高效產殼聚糖酶海洋菌株Mitsuaria sp.SH-50 的篩選、鑒定及產酶條件優化
2022-07-13 08:27:22盧波斯崔丹丹李志明袁麗君沈宏
中國調味品 2022年7期
關鍵詞:殼聚糖
盧波斯,崔丹丹,李志明,袁麗君,沈宏
(華南農業大學 資源環境學院,廣州 510642)
殼聚糖由D-氨基葡萄糖通過β-1,4-糖苷鍵連接而成,是自然界中唯一存在的堿性多糖,其獨特的結構特征使其具有特殊的生理功能[1]。低聚殼聚糖則是由殼聚糖分解成聚合度在2~20之間的低聚產品[2]。目前,工業化制備低聚殼聚糖的工藝主要分為化學法、物理法和生物酶解法。化學法一般采用酸解法或氧化降解法,但酸解過程需要在高溫高壓的環境中進行,且會排放大量酸性環境污染物[3],易造成環境污染;氧化法降解時容易引起低聚殼聚糖色澤的改變,增加產品提純難度。朱新鋒等[4]通過撞擊流技術制備低聚殼聚糖,使得反應物黏度下降率達到67.6%,但通過物理技術制備的低聚糖聚合度往往較難控制,且成本相對較高。相比于其他技術,生物酶解法是指利用專一性或非專一性的酶蛋白對殼聚糖進行水解[5],具有非常好的應用前景。研究表明,利用殼聚糖酶制備的低聚殼聚糖不僅保留了殼聚糖的生理活性,而且具有更好的水溶性及良好的生物相容性,已經被廣泛地應用于提高飲品澄清度[6]、改善腸道pH、食品保鮮、生產調味品、天然防腐劑[7]、抗氧化[8]、降甘油三酯、抗炎[9]、穩定動脈粥樣硬化[10]、抵御癌癥[11]等食品及醫療領域[12]。
殼聚糖酶(EC3.2.1.132)可以高效地水解殼聚糖[13],其主要來源于真菌和細菌,也有少部分存在于植物和病毒中[14]。細菌類以芽孢桿菌屬研究最為深入,真菌類主要有曲霉菌、鏈霉菌、淡紫擬青霉、木霉菌屬等,但海洋來源的菌株相對較少。酶法制備低聚殼聚糖具有反應效率高、產品純度高、后續分離純化簡單、環境友好等優勢[15]。但目前利用生物酶解法制備低聚殼聚糖同樣也面臨著高效產殼聚糖酶原始菌株較少、殼聚糖酶普遍活力低下、菌株產酶周期長以及純酶價格昂貴的問題。因此,篩選出高效產殼聚糖酶的原始菌株對未來大規模生產制備食品級低聚殼聚糖至關重要。
1 材料與方法
1.1 材料與試劑
濱海土壤樣品:海南三亞蝦蟹養殖區;粉末殼聚糖(脫乙酰度≥95%):西安百川生物科技有限公司;膠體殼聚糖(脫乙酰度≥85%):青島博智匯力生物科技有限公司;酵母粉、蛋白胨、瓊脂粉:賽默飛世爾科技公司;氨基葡萄糖鹽酸鹽(分析純):阿拉丁公司;革蘭氏染色試劑盒:廣東環凱生物科技有限公司;Green Taq Mix:諾唯贊生物科技股份有限公司;其余試劑均為國產分析純。
培養基:富集培養基(g/L):膠體殼聚糖10.0,硫酸鎂0.5,磷酸氫二鉀0.5,磷酸二氫鉀0.5,蛋白胨3.0,酵母粉0.5,氯化鈉5.0,pH 7.0;初篩培養基(g/L):膠體殼聚糖10.0,硫酸鎂1.0,磷酸氫二鉀1.0,磷酸二氫鉀1.0,硫酸銨2.0,氯化鈉5.0,瓊脂15.0,pH 7.0;復篩培養基(g/L):粉末殼聚糖10.0,硫酸鎂1.0,磷酸氫二鉀2.0,磷酸二氫鉀1.0,硫酸銨4.0,氯化鈉5.0,pH 6.5;種子培養基(g/L):酵母粉5.0,蛋白胨10.0,氯化鈉10.0,pH 7.0。
