持續風沙流脅迫對黑果枸杞葉片生理特性的影響
2022-07-08 06:05:08魏林源張瑩花侍新萍
草地學報 2022年6期
關鍵詞:植物
黨 緒, 馬 瑞*, 魏林源, 張瑩花, 侍新萍
(1. 甘肅農業大學林學院, 甘肅 蘭州 730070; 2. 甘肅省治沙研究所, 荒漠化與風沙災害防治重點實驗室,甘肅 蘭州 730070)
風沙流是風與其所搬運的固體顆粒組成的氣固二相流。在我國西北地區,由于大風天氣常年出現[1],風沙流活動頻繁,嚴重影響當地植物生存繁衍。一方面,風沙流通過搖曳、擊打、磨蝕對植物造成機械損傷;另一方面間接通過改變周圍氣流,引發植物內部失水,導致其生理特性產生一系列變化[2]。此前,關于風沙流對植物的影響研究涉及風沙流對植物的生長狀況[3-4],光合蒸騰特性[5-6],生物量分配[7],表觀特征[8],解剖結構[9]等方面。而植物受到風沙流脅迫時,會通過形態改變[10-11]和生理適應[12]進行積極反饋,抗氧化酶活性和滲透調節物質發揮重要作用,因此選擇長期生存在風沙環境中的植物,研究其抗氧化酶系統和滲透調節物質含量的變化,是了解植物在發展過程中如何適應風沙環境的重要基礎。
趙丁潔等[1]研究發現8 m·s-1和17 m·s-1風沙流脅迫后,沙拐棗(CalligonummongolicmTurcz.)幼苗過氧化物酶(Peroxidase,POD)活性有顯著差異;趙哈林等[13]發現同一風沙流強度下隨著風吹時間的增加,樟子松(Pinussylvestnisvar.mongolica)幼苗超氧化物歧化酶(Superoxide dismutase,SOD)活性下降,丙二醛(malonaldehyde,MDA)含量、膜透性、POD活性、可溶性糖(soluble sugar,SS)和脯氨酸(proline,Pro)含量趨于增加;周瑞蓮等[2]研究表明,隨風沙流風速提高至15 m·s-1,黑松(PinusthunbergiiParl)在短時低風速吹襲時葉片平均MDA含量、細胞膜透性、SOD、過氧化氫酶(Catalase,CAT)、POD活力較對照均有所升高;李道明等[14]研究發現不同強度風沙流脅迫下沙拐棗幼苗CAT活性均下降,POD活性均降低但大于對照;輕度風沙流脅迫下沙拐棗幼苗PSⅡ最大光化學效率(Fv/Fm)、PSⅡ最大效率(Fv′/Fm′)、PSⅡ運行效率(Fq′/Fm′)、非光化學淬滅系數NPQ變化均不明顯;重度風沙流脅迫下,Fv/Fm、Fv′/Fm′、Fq′/Fm′和光化學淬滅系數qp均下降,NPQ則升高。
