馬鈴薯StPEBP基因家族成員鑒定及其在塊莖中的表達分析
2022-07-05 00:12:06張文哲薦紅舉尚麗娜張梅花王季春AhmedMahmoud呂典秋
中國馬鈴薯 2022年2期
關鍵詞:分析
張文哲,薦紅舉,2,尚麗娜,張梅花,王季春,2,Ahmed Mahmoud,呂典秋,2*
(1.西南大學農學與生物科技學院,重慶 400715;2.薯類生物學與遺傳育種重慶市重點實驗室,重慶 400715;3.Department of Vegetable Crops,Faculty of Agriculture,Cairo University,Giza 12613,Egypt)
馬鈴薯是全球第四大糧食作物,種植面積僅次于小麥、水稻和玉米,在食品和工業(yè)中具有重要用途[1]。馬鈴薯作為主糧,其塊莖中富含淀粉、維生素、礦質元素和纖維素,且脂肪類物質少,提高馬鈴薯產(chǎn)量和質量可保障國家糧食安全。但馬鈴薯產(chǎn)量和品質易受環(huán)境因素影響,因此,鑒定馬鈴薯塊莖形成相關基因對于解析馬鈴薯塊莖形成機制進而提高馬鈴薯產(chǎn)量和品質至關重要。
磷脂酰乙醇胺結合蛋白(Phosphatidylethanolaminebinding protein,PEBP)基因家族廣泛存在于細菌、酵母、真菌和動植物中。在植物中,PEBP 基因家族根據(jù)功能主要分為3個亞家族:FT(FLOWERING LOCUS T)、TFL1(TERMINAL FLOWER 1)和MFT(MOTHER OF FT AND TFL1),MFT 亞家族是FT亞家族和TFL1 亞家族的祖先,最先被挖掘出,隨后鑒定出具有功能的FT 和TFL1 亞家族[2]。MFTlike 亞家族主要在種子的萌發(fā)中發(fā)揮重要作用[3]。在擬南芥中,F(xiàn)T-like 亞家族和TFL1-like 亞家族具有高度的氨基酸序列同源性,他們都在開花過程中發(fā)揮調節(jié)作用[2],F(xiàn)T-like 亞家族通過調節(jié)溫度和光周期促進花的形成并控制生殖生長階段的開始,而TFL1-like 亞家族則抑制花的形成[4]。同時,TFL1-like 亞家族通過維持頂端分生組織和花序分生組織的不確定性來控制營養(yǎng)生長和生殖生長階段的持續(xù)時間[2,5]。馬鈴薯塊莖形成與擬南芥開花過程共享相同或相似的調節(jié)分子和通路[6-8]。在擬南芥中,F(xiàn)T 蛋白作為成花素,可以從葉片運輸?shù)巾敹朔稚M織,并促進花器官的形成[9-11],在植物成花誘導過程中發(fā)揮重要調控作用。馬鈴薯StPEBP家族基因中SELF-PRUNING 6A(StSP6A)和StSP5G已被鑒定為擬南芥FT 蛋白的同源基因,并參與馬鈴薯塊莖的形成過程[12]。隨著馬鈴薯中FT 蛋白功能的深入研究,發(fā)現(xiàn)馬鈴薯開花調節(jié)機制和短日照條件促進馬鈴薯塊莖形成機制是由2種不同的FT類型同源基因(StSP5G和StSP6A)控制[6,13]。StSP5G和StSP6A作為塊莖形成的信號,其在馬鈴薯塊莖形成過程中發(fā)揮重要作用,StSP5G是塊莖形成的負調節(jié)因子,而StSP6A是塊莖形成的正調節(jié)因子[14-16]。
已在多個物種中鑒定出PEBP 基因家族成員,如擬南芥、陸地棉、甘藍型油菜等[17-21],目前馬鈴薯中StPEBP基因家族成員雖被鑒定[22],但該家族基因在結薯中的功能分析尚未深入研究。本研究將擬南芥PEBP 基因家族的蛋白序列與馬鈴薯全基因組的蛋白序列通過Blastp 方式以及保守結構域鑒定,獲得馬鈴薯StPEBP基因家族基因,并對其motif 序列、基因結構以及啟動子元件進行預測和分析,深入挖掘除StSP5G和StSP6A外在馬鈴薯塊莖誘導中發(fā)揮功能的基因,為馬鈴薯結薯基因的篩選提供理論參考。
