽序列與cDNA序列鑒定
蒸餾水沖洗蛙皮,釆用電刺激法(10 V,20 Hz)點(diǎn)式刺激背部,待產(chǎn)生白色沫狀物質(zhì)即用去離子水沖洗分泌物,并將所得分泌物溶液高速離心后冷凍干燥,保存于-20 ℃冰箱備用. 將5 mg凍干的福建大頭蛙蛙皮分泌物重新溶解后離心收集上清液,通過(guò) RP-HPLC對(duì)蛙皮中的組分以1 mL·min-1的流速進(jìn)行總計(jì)240 min的梯度洗脫. 初始流動(dòng)相為TFA/H2O (0.05/99.95, 體積比),終止流動(dòng)相為TFA/ACN/H2O (0.05/80/19.95, 體積比). 洗脫過(guò)程中持續(xù)監(jiān)測(cè)柱流出物在λ=214 nm 處的吸收峰,并使用自動(dòng)收集餾分收集洗脫組分. 洗脫組分使用 LCQ-Fleet電噴霧離子阱質(zhì)譜儀通過(guò) MS/MS碎片測(cè)序確定洗脫組分中FJ-2945的氨基酸序列. 根據(jù)上述步驟得到的FJ-2945的氨基酸序列設(shè)計(jì)簡(jiǎn)并引物,通過(guò)5‘-RACE 和3’-RACE法得到包含完整閱讀框的編碼相應(yīng)活性多肽的 cDNA 序列,通過(guò)與在線多肽數(shù)據(jù)庫(kù)(NCBI-Blast)對(duì)比確認(rèn)FJ-2945成熟肽序列后,最后委托上海生工公司通過(guò)固相蛋白質(zhì)合成系統(tǒng)合成FJ-2945,合成多肽純度達(dá)98%以上,置于-20 ℃保存并用于后續(xù)功能試驗(yàn)研究,臨用前用細(xì)胞培養(yǎng)基配制成所需濃度.
1.2.2細(xì)胞培養(yǎng)
將Hep3b細(xì)胞培養(yǎng)在完全培養(yǎng)基DMEM (10%FBS+1%P/S,體積分?jǐn)?shù))中,置于37 ℃、5% (體積分?jǐn)?shù))CO2細(xì)胞培養(yǎng)箱中進(jìn)行常規(guī)傳代培養(yǎng).
1.2.3CCK8實(shí)驗(yàn)
將對(duì)數(shù)生長(zhǎng)期的Hep3b細(xì)胞消化離心后稀釋至8 000個(gè)/孔的密度接種到96孔板中,置于孵箱中培養(yǎng)12 h 后,待細(xì)胞貼壁后,在實(shí)驗(yàn)組中加入不同終濃度(1、10、100 nmol·L-1和1、10、20、40、80、160 μmol·L-1)的多肽FJ-2945,每組設(shè)5個(gè)復(fù)孔,繼續(xù)培養(yǎng)24 h,隨后每孔加入10 μL的 CCK8試劑,繼續(xù)孵育2 h后在450 nm處測(cè)定吸光度并計(jì)算相對(duì)抑制率.
1.2.4RTCA技術(shù)
將對(duì)數(shù)生長(zhǎng)期的Hep3b細(xì)胞消化離心后稀釋至3 000個(gè)/孔的密度接種至實(shí)時(shí)無(wú)標(biāo)記細(xì)胞分析儀檢測(cè)盤(E-Plate)中,連續(xù)監(jiān)測(cè)76 h,每1 h記錄一次. 種盤12 h后,取出E-Plate,實(shí)驗(yàn)組加入100 μL終濃度為1、2 μmol·L-1FJ-2945的培養(yǎng)基,對(duì)照組加入等體積含有PBS的培養(yǎng)基并繼續(xù)培養(yǎng)至監(jiān)測(cè)結(jié)束,試驗(yàn)結(jié)束后,記錄實(shí)時(shí)傳輸數(shù)據(jù)并繪制細(xì)胞生長(zhǎng)情況.