風沙流脅迫對植物抗氧化酶、滲透調節物質和熒光參數等的影響會因脅迫強度、脅迫時間和植物種類而異。黑果枸杞(Lyciumruthenicum)是茄科(Solanceae)枸杞屬(Lycium)多棘刺灌木,適應性強,耐高溫、耐寒耐旱,喜生于鹽堿荒地,有防風固沙作用,可作為干旱、半干旱地區植被恢復重建的植物種[15-16],野生黑果枸杞植株矮小,其生育期均處于風沙流侵襲高度范圍內。目前,關于黑果枸杞在持續風沙流脅迫后生理特性如何變化研究較少。本試驗以民勤和永靖種源黑果枸杞盆栽苗為研究對象,通過風洞試驗模擬持續風沙流脅迫,記錄在9 m·s-1風沙流脅迫過程中葉片形態變化,分析其經歷脅迫后膜系統、抗氧化酶系統、滲透調節物質和熒光參數的變化規律,以期揭示民勤、永靖黑果枸杞在9 m·s-1風沙流持續脅迫后的生理響應機制,研究結果拓寬了植物逆境生理生態學理論,并為風沙地區黑果枸杞種植選育提供一定理論依據。
1 材料與方法
1.1 試驗材料
試驗材料為3齡黑果枸杞盆栽實生苗,種子采自民勤青土湖區(103°02′~104°02′ E,38°05′~39°06′ N)和永靖縣鹽鍋峽鎮(102°53′~103°39′ E,35°47′~36°12′ N)野生黑果枸杞。2018年用采集的種子在育苗缽中育苗,待其長至10 cm后帶土移栽至塑料盆中,每盆1株,置于甘肅省武威市治沙研究所綠洲站的苗圃內(102°35′ E,37°56′ N)培育,該地年平均氣溫7.8℃,降水量60~610 mm,蒸發量1 400~3 010 mm,日照時數2 200~3 030 h,無霜期85~165 d,是典型的大陸性氣候。
2020年8月22日,選取長勢良好且生物學特性無顯著差異的民勤、永靖黑果枸杞作為供試材料,移至風洞實驗室外通風、半蔭蔽樹冠下,澆等量水緩苗。
1.2 試驗設計
風沙流模擬試驗在甘肅省武威市治沙研究所的風沙環境風洞實驗室完成。該風洞為直流下吹式,軸線風速在4~30 m·s-1連續可調,試驗段長16 m。依據民勤和永靖季節性風速范圍[17-18],以往風吹研究報道中不同強度[2,19-20]、不同歷時[13]風沙流對植物葉片的損害程度,以9 m·s-1風速對應的風沙流(輸沙量為28.30 g·cm-1·min-1)進行脅迫。