1 材料與方法
1.1 序列的獲得
從Ensembl Plants 數(shù)據(jù)庫(https:∕∕plants.ensembl.org∕)中獲取擬南芥以及馬鈴薯的全基因組蛋白序列,在擬南芥全基因組蛋白序列中篩選出PEBP基因家族的蛋白序列。將擬南芥PEBP 基因家族的蛋白序列與馬鈴薯的全基因組蛋白序列在NCBI數(shù) 據(jù) 庫(https:∕∕www.ncbi.nlm.nih.gov∕)中 進 行Blastp,閾值E<10-5,初步篩選擬南芥PEBP 基因家族在馬鈴薯中的同源基因。將初步鑒定的基因輸入Pfam 數(shù)據(jù)庫(https:∕∕pfam.xfam.org∕)再次進行篩選(Pfam 號:PF01161),最終確定馬鈴薯中StPEBP基因家族的基因,同時提取馬鈴薯中StPEBP基因家族的蛋白序列和CDS序列。
1.2 序列比對及進化分析
利用MEGA 7.0 軟件對獲取的擬南芥、馬鈴薯、番茄、煙草和辣椒基因組內的PEBP 基因家族的序列進行多重序列比對,番茄、煙草和辣椒的PEBP 基因家族的序列均是比對擬南芥PEBP 基因家族的蛋白序列鑒定得出。隨后使用鄰接法(Neighbor joining,NJ)構建基因分子進化樹,參數(shù)均為默認,并利用iTOL(https:∕∕itol.embl.de∕)對進化樹進行優(yōu)化。
1.3 馬鈴薯StPEBP 基因結構和motif預測
根據(jù)獲得的馬鈴薯StPEBP基因家族的基因組DNA 和CDS 序 列,使 用 線 上 工 具GSDS(http:∕∕gsds.cbi.pku.edu.cn∕)對其基因結構進行分析。利用 在 線 工 具MEME(http:∕∕meme-suite.org∕)對 馬鈴薯StPEBP基因家族蛋白序列的motif 進行預測,motif 數(shù)量設置為10 個,其余參數(shù)為默認。利用TBtools v1.09 軟件對基因結構和motif 分析并作圖。
1.4 順式作用元件預測
使用TBtools 軟件提取馬鈴薯StPEBP基因的CDS 序列上游2 000 bp 的啟動子序列,提交到PlantCare 網(wǎng) 站(http:∕∕bioinformatics.psb.ugent.be∕webtools∕plantcare∕html∕)進 行 順 式 作 用 元 件 預測。將預測的順式作用元件位點、類型以及長度放入TBtools v1.09進行可視化。
1.5 染色體定位與共線性分析
在馬鈴薯全基因組注釋文件中獲得StPEBP家族基因的起始位點和終止位點,使用TBtools v1.09 軟件展示StPEBP家族基因在染色體上的位置。使用TBtools 軟件對馬鈴薯StPEBP基因家族成員與番茄和辣椒的PEBP 基因家族成員進行共線性分析。利用TBtools 軟件對幾個物種進行全基因組比對,得到馬鈴薯與幾個物種基因之間的對應關系。對所得到的基因信息、起始位點和終止位點等進行整理歸納,進而用于進一步可視化分析。
1.6 選擇壓力分析
使用TBtools v1.09 軟件進行比對得到馬鈴薯基因對信息,利用馬鈴薯StPEBP家族基因對信息、CDS 序列、蛋白序列進一步計算出非同義替換率(Non-synonymous substitution rate,Ka)和同義替換率(Synonymous substitution rate,Ks)。
1.7 馬鈴薯StPEBP 家族成員的表達分析
利用Potato Genomics Resource 數(shù)據(jù)庫[http:∕∕Spud DB(uga.edu)]中下載的馬鈴薯DM 1-3 516 R44(DM)材料的不同發(fā)育時期以及不同處理的轉錄組數(shù)據(jù),分析StPEBP基因家族的表達情況。并利用在線工具Heatmapper(http:∕∕www2.heatmapper.ca∕)繪制熱圖。