1.2 主要儀器與設備
UV-2450分光光度計 日本Shimadzu公司;PCR儀 日本Takara公司;WIX-EP600凝膠電泳儀 韋克斯科技(北京)有限公司;BSD-YX(F)3200恒溫搖床、BPX-82電熱恒溫培養箱、BJ-1CD超凈工作臺 上海博迅實業有限公司;HC-3618R臺式高速離心機 中科中佳科學儀器有限公司;HWS-5A恒溫水浴鍋 上海百典儀器設備有限公司。
1.3 菌株篩選
1.3.1 富集
將采自海南三亞蝦蟹養殖區的10 g土壤樣品與90 mL滅菌水混合后置于三角瓶中,180 r/min振蕩1 h,靜置后取1 mL上清液接種于100 mL/250 mL含富集培養基的三角瓶中,在28 ℃、180 r/min條件下搖瓶培養3 d。
1.3.2 初篩
取富集后的菌液按梯度法稀釋101~107倍,吸取稀釋102~107倍后的溶液100 μL均勻涂布在含初篩固體培養基的培養皿上,在28 ℃條件下恒溫培養3 d,觀察培養皿上菌落生長狀況,挑取生長良好、不分泌黏液且具有透明圈的單菌落,在初篩培養基上劃線純化2~3次。
1.3.3 復篩
挑選透明圈直徑/菌落直徑(D/d)最大的單菌落進行復篩,用接菌環輕蘸菌苔后接種于100 mL/250 mL含復篩培養基的三角瓶中,在28 ℃、200 r/min條件下搖瓶培養30 h。
1.3.4 保存
取復篩培養基中菌液1 mL接種于100 mL/250 mL含種子培養基的三角瓶中,培養6 h后保存于-80 ℃冰箱中。
1.4 菌株鑒定
1.4.1 菌株形態學特征鑒定
取保存菌株在初篩培養基上劃線,30 ℃恒溫培養20 h后用接菌環挑起少量菌苔于載玻片上,用無菌水稀釋一定倍數,明火干燥成膜后依照革蘭氏染色試劑盒說明書步驟進行染色,使用光學顯微鏡觀察菌株形態。
1.4.2 菌株分子生物學鑒定
取保存菌株在初篩培養基上劃線,30 ℃恒溫培養30 h后用接菌環刮取微量菌苔加入到150 μL無菌水中煮沸10 min,12000 r/min條件下離心10 min,取上清液進行PCR擴增。采用細菌16S rDNA 通用引物(正向引物 5′-AGAGTTTGATCCTGGCTCAG-3′,反向引物5′-GGTTACCTTGTTACGACTT-3′)進行 PCR 擴增,擴增程序為:94 ℃ 5 min;98 ℃ 10 s,55 ℃ 30 s,72 ℃ 1 kb/1 min,35個循環;72 ℃ 10 min;4 ℃保存。獲得的PCR產物利用1%的瓊脂糖凝膠電泳驗證后,交由廣州天一輝遠基因科技有限公司進行測序。將測序結果提交到NCBI 數據庫,利用BLAST程序搜索同源序列,選出同源性高的菌株的16S rDNA序列為參比對象,通過MEGA 7.0 軟件的Neighbor-Joining 法構建系統發育樹[16]。
1.5 粗酶液制備
發酵液在12000 r/min、4 ℃條件下離心10 min。上清液即為粗酶液[17]。
1.6 酶活測定