1.2.5WoundHealing劃痕實(shí)驗(yàn)
將 Hep3b細(xì)胞以90%密度接種于6孔板中,待細(xì)胞充分貼壁后,用10 μL槍頭垂直于皿底制造劃痕,每制造一條劃痕換一次槍頭,隨后用 PBS洗3次,實(shí)驗(yàn)組加入100 μL終濃度為1 μmol·L-1FJ-2945的培養(yǎng)基,對(duì)照組加入等體積含有PBS的培養(yǎng)基,置于恒溫孵箱培養(yǎng)24 h,分別在孵育0、12、24 h顯微鏡下拍照記錄劃痕的愈合情況.
1.2.6Transwell實(shí)驗(yàn)
將Hep3b細(xì)胞用無(wú)血清培養(yǎng)液處理24 h后,實(shí)驗(yàn)組加入100 μL終濃度為1 μmol·L-1FJ-2945的培養(yǎng)基,對(duì)照組加入等體積含有PBS的培養(yǎng)基,繼續(xù)培養(yǎng)24 h,消化離心,無(wú)血清培養(yǎng)基重懸細(xì)胞,按4×105個(gè)/孔的密度接種于小室中,下室加入500 μL含10%(體積分?jǐn)?shù))FBS的培養(yǎng)基,置于恒溫孵箱中培養(yǎng)24 h,取出小室清潔并加入1 mL 4%(體積分?jǐn)?shù))多聚甲醛固定15 min,棄固定液,倒置風(fēng)干后,PBS洗3次,0.1%(體積分?jǐn)?shù))結(jié)晶紫染色15 min,棄染色液,PBS洗3次,顯微鏡觀察拍照,隨機(jī)選取3個(gè)細(xì)胞均勻穿膜的視野統(tǒng)計(jì)并比較對(duì)照組與實(shí)驗(yàn)組的穿膜細(xì)胞數(shù)量.
1.2.7細(xì)胞周期檢測(cè)
將對(duì)數(shù)生長(zhǎng)期的Hep3b細(xì)胞以50%~60%的密度接種至6 cm培養(yǎng)皿中,貼壁培養(yǎng)后,實(shí)驗(yàn)組加入100 μL終濃度為1、2 μmol·L-1FJ-2945的培養(yǎng)基,對(duì)照組加入等體積含有PBS的培養(yǎng)基繼續(xù)培養(yǎng)24 h,胰酶消化離心后,PBS 清洗2次,隨后加入預(yù)冷的70%(體積分?jǐn)?shù))乙醇固定過(guò)夜,離心棄去上清液,PBS清洗1次,加入PI/RNase染色緩沖液室溫避光染色30 min,隨后流式細(xì)胞儀檢測(cè)細(xì)胞熒光,觀察細(xì)胞周期分布.
1.2.8qRT-PCR實(shí)驗(yàn)
實(shí)驗(yàn)組加入100 μL終濃度為1、2 μmol·L-1FJ-2945的培養(yǎng)基,分別用實(shí)驗(yàn)組1、2表示; 對(duì)照組加入等體積含有PBS的培養(yǎng)基,培養(yǎng)24 h后,使用RNA提取試劑盒提取細(xì)胞總RNA,測(cè)定濃度后,進(jìn)行逆轉(zhuǎn)錄反應(yīng)獲得cDNA,隨后將cDNA 模板與Master Mix、待測(cè)基因的正向引物和反向引物混合的反應(yīng)體系進(jìn)行qRT-PCR反應(yīng),以檢測(cè)目的基因mRNA表達(dá)情況的變化.
1.2.9WesternBlotting實(shí)驗(yàn)
實(shí)驗(yàn)組加入100 μL終濃度為1、2 μmol·L-1FJ-2945的培養(yǎng)基(實(shí)驗(yàn)組1、2),對(duì)照組加入等體積含有PBS的培養(yǎng)基,培養(yǎng)24 h后,消化并收集細(xì)胞,提取總蛋白,BCA 試劑盒測(cè)定蛋白濃度,采用上樣緩沖液(5×)將樣品稀釋至20 μg·μL-1. 將等量的細(xì)胞蛋白通過(guò)SDS-PAGE凝膠分離并濕轉(zhuǎn)至PVDF膜,用脫脂牛奶封閉 1 h,加入一抗(1∶3 000,體積比)4 ℃孵育過(guò)夜,TBST清洗殘余一抗后,加入二抗室溫孵育1 h,TBST洗滌,最后在PVDF膜上滴加發(fā)光液進(jìn)行發(fā)光檢測(cè).