2020年8月24日開始脅迫試驗:在試驗段鋪滿10~12 cm厚度的沙子,平整沙面后,選取4盆民勤供試黑果枸杞放于風洞出風口,調整植株高度,使其冠部處于風沙流吹襲范圍內進行脅迫,脅迫時間為1 h。另做2個重復,每個重復為4盆供試枸杞。永靖種源脅迫處理與民勤相同,試驗持續4 d。
每次脅迫結束后將植株搬至緩苗區,并記錄葉片變化情況。第4 d脅迫結束后將植株搬至緩苗區靜置30 min后快速隨機剪取冠部上、中、下葉片,一部分帶回實驗室立即進行膜透性和熒光參數測定,另一部分用錫紙包裹經液氮移至超低溫冰箱保存,用于其余生理生化指標測定。植株于風洞實驗室外通風、半蔭蔽樹冠下進行恢復生長,第11 d再取樣一次。試驗前隨機采集供試植株冠部上、中、下各部位葉片作為對照(CK)。
1.3 測定指標及方法
膜透性采用電導率法[19]測定;MDA含量采用硫代巴比妥酸法[19]測定;POD活性采用愈創木酚法[19]測定;SOD活性采用氮藍四唑染色法[21]測定;CAT活性采用紫外吸收法[22]測定;可溶性蛋白(soluble protein,SP)含量采用考馬斯亮藍G-250法[21]測定;Pro含量采取酸性茚三酮法[21]測定;采用葉綠素熒光圖像分析系統(CF imager,UK)測定葉片的葉綠素熒光參數,各參數計算將由內置軟件自動記錄并控制運行,具體測定程序參照師生波等[23]的方法。測定時,每個指標均做6個重復。
1.4 數據處理
運用SPSS 19.0統計分析軟件對實驗數據進行分析。采用單因素方差分析(One-Way ANOVA),Duncan法對同一種源不同取樣時間內的差異性進行分析(P<0.05);采用雙因素方差分析(Two-way ANOVA)檢驗種源和脅迫后不同時間及其交互作用對各生理指標的影響。用Microsoft Excel 2010制作圖表。
2 結果與分析
2.1 風沙流脅迫下黑果枸杞葉片形態受損情況
風沙流脅迫第1 d,永靖黑果枸杞較民勤落葉更多,民勤有極少量嫩葉出現褐色斑塊;風沙流脅迫第2 d,民勤葉片多數出現褐色斑塊、彎曲,永靖只有少數葉片出現褐色斑塊;風沙流脅迫第3 d,民勤有褐色斑塊葉片數量增多、彎曲、少量嫩莖斷裂,永靖有褐色斑塊葉片數量增多,二者葉片均有萎蔫;風沙流脅迫第4 d,民勤葉片整體萎蔫、部分葉片發黑脫落,永靖有褐色斑塊的葉片數量增多;第11 d,兩種源葉片萎蔫(表1)。脅迫過程中兩種源黑果枸杞均有落葉,第3 d脅迫后葉片出現積沙。可見,風沙流導致黑果枸杞葉片受損,出現褐色斑塊,萎蔫。