采用3,5-二硝基水楊酸(DNS)法測定粗酶液的酶活[18]。取100 μL酶液和900 μL 1%殼聚糖溶液加入至25 mL比色管中,75 ℃下恒溫反應10 min后迅速加入1.5 mL DNS溶液,冷卻至室溫后沸水浴5 min顯色,加蒸餾水定容至25 mL;在9000 r/min、4 ℃條件下離心10 min,取上清液在540 nm處進行比色,測定酶活。
1.7 產酶條件優化
1.7.1 單因素試驗設計
在復篩培養基的基礎上分析單一組分變化對SH-50菌株產酶活性的影響,影響因素主要包括:碳源的種類及濃度、氮源的種類及濃度、無機鹽的種類及濃度、初始發酵pH、發酵溫度、發酵時間。
1.7.2 Plackett-Burman(PB)試驗設計
在獲得單因素試驗結果的基礎上設計PB試驗,設計影響SH-50菌株產酶活性的7個因素,每個因素分為高水平(1)和低水平(-1),挑選置信度≥95%且影響最大的3個因素進行爬坡試驗,找到菌株產酶活性的拐點。PB試驗設計見表1。

表1 PB試驗不同因素及水平設計Table 1 Design of different factors and levels of PB test
1.7.3 Box-Behnken Design(BBD)試驗設計
設計BBD試驗(見表2),并將結果輸入Design Expert V8.0.6軟件構建回歸模型,對數據進行方差及可信度分析,根據所得最優產酶活性的編碼水平進行重復試驗,驗證回歸模型與實際產酶活性之間的相關性。

表2 BBD試驗不同因素及水平設計Table 2 Design of different factors and levels of BBD test
2 結果和分析
2.1 菌株形態學鑒定

圖1 SH-50菌株的形態學鑒定Fig.1 Morphological identification of SH-50 strain
土壤樣品在28 ℃條件下搖瓶富集3 d后,經篩選得到一株產殼聚糖酶菌株,由圖1中a可知,菌落呈乳白色,能夠降解殼聚糖,將其標記為SH-50。挑取圖1中a的SH-50單菌落在唯一碳源為殼聚糖的培養基上繼續培養生長2 d,由圖1中b可知,菌落為邊緣規則的圓形,菌苔呈乳白色,微隆起,表面光滑,易挑起,有明顯的殼聚糖水解圈。刮取微量圖1中b的菌苔進行革蘭氏染色,在顯微鏡下觀察,結果見圖1中c,細菌呈短棒狀,革蘭氏染色結果為陰性。
2.2 菌株的生理生化特征
將SH-50菌株與模式菌株M.chitosanitabida3001的生理生化特性進行比較,結果見表3。
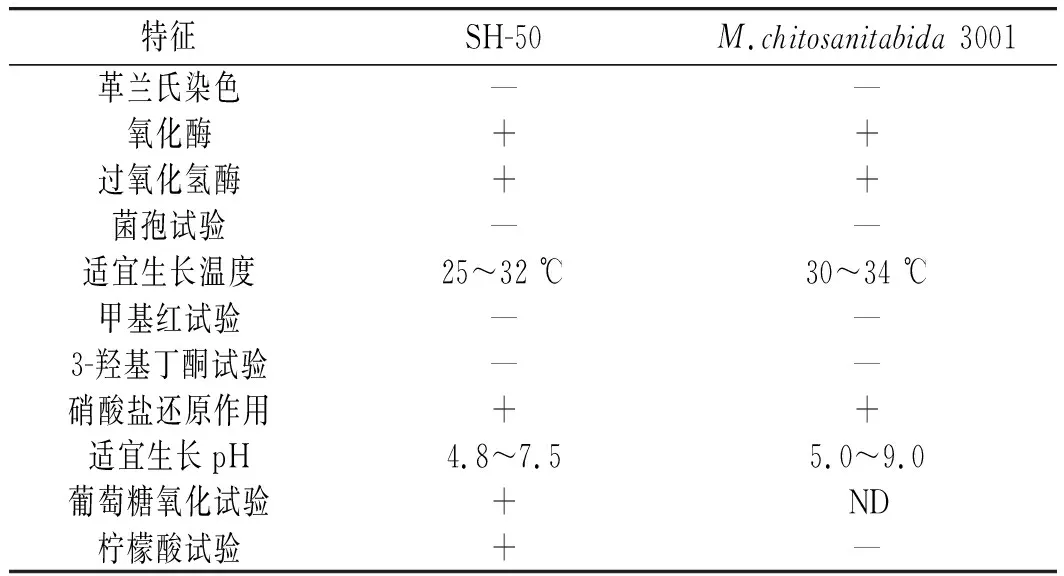
表3 SH-50菌株的生理生化性質Table 3 Physiological and biochemical properties of SH-50 strain
由表3可知,SH-50菌株的生理生化特征與模式菌株基本相同,但適宜生長溫度、pH及檸檬酸試驗存在一定差異,可能是因為生長環境不同,菌株生理生化性質有所改變,同時也表明SH-50菌株可能是松江菌屬潛在的新種。
2.3 分子生物學鑒定