1.2.10統(tǒng)計(jì)學(xué)分析
使用GraphPad Prism 9.0軟件進(jìn)行統(tǒng)計(jì)學(xué)分析,計(jì)量資料用平均值±標(biāo)準(zhǔn)差(x±s)表示,組間均數(shù)比較采用t檢驗(yàn),P<0.05表示差異具有統(tǒng)計(jì)學(xué)意義.
2 實(shí)驗(yàn)結(jié)果與分析
2.1 FJ-2945的多肽序列與 cDNA 序列鑒定
圖1(a)為RP-HPLC梯度洗脫結(jié)果曲線. LCQ-MS/MS 鑒定#121洗脫峰中含有相對(duì)分子質(zhì)量為2 945的多肽組分,通過(guò)在線多肽序列分析工具預(yù)測(cè)得到的的FJ-2945碎片化單電荷和雙電荷 b-ion和片段的分子質(zhì)量表,質(zhì)譜中檢測(cè)到的b-ion碎片用紅色字體標(biāo)記,而y-ion用藍(lán)色標(biāo)記,驗(yàn)證其多肽一級(jí)序列為TLKNLAKTAGKGALQSLLNHASCKLSGQC(見(jiàn)圖1(b)). 測(cè)序得到FJ-2945的開(kāi)放閱讀框 cDNA(見(jiàn)圖1(c)),與數(shù)據(jù)庫(kù)(NCBI-Blast)對(duì)比確認(rèn)FJ-2945成熟肽序列,說(shuō)明蛙皮中存在編碼FJ-2945的核苷酸序列,應(yīng)用 SWISS-MODEL 多肽二級(jí)結(jié)構(gòu)預(yù)測(cè)軟件預(yù)測(cè)得到FJ-2945多肽具有兩親性的α螺旋二級(jí)結(jié)構(gòu)(α-Helix)(見(jiàn)圖1(d)). 根據(jù)文獻(xiàn)報(bào)道,兩親性的α螺旋多肽常具有抗菌與抗腫瘤增殖作用[13].

圖1 FJ-2945分離與結(jié)構(gòu)鑒定Fig.1 Isolation and structure identification of FJ-2945
2.2 FJ-2945對(duì)Hep3b細(xì)胞生長(zhǎng)與增殖的抑制作用
CCK8實(shí)驗(yàn)結(jié)果表明,多肽FJ-2945呈劑量依賴性的抑制Hep3b細(xì)胞的增殖過(guò)程,其半數(shù)抑制濃度(IC50)為(1.87±0.499)μmol·L-1(見(jiàn)圖2(a)). 隨后,通過(guò)RTCA技術(shù)同樣驗(yàn)證多肽FJ-2945對(duì)Hep3b細(xì)胞的增殖呈劑量依賴性的抑制作用,在細(xì)胞培養(yǎng)76 h后,實(shí)驗(yàn)組中Hep3b細(xì)胞增殖速度顯著低于對(duì)照組,接種2 μmol·L-1FJ-2945的實(shí)驗(yàn)組2細(xì)胞總量?jī)H為對(duì)照組細(xì)胞的50%,實(shí)驗(yàn)組1細(xì)胞總量為對(duì)照組細(xì)胞的83%,表明FJ-2945對(duì)Hep3b細(xì)胞的生長(zhǎng)增殖具有較強(qiáng)的抑制作用(見(jiàn)圖2(b)).