表1 風沙流脅迫對黑果枸杞葉片形態的影響Table 1 Effects of wind-sand flow stress on the leaf morphology of Lycium ruthenicum
2.2 風沙流脅迫下黑果枸杞葉片膜透性和MDA含量變化
民勤和永靖葉片MDA含量在第4 d分別較CK顯著(P<0.05)增加24.6%,13.0%;民勤葉片在第11 d較CK顯著(P<0.05)下降了24.0%,而永靖葉片則較CK顯著(P<0.05)增加了13.9%(圖1A)。民勤、永靖葉片膜透性在第4 d與CK無顯著差異,均在第11 d最大,分別較CK顯著(P<0.05)增加了56.05%,25.91%(圖1B)。可見,持續4 d風沙流脅迫導致永靖葉片MDA含量持續積累,而民勤葉片MDA僅在第4 d增加;脅迫后在第11 d,黑果枸杞葉片細胞膜透性增大。
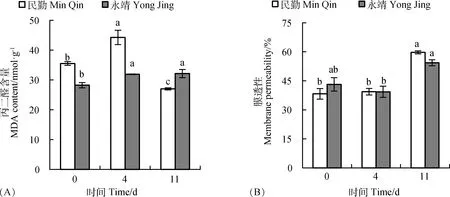
圖1 風沙流脅迫對黑果枸杞葉片MDA含量和膜透性的影響Fig.1 Effects of wind-sand flow stress on the MDA content and membrane permeability in the Leaves of Lycium ruthenicum注:不同小寫字母表示同一種源不同時間之間差異顯著(P<0.05),下同Note:Values with different lowercase letters indicate significant differences between different times in the same seed source at the 0.05 level,the same as below
2.3 風沙流脅迫下黑果枸杞葉片抗氧化酶活性變化
民勤、永靖葉片CAT活性均先增后減,在第4 d有最大值,分別較CK顯著(P<0.05)增加了104.04%,44.38%(圖2A)。民勤葉片POD活性先減后增,在第4 d有最小值,較CK顯著(P<0.05)下降了33.08%,永靖POD活性先增后減,在第4 d有最大值,為CK的3.01倍(圖2B);民勤葉片SOD活性先減后增,在第11 d較CK顯著(P<0.05)增加,而永靖SOD活性呈下降趨勢,與CK差異不顯著(P<0.05)(圖2C)。民勤葉片主要通過提高CAT、SOD活性,永靖葉片主要通過提高CAT、POD活性來減輕持續風沙流脅迫對其細胞造成的損傷。