利用16S rDNA通用引物對SH-50菌株進行PCR擴增后,對6個重復樣進行瓊脂糖凝膠電泳試驗,產物長度約為1500 bp,結果見圖2。
將PCR產物送至廣州天一輝遠基因科技有限公司進行測序,將得到的序列在NCBI(https://www.ncbi.nlm.nih.gov)上進行BLAST比對后發現SH-50與Mitsuariasp.菌株同源性最高。其系統進化樹見圖3,根據16S rDNA序列,可以將Mitsuariasp.分為4簇,其中SH-50與Mitsuariastrain II3親緣關系最近,位于同一簇的不同分支,因此將其命名為Mitsuariasp. SH-50,菌株16S rDNA上傳至GenBank,編號為MW911724。
2.4 菌株生長及產酶曲線的測定
取保存的SH-50菌株接入種子培養基中培養至對數生長期,按1%的接種量轉接至100 mL/250 mL含復篩培養基的三角瓶中,在28 ℃、180 r/min條件下培養,每隔3 h取樣測定菌液OD600、菌液酶活,結果見圖4。
由圖4可知,SH-50菌株在生長6 h左右時逐漸從對數生長期進入穩定期,繼續生長至12~24 h時,OD600繼續增加,此時菌體分泌大量代謝產物,在24~30 h時OD600開始下降,表明培養基營養供應不足,菌體密度下降。SH-50菌株的產酶曲線與生長曲線呈一定程度上的正相關,從OD540對應的產酶曲線可以看出,產酶活性在12 h時達到最高值,但隨著時間增加,酶液活性逐漸開始下降。因此,選取12 h作為該菌株最佳的發酵產酶時間。
2.5 培養基單因素變化對SH-50菌株產酶的影響
2.5.1 碳源對SH-50菌株產酶活性的影響
碳源主要在微生物生長代謝時為其提供細胞碳架,提供微生物生命活動所需的能量,同時為合成產物提供碳架,是細胞正常生長以及分裂的物質基礎。該研究分別以殼聚糖、蔗糖、葡萄糖、淀粉、纖維素作為SH-50菌株的唯一碳源,探究不同碳源對該菌株產殼聚糖酶活性的影響,結果見圖5。
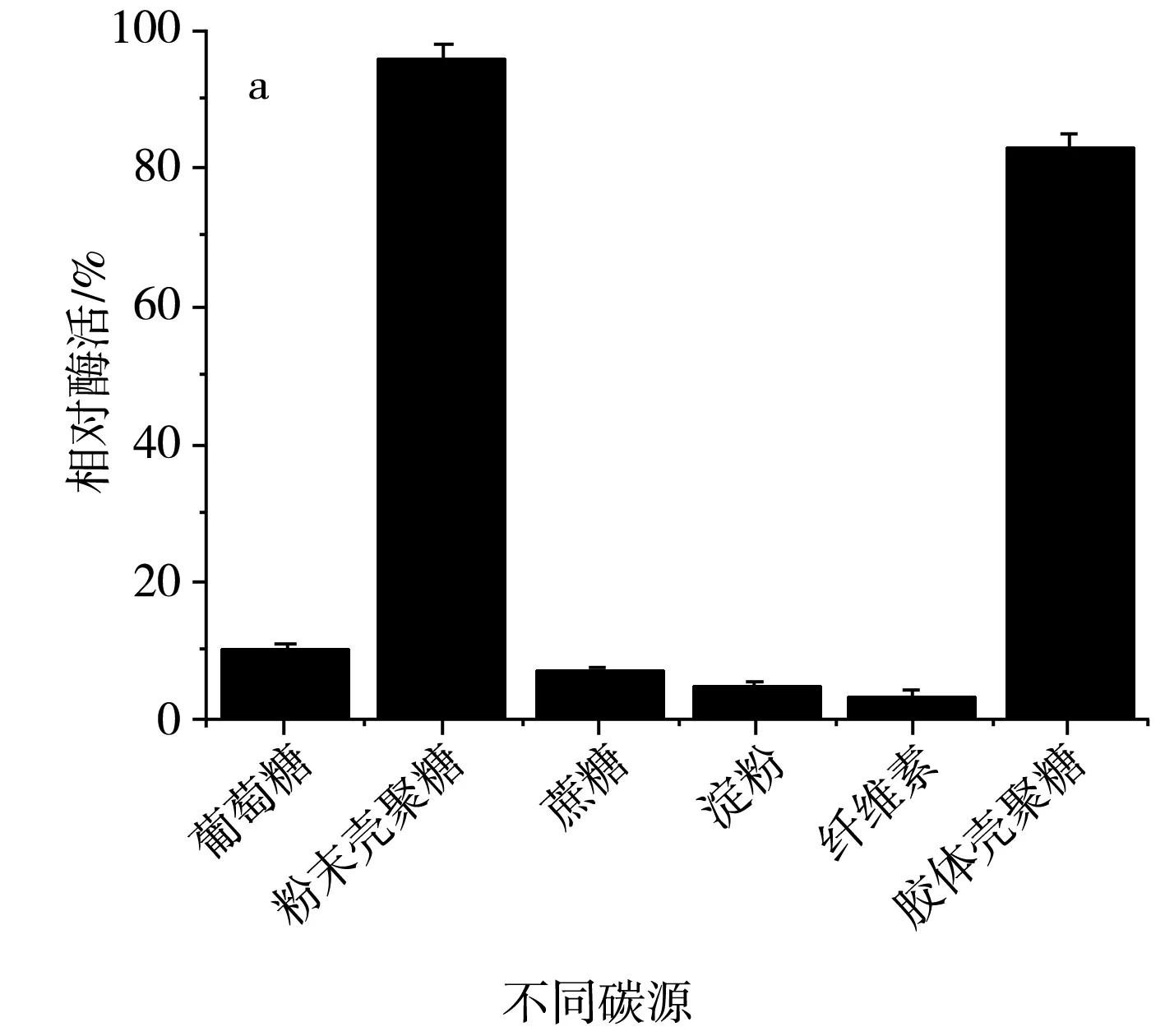
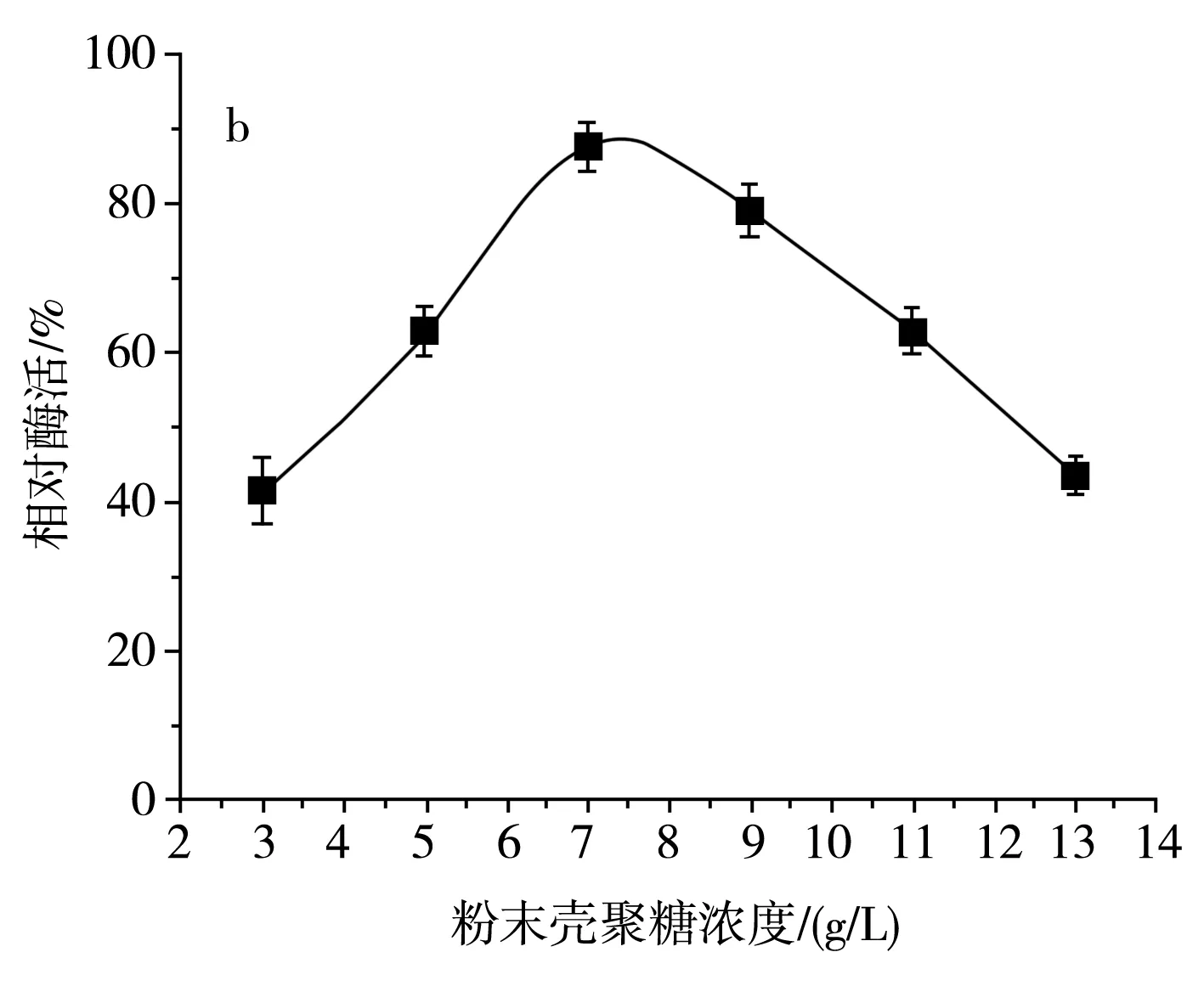
圖5 不同碳源及殼聚糖濃度對SH-50菌株產酶活性的影響Fig.5 Effects of different carbon sources and chitosanase concentrations on enzyme production activity of SH-50 strain
由圖5中a可知,SH-50菌株在碳源為殼聚糖時產酶活性較高,碳源為其他糖類時該菌株幾乎不產殼聚糖酶。為明確殼聚糖濃度對SH-50菌株產酶活性的影響,分別設置粉末殼聚糖濃度為3,5,7,9,11,13 g/L,由圖5中b可知,隨著殼聚糖濃度的增加,SH-50菌株產殼聚糖酶活性呈先上升后下降的趨勢,在殼聚糖濃度較低時,酶活性上升趨勢明顯,當殼聚糖濃度達到9 g/L時,酶活性明顯下降。最適的殼聚糖濃度應在7~8 g/L,因此,選取中間值7.5 g/L作為SH-50菌株發酵的殼聚糖濃度。
2.5.2 氮源對SH-50菌株產酶活性的影響