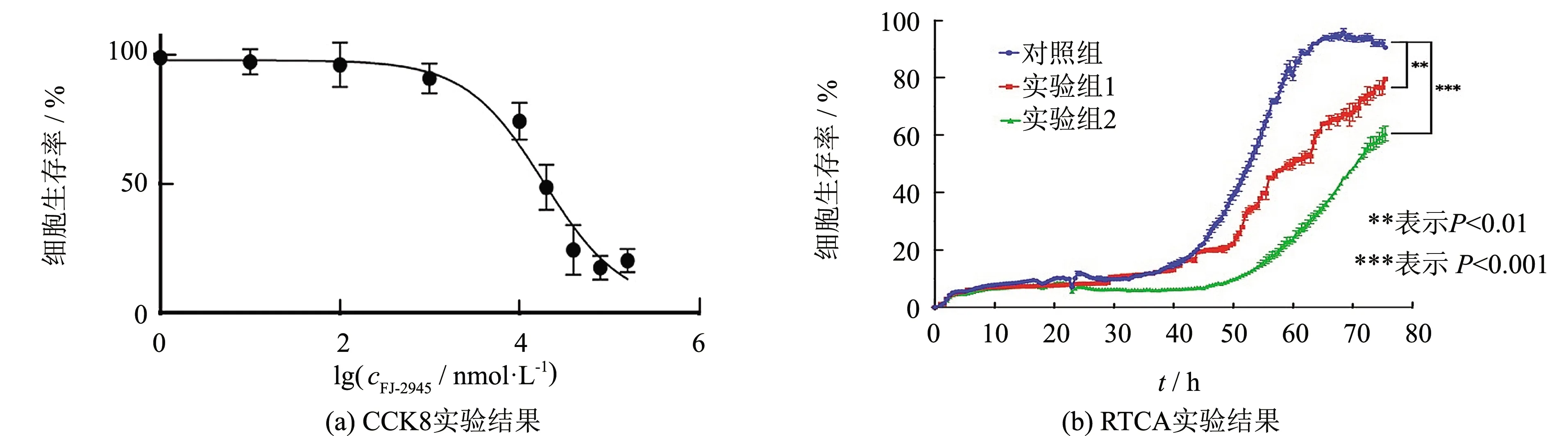
圖2 FJ-2945對(duì)Hep3b細(xì)胞增殖的影響Fig.2 Effect of FJ-2945 on the proliferation of Hep3b cell
2.3 FJ-2945對(duì)Hep3b細(xì)胞周期分布的影響
流式細(xì)胞儀檢測(cè)結(jié)果表明,實(shí)驗(yàn)組中Hep3b細(xì)胞呈現(xiàn)細(xì)胞周期G1/S期阻滯,并以2 μmol·L-1多肽FJ-2945 (實(shí)驗(yàn)組2)的作用更為顯著(見(jiàn)圖3(a~c)). 如圖3(d)統(tǒng)計(jì)圖中所示,經(jīng)1、2 μmol·L-1多肽FJ-2945處理后,G0/G1期細(xì)胞數(shù)量提高至66.8%、70.9%,相比對(duì)照組60.7%有不同程度的提高. 此外,經(jīng)過(guò)2 μmol·L-1FJ-2945處理后,處于sub-G1期的細(xì)胞數(shù)量由對(duì)照組中的5.12%提高到10.95%,說(shuō)明FJ-2945可能同時(shí)誘導(dǎo)細(xì)胞凋亡.