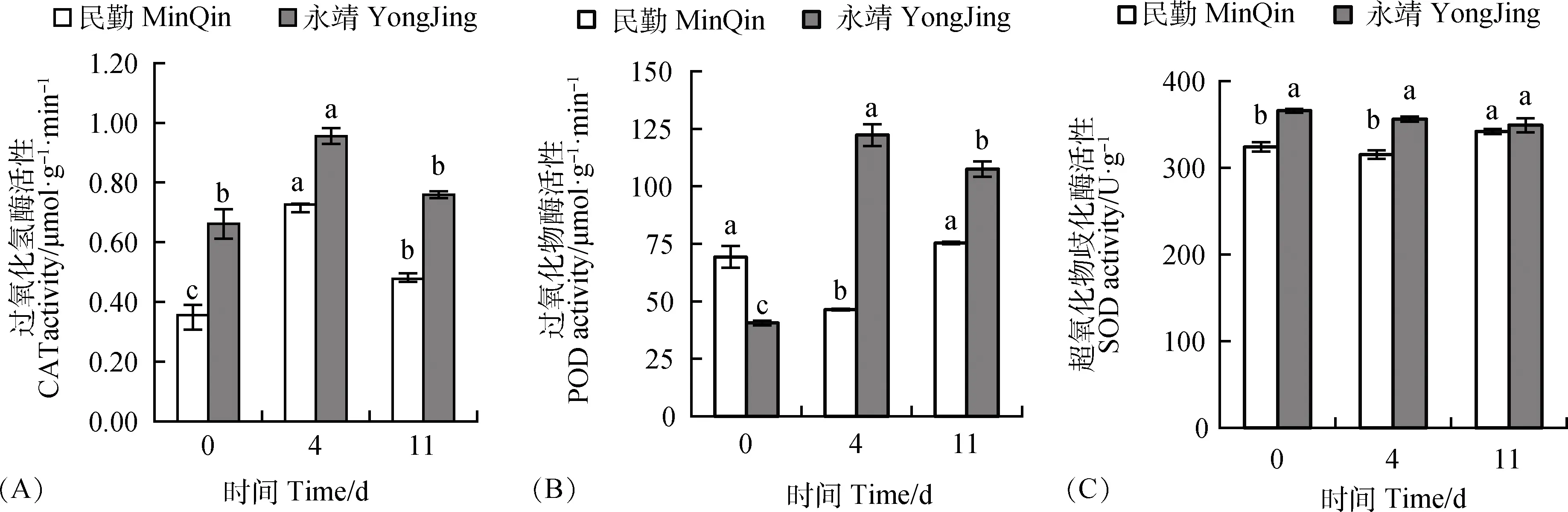
圖2 風沙流脅迫對黑果枸杞葉片抗氧化酶活性的影響Fig.2 Effects of wind-sand flow stress on the protective enzyme activity in the leaves of Lycium ruthenicum
2.4 風沙流脅迫下滲透調節物質含量變化
民勤葉片Pro含量逐漸積累,在第11 d有最大值,較CK顯著(P<0.05)增加了149.82%,永靖葉片Pro含量先降后增,但均小于CK含量(圖3A)。民勤、永靖葉片SP含量均逐漸減少,在第11 d時有最小值,分別較CK顯著(P<0.05)降低了11.98%,19.97%(圖3B)。風沙流脅迫后,民勤葉片通過增加Pro含量來減輕其所受傷害,永靖葉片滲透調節作用有限。
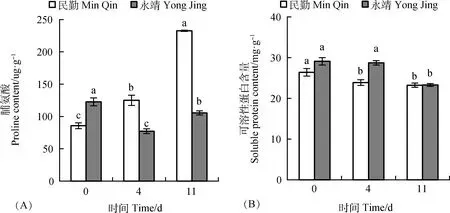
圖3 風沙流脅迫對黑果枸杞葉片滲透調節物質含量的影響Fig.3 Effects of wind-sand flow stress on osmotic regulation substances in the leaves of Lycium ruthenicum
2.5 風沙流脅迫下葉片葉綠素熒光參數的變化
由圖4A可知,民勤葉片Fv/Fm在受到風沙流脅迫后逐漸下降,但與CK差異不顯著(P<0.05),永靖葉片Fv/Fm先減后增,在第4 d較CK顯著(P<0.05)下降了7.8%,第11 d與CK無顯著(P<0.05)差異。
在低穩態光(400 μmol·m-2·s-1)照射下,民勤葉片PSⅡ最大效率(Fv′/Fm′),PSⅡ運行效率(Fq′/Fm′)和光化學猝滅系數qp均呈下降趨勢,在第11 d有最小值,且在第11 d與CK存在顯著(P<0.05)差異,非光化學猝滅系數NPQ先減后增,但均與CK差異不顯著(P<0.05);永靖葉片Fv′/Fm′先減后增,在第11 d較CK顯著(P<0.05)增加了6.40%,Fq′/Fm′、qp呈下降趨勢,第11 d有最小值,NPQ先增后減,但均與CK差異不顯著(P<0.05)(圖4B、圖4C、圖4D、圖4E)。