氮源主要用于菌體細胞物質(氨基酸、蛋白質、核酸等)和含氮代謝物的合成,同樣也是微生物生長不可或缺的物質,許多功能微生物也常常被用于水體氮污染的凈化。在設置等量氮的前提下,該研究分別選用蛋白胨、酵母粉、硫酸銨、尿素、硝酸鉀、牛肉浸膏作為SH-50菌株的唯一氮源,然后進一步確定最佳氮源的添加濃度,結果見圖6。
由圖6中a可知,在不同氮源培養條件下,SH-50菌株的唯一氮源為硫酸銨和酵母粉時菌株產酶活性較高,然而硫酸銨在培養基中易造成粉末殼聚糖析出,因此選用酵母粉作為SH-50菌株的最適氮源。由圖6中b可知,不同濃度的酵母粉對SH-50菌株所產殼聚糖酶的酶活性具有一定影響,酵母粉濃度小于1 g/L時,菌株產酶活性隨濃度增加而增加,酵母粉濃度在1~2 g/L時菌株產酶活性變化不大,綜合經濟因素,選取1 g/L作為SH-50菌株發酵的酵母粉濃度。
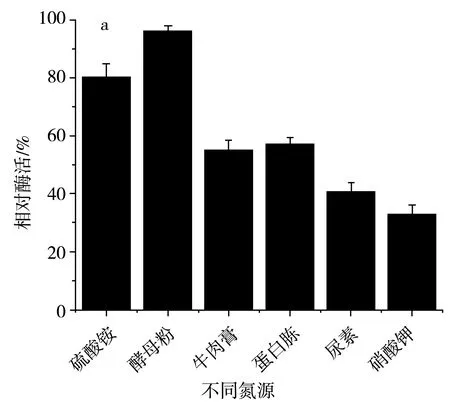
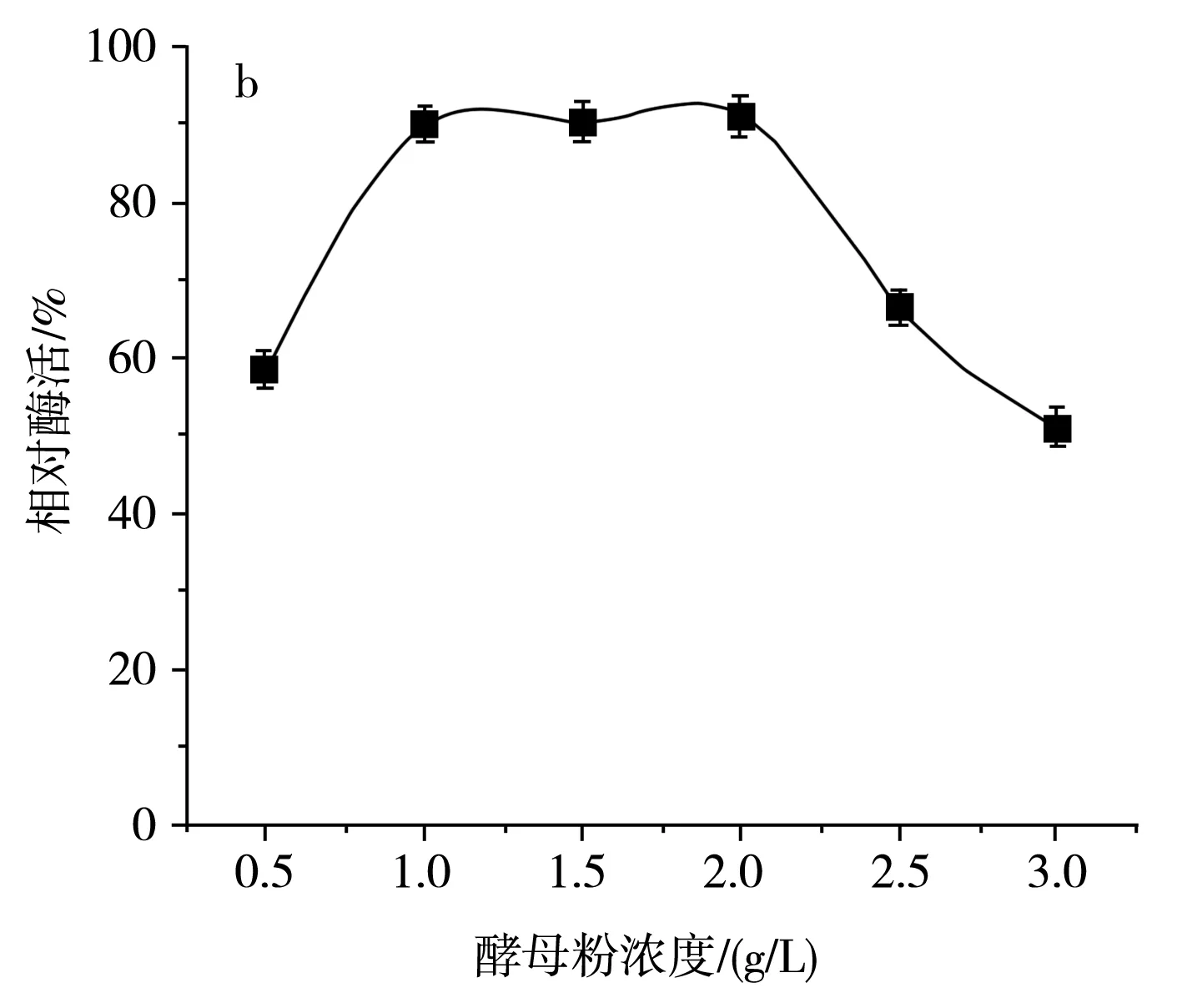
圖6 不同氮源及酵母粉濃度對SH-50菌株產酶活性的影響Fig.6 Effects of different nitrogen sources and yeast powder concentrations on enzyme production activity of SH-50 strain
2.5.3 無機鹽對SH-50菌株產酶活性的影響
無機鹽中的各類離子同樣是菌體細胞的組成成分之一,同時也可以作為酶的組成部分、酶的激活劑或抑制劑[19],具有調節細胞的滲透壓、pH、氧化還原電位等功能。該研究主要分析常規無機鹽氯化鈉及磷酸氫二鉀的濃度對SH-50菌株產酶活性的影響,結果見圖7。


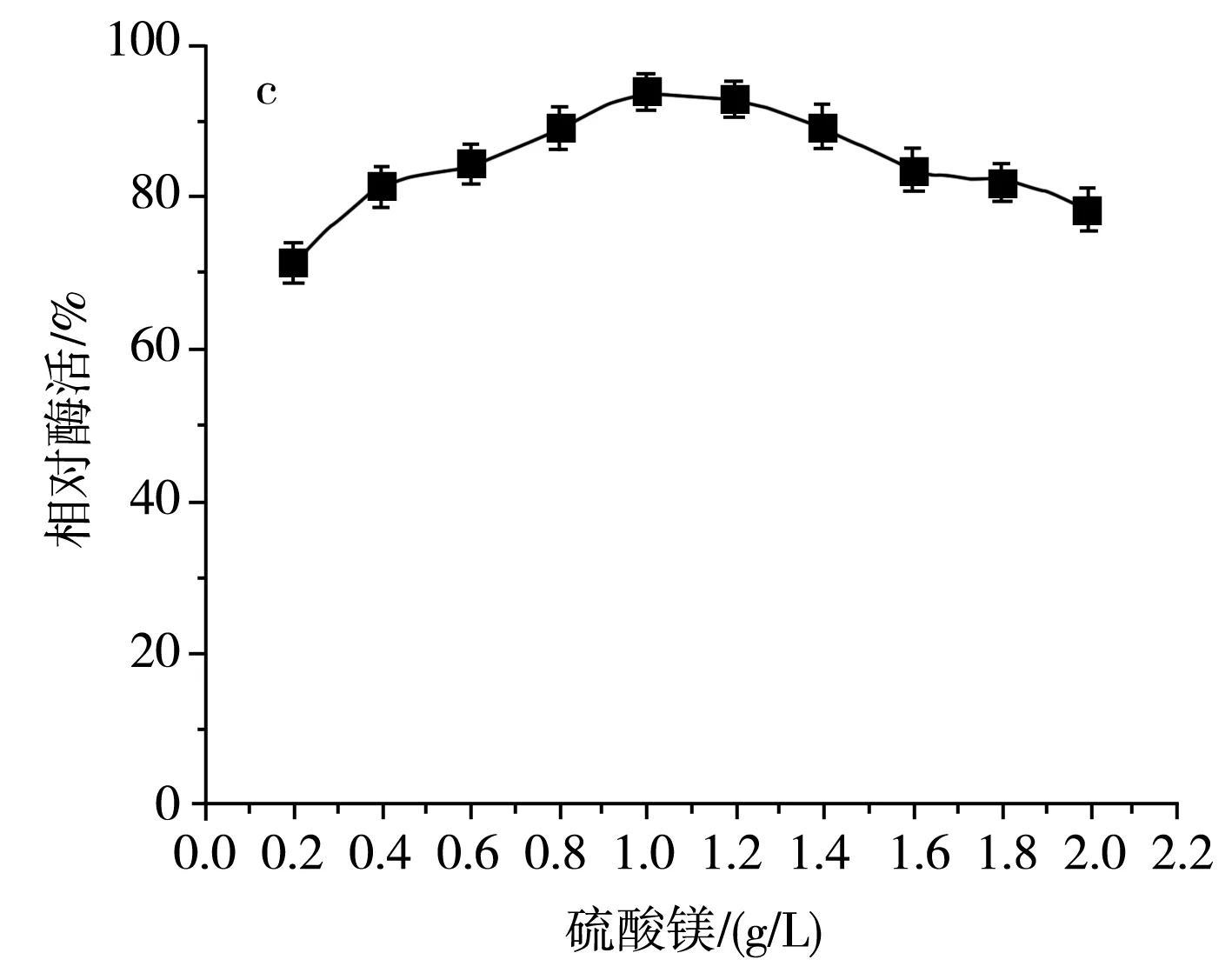
圖7 不同無機鹽及其濃度對SH-50菌株產酶活性的影響Fig.7 Effects of different inorganic salts and their concentrations on enzyme production activity of SH-50 strain
由圖7中a可知,隨著氯化鈉濃度的增加,SH-50菌株產殼聚糖酶活性呈先上升后下降的趨勢,這表明在一定濃度范圍內,氯化鈉的濃度對菌株產酶具有一定激活效應。由圖7中b可知,隨著磷酸氫二鉀濃度的增加,SH-50菌株所產殼聚糖酶的酶活性呈先上升后平穩的趨勢,表明磷酸根離子和鉀離子在一定濃度范圍內可以促進SH-50菌株產酶,但隨著離子濃度的增加,酶活性無明顯變化。由圖7中c可知,硫酸鎂濃度低于1 g/L時對菌株產酶具有一定正激活效應,但隨著濃度的繼續增加,菌株產酶活性逐漸降低。
2.5.4 初始pH及發酵溫度對SH-50菌株產酶活性的影響
該研究分析了發酵液初始pH和溫度對SH-50菌株產酶活性的影響,結果見圖8。
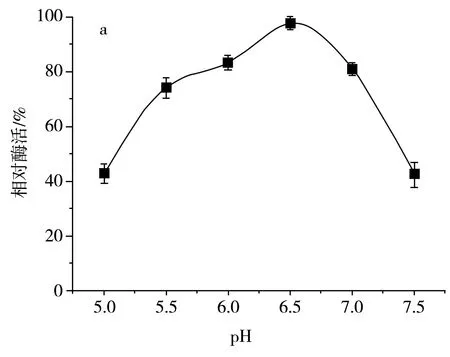