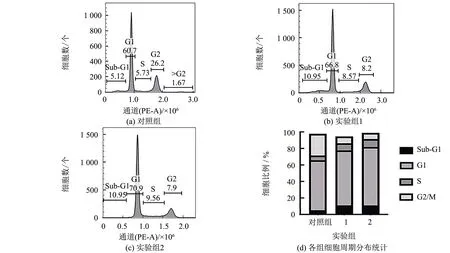
圖3 FJ-2945對(duì)Hep3b細(xì)胞周期的影響Fig.3 Effect of FJ-2945 on the cell cycle of Hep3b
2.4 FJ-2945對(duì)Hep3b細(xì)胞遷移的影響
劃痕實(shí)驗(yàn)和Transwell實(shí)驗(yàn)檢測(cè)多肽FJ-2945對(duì)Hep3b遷移能力的影響. 為避免因高濃度FJ-2945造成的細(xì)胞殺傷作用導(dǎo)致假陽(yáng)性結(jié)果,選擇1 μmol·L-1的FJ-2945處理細(xì)胞,分別在處理后0、12、24 h進(jìn)行拍照記錄. 結(jié)果如圖4(a)所示,實(shí)驗(yàn)組中Hep3b細(xì)胞遷移速率明顯低于對(duì)照組. 對(duì)照組在劃傷24 h后劃痕基本愈合,而實(shí)驗(yàn)組仍保留劃痕,證實(shí)了多肽FJ-2945能明顯抑制Hep3b細(xì)胞的遷移. Transwell結(jié)果如圖4(b)所示,每個(gè)單獨(dú)視野中實(shí)驗(yàn)組Hep3b細(xì)胞穿過(guò)至下室的細(xì)胞數(shù)量為(40±7)個(gè),與對(duì)照組(83±4)個(gè)相比,差異具有統(tǒng)計(jì)學(xué)意義(***表示P<0.001),提示FJ-2945能抑制Hep3b細(xì)胞的遷移能力.

圖4 FJ-2945對(duì)Hep3b細(xì)胞遷移能力的影響Fig.4 Effect of FJ-2945 on the cell migration of Hep3b
2.5 FJ-2945抗腫瘤機(jī)制研究
通過(guò)qRT-PCR檢測(cè)細(xì)胞增殖、細(xì)胞周期相關(guān)因子E2F1、Skp2、CDK4、CCNA2、CDKN1B和細(xì)胞遷移相關(guān)因子Slug、Snail、E-cadherin、N-cadherin的變化,結(jié)果如圖5(a)所示. 實(shí)驗(yàn)組中與細(xì)胞周期進(jìn)展相關(guān)的基因如Skp2、E2F1、CCNA2、CDK4均沒(méi)有發(fā)生顯著變化,細(xì)胞遷移相關(guān)的基因Slug與Snail水平顯著降低(*表示P<0.05, *** 表示P<0.001),CDKN1B、E-cadherin的mRNA水平顯著上升(***表示P<0.001),說(shuō)明多肽FJ-2945并非從轉(zhuǎn)錄水平上影響細(xì)胞周期相關(guān)因子,但是對(duì)細(xì)胞遷移調(diào)控因子E-cadherin、Slug與Snail則具有顯著影響.
Western blotting實(shí)驗(yàn)結(jié)果如圖5(b)所示,實(shí)驗(yàn)組中Skp2、Cyclin A蛋白的表達(dá)顯著下降. 有研究報(bào)道細(xì)胞中Skp2含量降低會(huì)導(dǎo)致細(xì)胞增殖抑制并誘導(dǎo)細(xì)胞凋亡[14],或解釋了FJ-2945的主要抗腫瘤分子靶標(biāo). 另一方面,實(shí)驗(yàn)組中Skp2靶蛋白p27含量則顯著上升. 據(jù)文獻(xiàn)報(bào)道,Skp2介導(dǎo)p27周期性的泛素-蛋白酶體降解,使p27釋放Cyclin A 或Cyclin E/Cdk2復(fù)合物的抑制狀態(tài),從而允許細(xì)胞通過(guò) G1/S 期[15]. 這些結(jié)果可能說(shuō)明FJ-2945是通過(guò)降低 Skp2在肝癌細(xì)胞中的蛋白含量,從而抑制了由Skp2介導(dǎo)的下游靶蛋白p27泛素-蛋白酶體降解過(guò)程,該結(jié)果導(dǎo)致細(xì)胞周期蛋白p27在細(xì)胞中積累并抑制CDK-cyclin復(fù)合體,從而造成細(xì)胞周期G1/S 期阻滯[16]. 此外,實(shí)驗(yàn)組中細(xì)胞錨定蛋白E-cadherin表達(dá)量升高而促進(jìn)細(xì)胞遷移的N-cadherin表達(dá)量降低, 則說(shuō)明FJ-2945可能通過(guò)抑制癌細(xì)胞上皮間質(zhì)轉(zhuǎn)換(EMT)轉(zhuǎn)變,從而降低肝癌細(xì)胞的遷移能力[17].