在高穩態光(1 500 μmol·m-2·s-1)照射下,民勤Fv′/Fm′、Fq′/Fm′、qp先減后增,Fv′/Fm′、Fq′/Fm′在第4 d較CK顯著(P<0.05)下降,第11 d與CK無顯著(P<0.05)差異,NPQ先增后減,但均與CK無顯著(P<0.05)差異;永靖Fv′/Fm′、NPQ先增后減,在第4 d有最大值,Fq′/Fm′呈增加趨勢、qp先減后增,均在第4 d與CK差異不顯著(P<0.05),在第11 d達到最大值(圖4F、圖4G、圖4H、圖4I)。
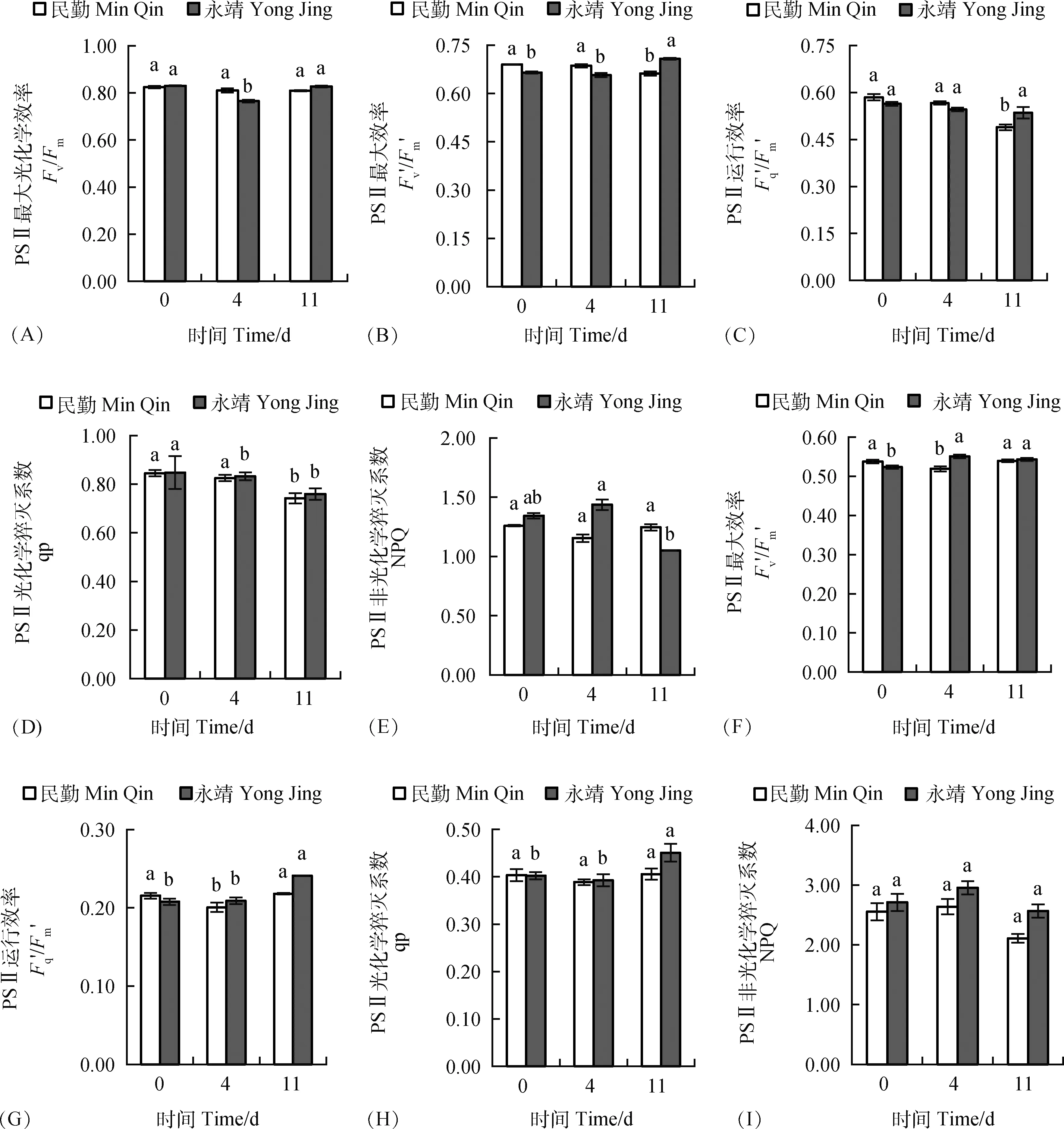
圖4 風沙流脅迫對黑果枸杞葉綠素熒光參數的影響Fig.4 Effects of wind-sand flow stress on Chlorophyll fluorescence parameters in the leaves of Lycium ruthenicum
2.6 種源和時間對黑果枸杞葉片各生理指標的雙因素方差分析
由表2可知,種源對黑果枸杞葉片MDA含量,CAT、POD和SOD活性,Pro和SP含量,1 500 μmol·m-2·s-1光強下Fv′/Fm′、Fq′/Fm′有顯著(P<0.05)影響;脅迫后時間對黑果枸杞葉片MDA含量、膜透性、CAT和POD活性,Pro和SP含量、Fv/Fm、400 μmol·m-2·s-1和1 500 μmol·m-2·s-1光強下Fv′/Fm′、Fq′/Fm′、qp有顯著(P<0.05)影響;種源和時間的交互作用對MDA含量、POD和SOD活性,Pro和SP含量、Fv/Fm、400 μmol·m-2·s-1光強下Fv′/Fm′、Fq′/Fm′、NPQ及1 500 μmol·m-2·s-1光強下Fv′/Fm′、Fq′/Fm′均有顯著(P<0.05)影響。
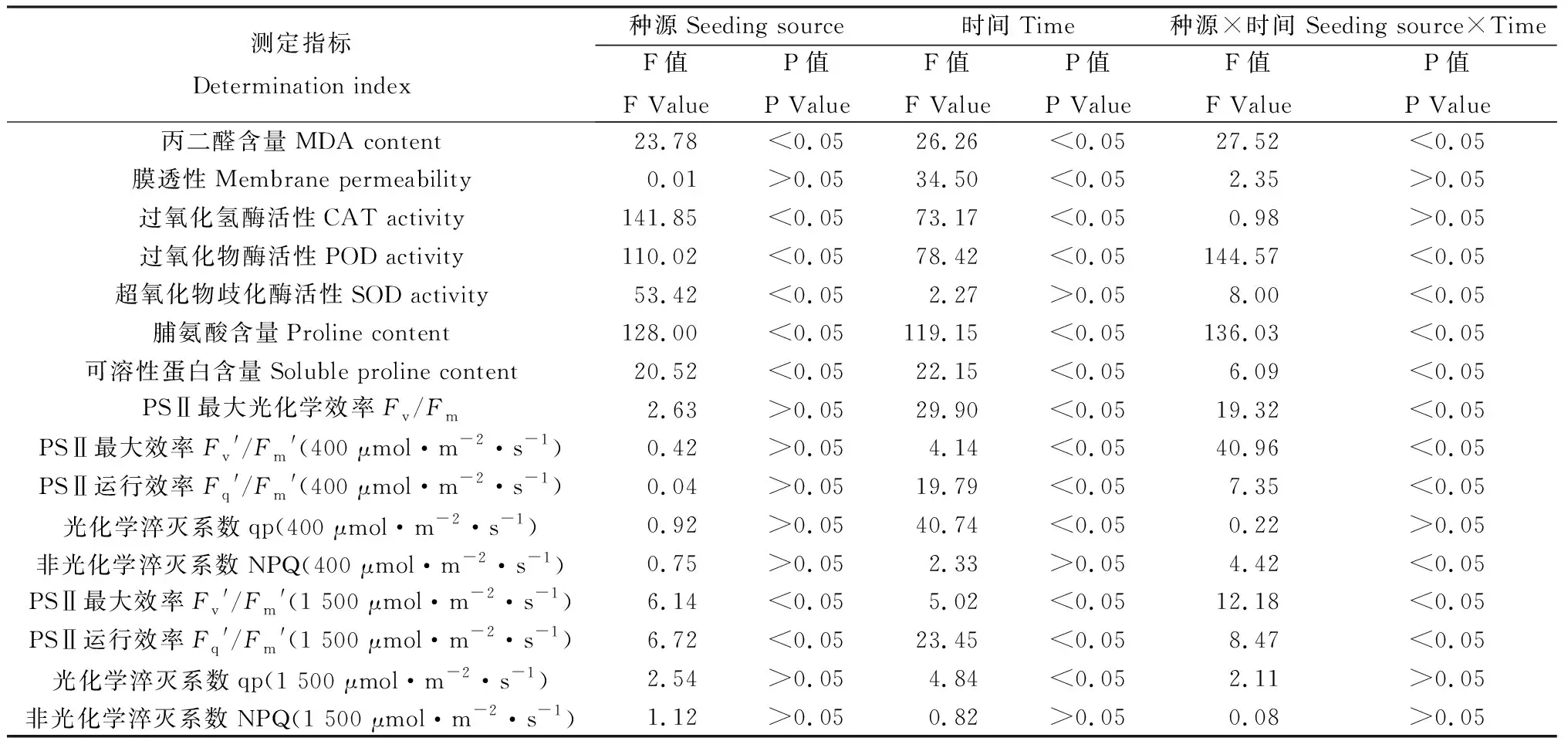
表2 種源和時間對黑果枸杞葉片各生理指標的雙因素方差分析Table 2 Double factor variance analysis of seeding sources and times on physiological indexes of Lycium ruthenicum leaves