圖8 不同初始pH和溫度對SH-50菌株產酶活性的影響Fig.8 Effects of different initial pH values and temperatures on enzyme production activity of SH-50 strain
由圖8中a可知,初始pH在5.0~6.5范圍內時,SH-50菌株產殼聚糖酶活性隨pH的增大而增大,pH在6.5以上時,SH-50菌株產酶活性逐漸開始降低,故選取pH為6.5作為該菌株初始發酵pH。由圖8中b可知,發酵溫度在25~30 ℃范圍內時,SH-50菌株產殼聚糖酶活性隨溫度升高而增加,在30 ℃以上時,SH-50菌株產酶活性開始顯著下降,故選取30 ℃作為該菌株的發酵溫度。
2.6 響應面優化產酶試驗
2.6.1 PB試驗結果及方差分析
PB試驗結果見表4,試驗回歸及方差分析見表5。
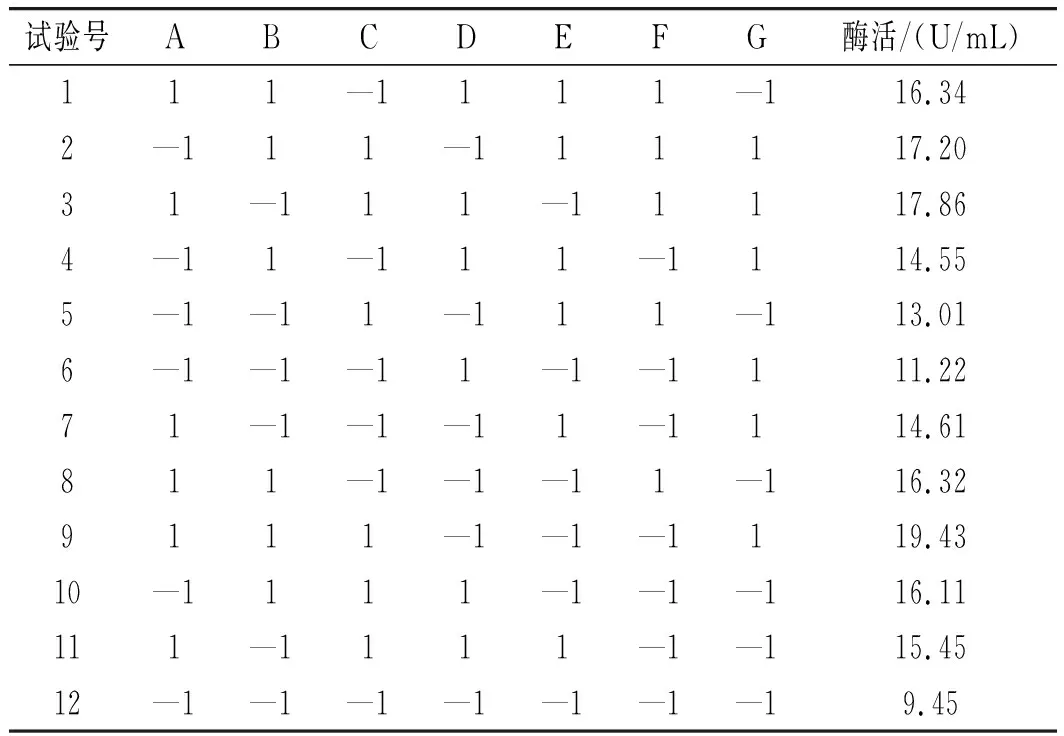
表4 Plackett-Burman試驗設計及結果Table 4 Plackett-Burman experiment design and results

表5 Plackett-Burman試驗回歸及方差分析Table 5 Plackett-Burman test regression and variance analysis
由表5可知,(A)初始pH、(B)發酵溫度、(C)殼聚糖濃度、(G)發酵時間4個因素的P<0.05,表明這4個因素對SH-50菌株產酶活性具有顯著的影響,效應大小依次為A>B>C>G,因此選取A(初始pH)、B(發酵溫度)、C(殼聚糖濃度)3個因素做最陡爬坡試驗。
2.6.2 最陡爬坡試驗
根據3個因素的效應值合理設計原點及步長,結果見表6。

表6 最陡爬坡試驗設計及結果Table 6 Design and results of steepest climbing test

續 表
由表6可知,隨著殼聚糖濃度的增加、初始pH的增加、溫度的降低,SH-50菌株所產酶活性先升高后降低,其中項目4達到最高值,為爬坡試驗的拐點,因此將試驗項目4作為BBD試驗的中心值,進行下一步試驗。
2.6.3 響應面結果及方差分析
在最陡爬坡試驗的基礎上,以(A)初始pH、(B)發酵溫度、(C)殼聚糖濃度作為3個因素,殼聚糖酶活作為響應值設計響應面分析,利用Design Expert V8.0.6軟件對結果進行分析,試驗結果見表7,構建的響應面回歸模型為:Y=24.08+0.36A+2.41B+0.25C-0.67AC-0.88BC-10.01A2-7.02B2-6.44C2。
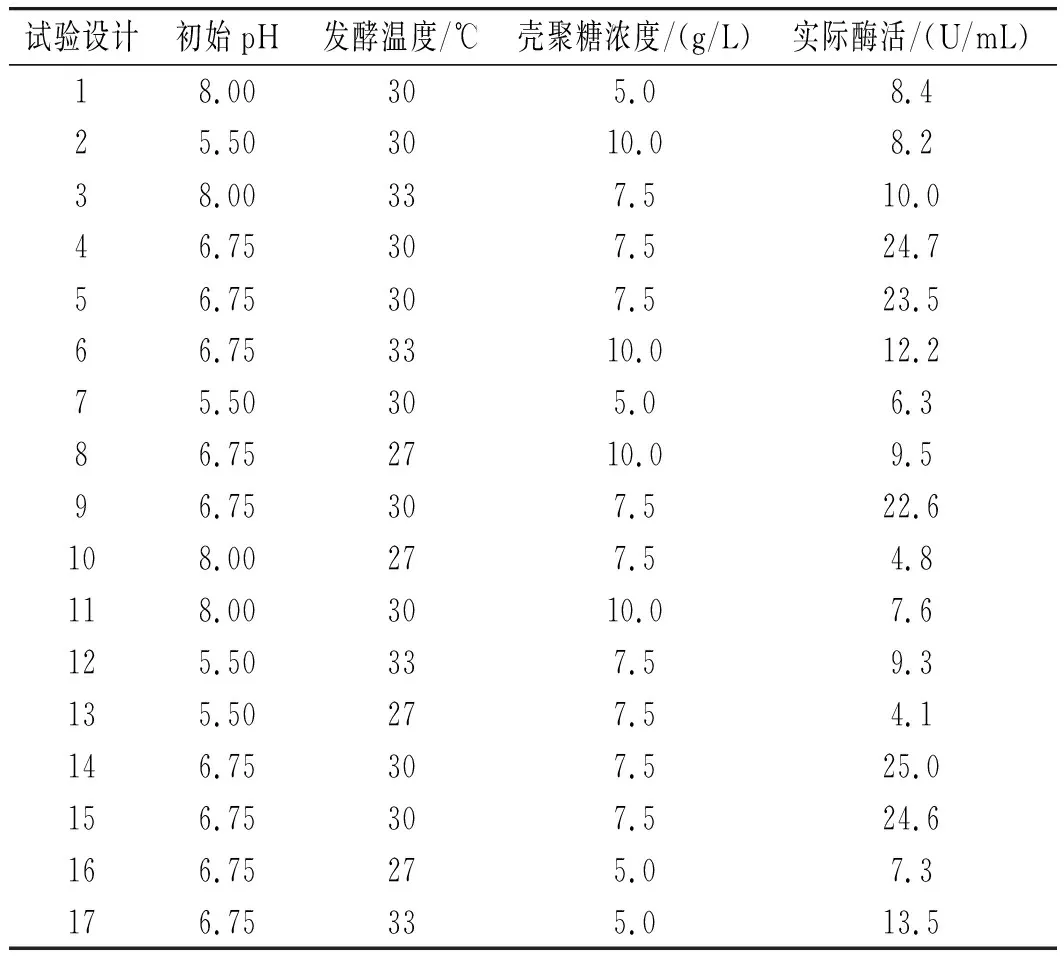
表7 Box-Behnken試驗數據及結果Table 7 Box-Behnken test data and results
分別對模型的方差和顯著性進行統計分析,結果見表8。
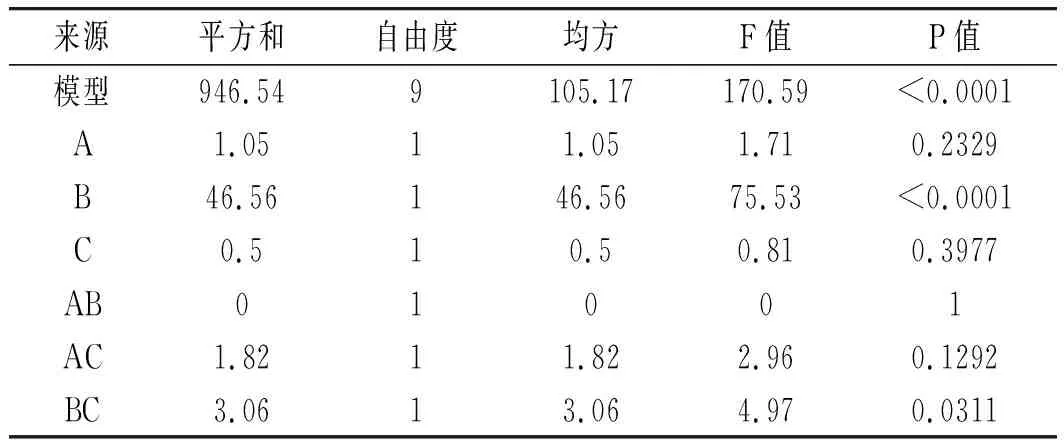
表8 Box-Behnken試驗回歸模型方差分析Table 8 Variance analysis of Box-Behnken test regression model

續 表