圖5 FJ-2945抗腫瘤機(jī)制研究Fig.5 Anti-cancer mechanism study of FJ-2945
3 討論
我國(guó)肝癌的發(fā)病率在惡性腫瘤中位居第四位,死亡率位居第二位,并且肝癌是所有腫瘤中預(yù)后最差的腫瘤[18]. 肝癌的診治水平可以提高患者的生存率,除了傳統(tǒng)化療藥物,加快研發(fā)新型的藥物尤為重要. 國(guó)內(nèi)外已發(fā)現(xiàn)蛙類皮膚腺分泌物中部分抗菌、抗腫瘤等生物活性成分[11, 19-20],且由于蛙皮活性多肽的相對(duì)分子質(zhì)量小、特異性強(qiáng)、毒性小等特點(diǎn)一直被認(rèn)為是極具潛力的新型抗腫瘤藥物的研究對(duì)象,但其抗腫瘤的機(jī)制多樣且尚未被徹底明確限制了其抗腫瘤的研究進(jìn)展[21]. 福建大頭蛙(Limnonectesfujianensis)屬于福建省武夷山地區(qū)的兩棲動(dòng)物優(yōu)勢(shì)種,其皮膚腺體分泌物含有大量的多肽類活性物質(zhì),本課題組從福建大頭蛙皮膚分泌物中分離并鑒定新型活性多肽FJ-2945的氨基酸與cDNA序列,并使用合成的FJ-2945多肽進(jìn)行抗腫瘤活性與抗腫瘤機(jī)制研究.
本研究結(jié)果表明,多肽FJ-2945能夠抑制Hep3b細(xì)胞的生長(zhǎng)增殖、遷移與凋亡,表明多肽FJ-2945對(duì)肝癌細(xì)胞Hep3b抑制作用以及潛在的治療價(jià)值. 在抗腫瘤機(jī)制方面,本研究明確了多肽FJ-2945通過(guò)降低細(xì)胞內(nèi)Skp2蛋白含量造成p27蛋白在胞內(nèi)累積,從而造成細(xì)胞周期的G1/S 期阻滯,最終抑制Hep3b細(xì)胞增殖. 根據(jù)文獻(xiàn)報(bào)道,在細(xì)胞增殖過(guò)程中,Skp2通過(guò)降解p27促進(jìn)細(xì)胞周期G1/S期轉(zhuǎn)換,而在細(xì)胞遷移過(guò)程中,Skp2蛋白的高表達(dá)被認(rèn)為與腫瘤增殖與遷移呈正相關(guān),其機(jī)制主要與胞質(zhì)Skp2誘導(dǎo)的細(xì)胞錨定蛋E-cadherin的過(guò)度降解有關(guān)[22-24]. 本實(shí)驗(yàn)抗腫瘤機(jī)制初步研究結(jié)果表明,多肽FJ-2945極有可能是通過(guò)影響細(xì)胞中Skp2的蛋白質(zhì)表達(dá),從而影響Skp2的下游因子如p27、E-cadherin等,最終抑制肝癌細(xì)胞的增殖與遷移能力.
綜上所述,多肽FJ-2945具有顯著的抑制Hep3b細(xì)胞增殖、遷移的作用,并誘導(dǎo)Hep3b細(xì)胞凋亡的抗腫瘤活性,但同時(shí)與其他多肽類物質(zhì)一樣也具有易水解、穩(wěn)定性差與靶向性不強(qiáng)等缺點(diǎn),因此需要在此基礎(chǔ)上對(duì)多肽FJ-2945進(jìn)行結(jié)構(gòu)改造,以期進(jìn)一步提升其穩(wěn)定性與抗腫瘤活性,通過(guò)臨床前實(shí)驗(yàn)更深入研究FJ-2945抗腫瘤作用機(jī)理,挖掘其臨床應(yīng)用的潛力.