3 討論
植物受到逆境脅迫時,體內自由基產生與清除的平衡被打破,過量的自由基積累破壞細胞[24],導致膜脂發生過氧化反應,其產物MDA積累[25];發生脫脂化反應,膜透性增大[21],因而MDA含量和細胞膜透性是衡量質膜受傷害程度的重要指標。有研究表明,風沙流脅迫下霸王(Zygophyllumxanthoxylum)葉片[26]和沙蒿(Artemisiasphaerocephala)葉片[27]MDA含量并未顯著增加,本研究結果與之不同,在第4 d風沙流脅迫后,兩種源葉片MDA含量均顯著增加,發生明顯膜脂過氧化反應,這主要是因為在霸王和沙蒿的研究中風沙流脅迫時間分別為20 min和30 min且均為單次脅迫,而本試驗脅迫時間為1 h且持續4 d,導致自由基大量積累,MDA含量顯著增加。第11 d,民勤葉片MDA含量顯著低于第4 d含量,而永靖葉片MDA含量高于第4 d含量,這可能是因為在第11 d民勤葉片CAT、SOD活性及Pro含量顯著增高,協調清除產生的自由基,因此MDA含量顯著降低,而永靖葉片只有POD活性顯著升高,清除作用有限。第11 d兩種源葉片膜透性均增大,風沙流脅迫過程中沙粒擊打、磨蝕黑果枸杞產生的作用在其此后生長過程中逐漸表現出來,其傷害具有一定程度的滯后性。植物受逆境脅迫時會積極啟動抗氧化酶系統,提高CAT、SOD、POD活性,清除活性氧,以保護植物細胞膜[28],同時,通過滲透調節積累溶質,提高Pro、SP等的含量維持細胞膨壓或滲透平衡、提高細胞液濃度等保護植物[29-30]。楊永義等[20]研究發現,20 min風和風沙流脅迫會提高民勤黑果枸杞葉片POD活性,本試驗中民勤葉片POD活性在第4 d顯著下降,這可能是因為本試驗中風沙流持續時間較長,脅迫后POD活性受到了抑制,這與張榮梅等[31]和姜霞等[32]對黑果枸杞耐鹽性的研究中POD活性變化類似。在第4 d,永靖Pro含量明顯下降,可能是因為持續風沙流脅迫導致其細胞嚴重失水,結構和功能受到損傷,Pro合成能力受到抑制[33],且在第11 d尚未完全恢復。SP含量減少,可能是因為脅迫后植物蛋白質合成受阻,加速分解成各種氨基酸通過降低葉片水勢[34]來減輕黑果枸杞的受害程度。