回歸模型P<0.0001,表明回歸方程擬合顯著,失擬項的P不顯著,說明模型誤差較小。模型的決定系數R2=0.9955,修正系數RAdj2=0.9896,說明試驗數據變化可以用該模型來解釋且模型顯著性較好。
2.6.4 響應面因素之間的交互作用
運用響應面Design Expert V8.0.6軟件對試驗結果進行擬合,得到影響SH-50菌株產酶活性兩因素之間的等高線圖及三維立體圖,并由此判斷交互影響程度,結果見圖9。
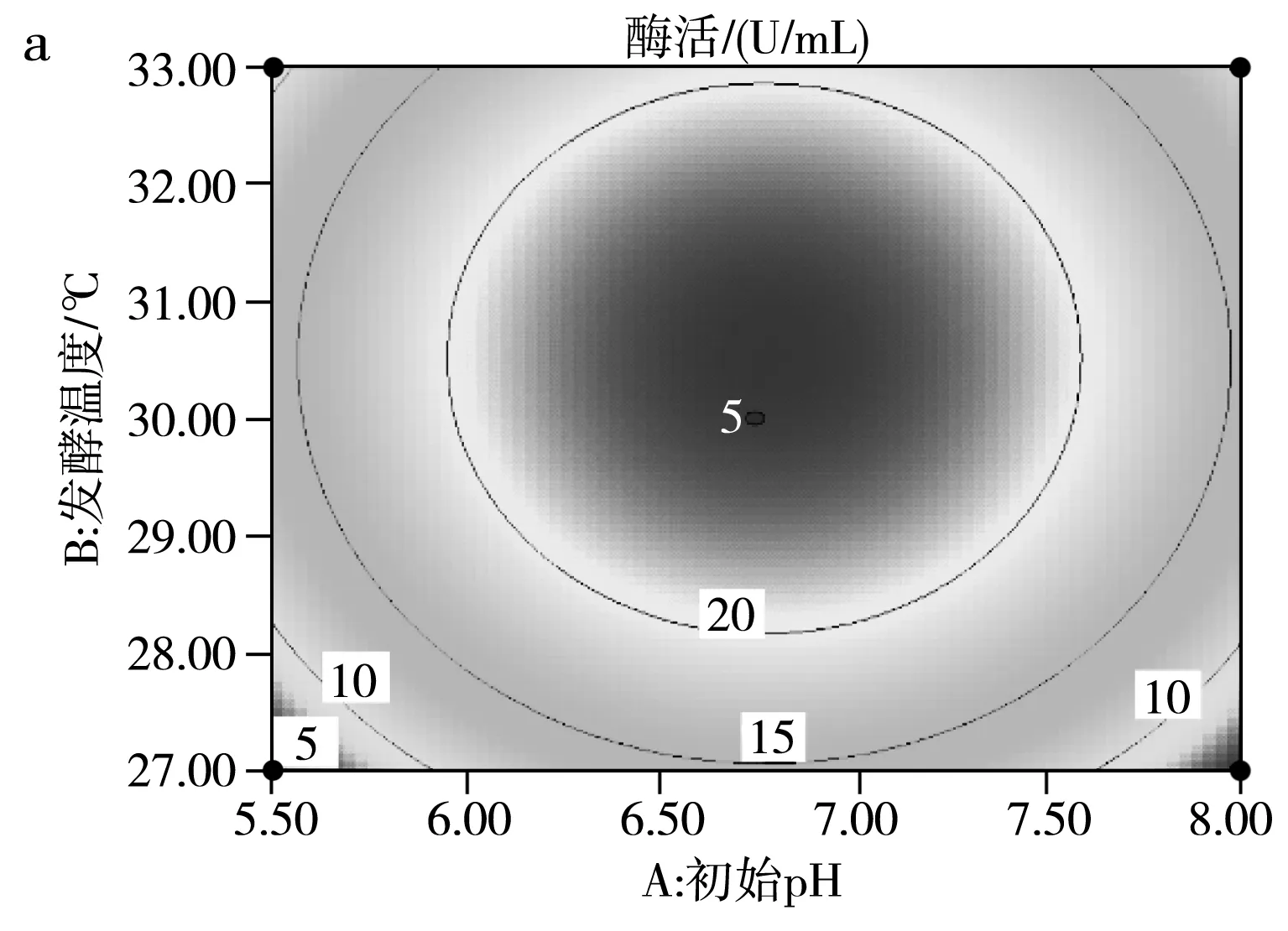



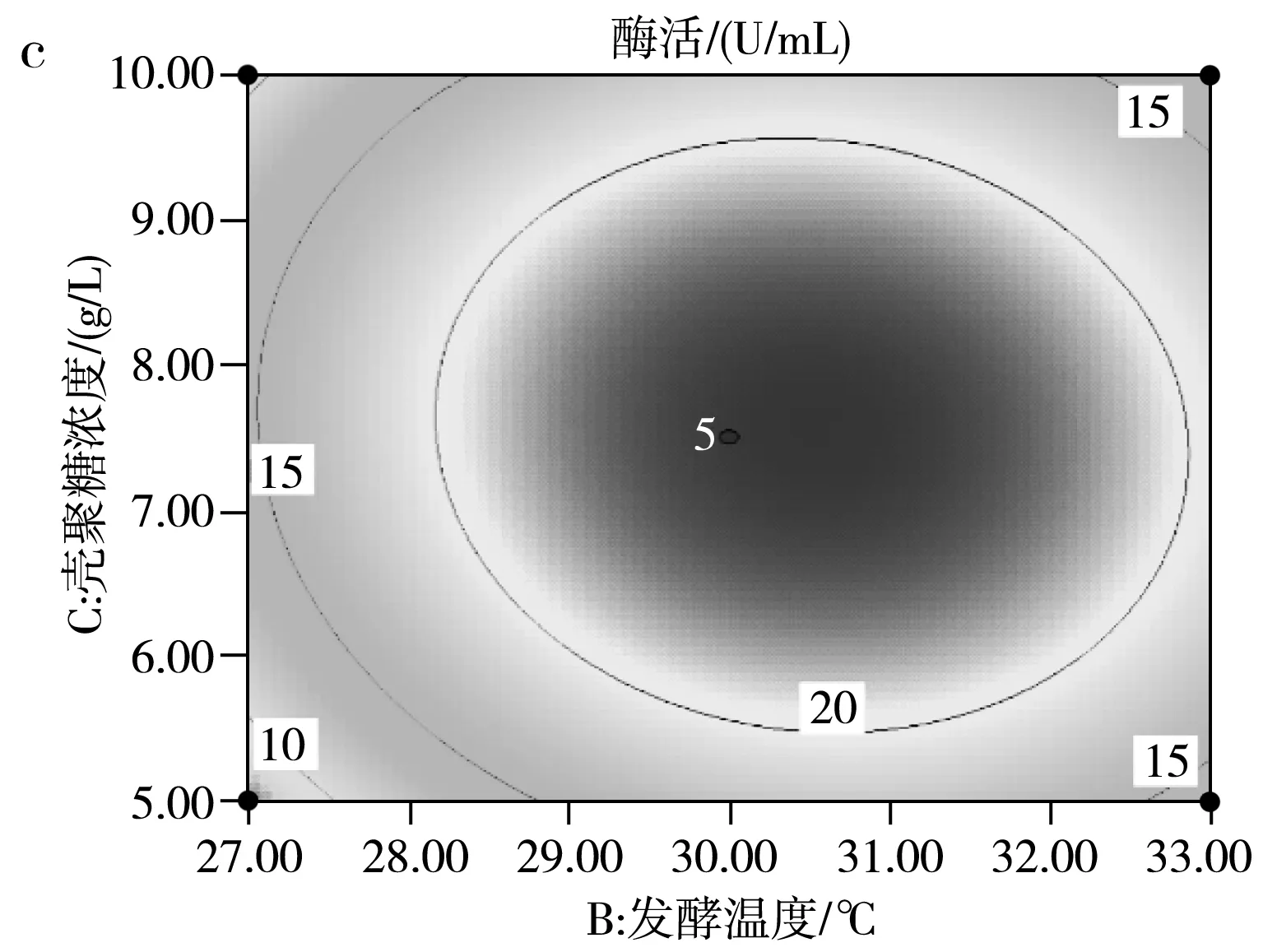

圖9 發酵溫度、初始pH、殼聚糖濃度兩兩因素對SH-50菌株 產酶影響的等高線圖及響應面三維圖Fig.9 Contours and response surface 3D diagrams of the effects of interaction of fermentation temperature, initial pH and chitosan concentration on enzyme production of SH-50 strain
由圖9中a的等高線圖可知,(B)發酵溫度和(A)初始pH兩因素交互作用并不明顯,由圖9中a的三維立體圖可知,隨著發酵溫度和初始pH的增高,SH-50菌株所產殼聚糖酶的酶活先升高后降低,存在中心最大值,中心點即兩因素之間的最適條件。由圖9中b的等高線圖可知,(A)初始pH和(C)殼聚糖濃度兩因素之間交互作用不明顯,同樣存在條件中心點。由圖9中c的等高線圖可知,(B)發酵溫度和(C)殼聚糖濃度兩因素存在明顯交互作用,同樣存在條件中心點。綜上可知,在pH 6.5~7.0、溫度30~31 ℃、殼聚糖濃度7.5~8 g/L的范圍內,存在3個因素間最優產酶條件。
2.6.5 回歸模型的驗證
響應面預測SH-50菌株產酶活性最高的條件為:初始pH 6.77,發酵溫度30.52 ℃,殼聚糖濃度7.52 g/L,預測酶活為24.29 U/mL。實際試驗中設置初始pH 6.75,發酵溫度30.5 ℃,殼聚糖濃度7.52 g/L,重復3次試驗測得平均酶活為26.6 U/mL,與預測值較為接近,說明該模型具有較高可信度。
3 結果與討論
殼聚糖酶的概念是由Shimosaka等在1973年首次提出,于2004年被國際生物化學酶學委員會命名為殼聚糖酶(EC3.2.1.132殼聚糖N-乙酰氨基葡萄糖水解酶)。該研究從海南三亞蝦蟹養殖區土壤中篩選到一株高效產殼聚糖酶的菌株SH-50,結合菌株的生理生化特性及16S rDNA序列,初步鑒定該菌株為松江菌屬,Mitsuariasp.是2005年建立的一個新屬,歸屬β-變形菌門[20],最早在日本松江市被發現。
響應面分析法(RSM)是一種優化生物發酵過程的綜合技術,能夠對微生物發酵影響因素及水平的交互作用進行較好的優化和評價[21],本研究通過響應面法對該菌株產酶條件進行優化,得到的最佳發酵條件為:殼聚糖濃度7.52 g/L,氯化鈉4.0 g/L,硫酸鎂1.0 g/L,磷酸氫二鉀4.0 g/L,溫度30.5 ℃,初始pH 6.75,裝液量100 mL/250 mL,接種量1%,在180 r/min條件下發酵12 h,粗酶液酶活為26.6 U/mL,優化后的產酶活性較初始產酶活性提高了5.3倍。目前Mitsuariasp.菌屬的菌株主要應用在生物防治及生物修復等領域,在殼聚糖酶領域的研究并不多,胡遠亮等[22]對Mitsuariasp. 141-2菌株的突變株進行產酶優化后,得到的殼聚糖酶活性為3.631 U/mL。張馨月等[23]通過響應面法優化了一株產殼聚糖酶的淡紫紫孢菌的發酵條件,得到其發酵5.5 d時的殼聚糖酶活為16.80 U/mL。孫玉英等[24]通過正交試驗優化Cellulophagasp. M5菌株的產酶條件,使其在發酵84 h后產酶活性達到6.67 U/mL。相比其他菌株,該研究中的SH-50菌株產酶活性達到26.6 U/mL,且產酶周期僅為12 h,產酶效率高。值得注意的是,大多數殼聚糖酶屬于誘導酶,所以將殼聚糖作為碳源時需合理考慮其使用濃度,黏度過高易產生大量氣泡從而成為大規模發酵中的限制因素,SH-50菌株產酶培養基中殼聚糖的濃度僅為7.52 g/L,酵母粉僅為1.0 g/L,不僅培養基黏度低,而且所需原料成本低。其次,SH-50菌株最適產酶溫度為30.5 ℃,pH為6.75,發酵條件溫和且易實現。因此,SH-50菌株已經具有工業化的應用能力。
生物酶解法制備低聚殼聚糖的過程既不存在環境風險也不會引入對人體有害的物質,是工業化生產食品級低聚殼聚糖的首選。馮林慧等[25]將殼聚糖作為一種天然防腐劑對食品中容易滋生的李斯特菌、金黃色葡萄球菌、綠膿桿菌、白色假絲酵母及黑曲霉進行抑菌試驗,發現一定濃度的殼聚糖具有良好的抑菌效果,且對綠膿桿菌的抑制效果最為明顯。劉安等[26]使用殼聚糖作為羅漢果及陳皮浸提液的脫色劑,發現低濃度的殼聚糖具有良好的脫色功能,可以顯著提高浸提液的透光率及色度。蘇維發等[27]發現在豬飼料中添加微量的低聚殼聚糖可以提高豬機體的免疫力、抗氧化功能、腸道黏膜形態,進而改善豬肉品質。盡管低聚殼聚糖的聚合度、分子量、水溶性、生物活性均優于殼聚糖[28],但目前仍不能確定其中單一組分的具體功能。新型生物酶制劑的開發研究有望為日后生產特定功能的食品添加劑提供一定的理論依據。
猜你喜歡
河北科技師范學院學報(2022年2期)2022-08-26 08:55:40
河北科技師范學院學報(2021年1期)2021-05-10 03:34:20
中成藥(2017年12期)2018-01-19 02:06:57
電源技術(2017年1期)2017-03-20 13:37:59
廣西科技大學學報(2016年1期)2016-06-22 13:10:38
天然產物研究與開發(2016年1期)2016-06-05 10:29:25
食品界(2016年4期)2016-02-27 07:36:46
中國果菜(2015年2期)2015-03-11 20:01:01
應用化工(2014年7期)2014-08-09 09:20:21
應用技術學報(2014年4期)2014-02-28 14:52:40