葉綠素熒光反應植物光能吸收,激發能傳遞和光化學反應,作為光合作用的探針,幾乎可以反映所有光合作用過程[35-36]。Fv/Fm作為衡量植物葉片中PSⅡ原初光能轉換效率高低的重要依據,在沒有受到脅迫時該參數很穩定(0.80~0.85)[37],永靖葉片Fv/Fm在第4 d風沙流脅迫后顯著下降,第11 d沒有風沙流脅迫時恢復至正常水平,這說明風沙流脅迫后永靖黑果枸杞受到了光抑制而非光破壞,風沙流對其葉片PSⅡ反應中心造成的損傷可恢復。Fv′/Fm′反映開放的PSⅡ反應中心原初光能捕獲效率[37];PSⅡ運行效率(Fq′/Fm′)是實際電子傳遞的量子效率,反映電子在PSⅠ和PSⅡ的傳遞情況[38];qp表示PSⅡ天線色素吸收的光能用于光化學電子傳遞的份額,一定程度上可衡量PSⅡ反應中心的開放比例[39];NPQ則表示PSⅡ所吸收的光能不能用于光合電子傳遞,而是以熱能耗散,對植物光合機構免受傷害起到了積極作用[40-41]。第4 d,低光強(400 μmol·m-2·s-1)下民勤、永靖葉片Fv′/Fm′、Fq′/Fm′及高光強下民勤Fv′/Fm′、Fq′/Fm′均下降,這說明風沙流脅迫導致黑果枸杞葉片PSⅡ反應中心原初光能捕獲效率降低、PSⅡ實際傳遞的量子效率降低,從而導致從天線上捕獲的光能用于光化學的份額減少,PSⅡ實際量子產量反應中心的光化學活性減弱[42]。第11 d高光強(1 500 μmol·m-2·s-1)下民勤、永靖葉片Fv′/Fm′、Fq′/Fm′均增加,這說明脅迫后恢復過程中高穩態光強下PSII反應中心原初光能捕獲效率增大,從而擁有較多的能量捕獲陷阱;光能電子傳遞的量子效率增大,有利于維持植物的光能轉化效率,同時為暗反應的碳同化積累更多的同化力。1 500 μmol·m-2·s-1光強下民勤、永靖葉片qp均先減后增,NPQ均先增后減,存在一定互補關系,即第4 d風沙流脅迫后,高光強下民勤、永靖黑果枸杞葉片吸收的光能以熱量散失的能量增多,這與李德祿等[43]研究中水分脅迫后云杉屬兩種植物的qp和NPQ的變化類似;第11 d葉片吸收的光能則多用于電子傳遞,減少了以熱能消耗的比例。
4 結論
9 m·s-1持續風沙流脅迫后,民勤黑果枸杞葉片主要通過提高CAT、SOD活性,增加Pro含量,永靖葉片主要通過提高CAT、POD活性以減輕風沙流對其造成的傷害。研究結果可為風沙地區黑果枸杞種植選育提供一定的理論依據。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