1.8 匍匐莖不同發(fā)育時期的基因表達變化
以馬鈴薯品種‘Desiree’為試驗材料,取彎鉤角度為30°、60°、90°、120°、150°的匍匐莖和不同發(fā)育時期的塊莖。采樣后進行RNA 提取和實時熒光定量PCR(qRT-PCR)分析。采集的樣本在提取RNA之前立即冷凍在液氮中,并保存在-80℃超低溫冰箱。每個樣品進行3次生物學重復。RNA提取使用生工生物工程(上海)股份有限公司EZ-10 DNAaway RNA Mini-Preps Kit 試劑盒,具體操作步驟參考試劑盒說明書。使用Taq Universal SYBR Green Supermix 試劑盒進行定量,以StEf1a為內參基因。反應體系為10 μL:1 μL cDNA,上、下游引物各0.2 μL,SYBR(BioRad)5 μL,ddH2O補齊至10 μL。qRT-PCR 程序為:95℃預變性30 s,95℃變性10 s,60℃退火30 s,72℃延伸30 s,34個循環(huán),重復3 次。根據(jù)2-ΔΔCt法得到StPEBP的相對表達量,使用Graphpad Prism 8 繪制表達圖。qRT-PCR 所用引物見表1。

表1 qRT-PCR引物序列信息Table 1 Primers used for qRT-PCR
2 結果與分析
2.1 馬鈴薯StPEBP 家族基因成員鑒定
利用6 個擬南芥中PEBP 家族基因的蛋白序列比對,最終在馬鈴薯基因組中鑒定出10 個StPEBP家族基因(表2),均具有保守結構域(InterPro:IPR00891)。StPEBP家族基因蛋白的分子量最大值為20.01 kDa(StPEBP9),最小值為9.27 kDa(StPEBP1);等電點分布在5.21(StPEBP4)~9.07(StPEBP3);親水性最大值為-0.08(StPEBP7),最小值為-0.47(StPEBP2)。亞細胞定位預測結果表明,StPEBP家族基因編碼蛋白定位在細胞質中。
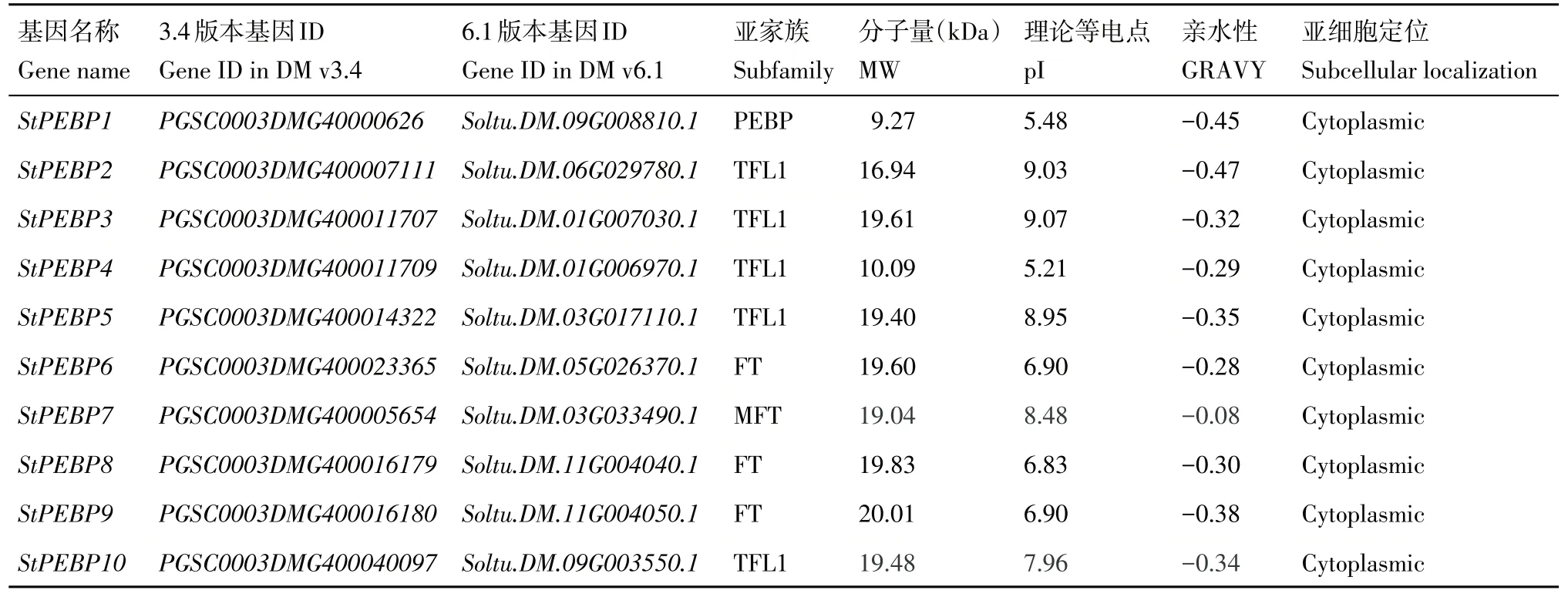
表2 馬鈴薯StPEBP基因家族鑒定與分子特征分析Table 2 Genome-wide identification and molecular characterization of StPEBP gene family in potato
2.2 序列比對與進化樹構建
利用MEGA 7.0軟件將5個物種(擬南芥、番茄、辣椒、煙草和馬鈴薯)PEBP基因家族蛋白序列進行多序列比對,并通過鄰接法構建進化樹(圖1),該家族基因可分為4個亞家族,第一亞家族為PEBPlike 亞家族(StPEBP1),第二亞家族為MFT-like亞家族(StPEBP7),第三亞家族為FT-like 亞家族包括FT(StSP6A)和TSF(StSP3D和StSP5G),第四亞家族為TFL-like 亞家族包括ATC(StPEBP2)、BFT(StPEBP3、StPEBP4和StPBEP5)和TFL(StPEBP10)。StSP6A(StPEBP6)屬于FT-like 亞家族中的FT 小亞家族,StSP5G(StPEBP9)屬于FT-like 亞家族TSF 小亞家族。TFL-like 亞家族是馬鈴薯StPEBP家族中基因數(shù)量最多的亞家族,有5 個 成 員,PEBP-like 亞 家 族 和MFT-like 亞 家 族是StPEBP基因家族中基因數(shù)量最少的亞家族,均僅有1 個成員。同時分析了馬鈴薯與其他物種PBEP 基因家族進化關系,可見馬鈴薯PBEP 基因家族與茄科植物番茄在進化過程中親緣關系更為接近。
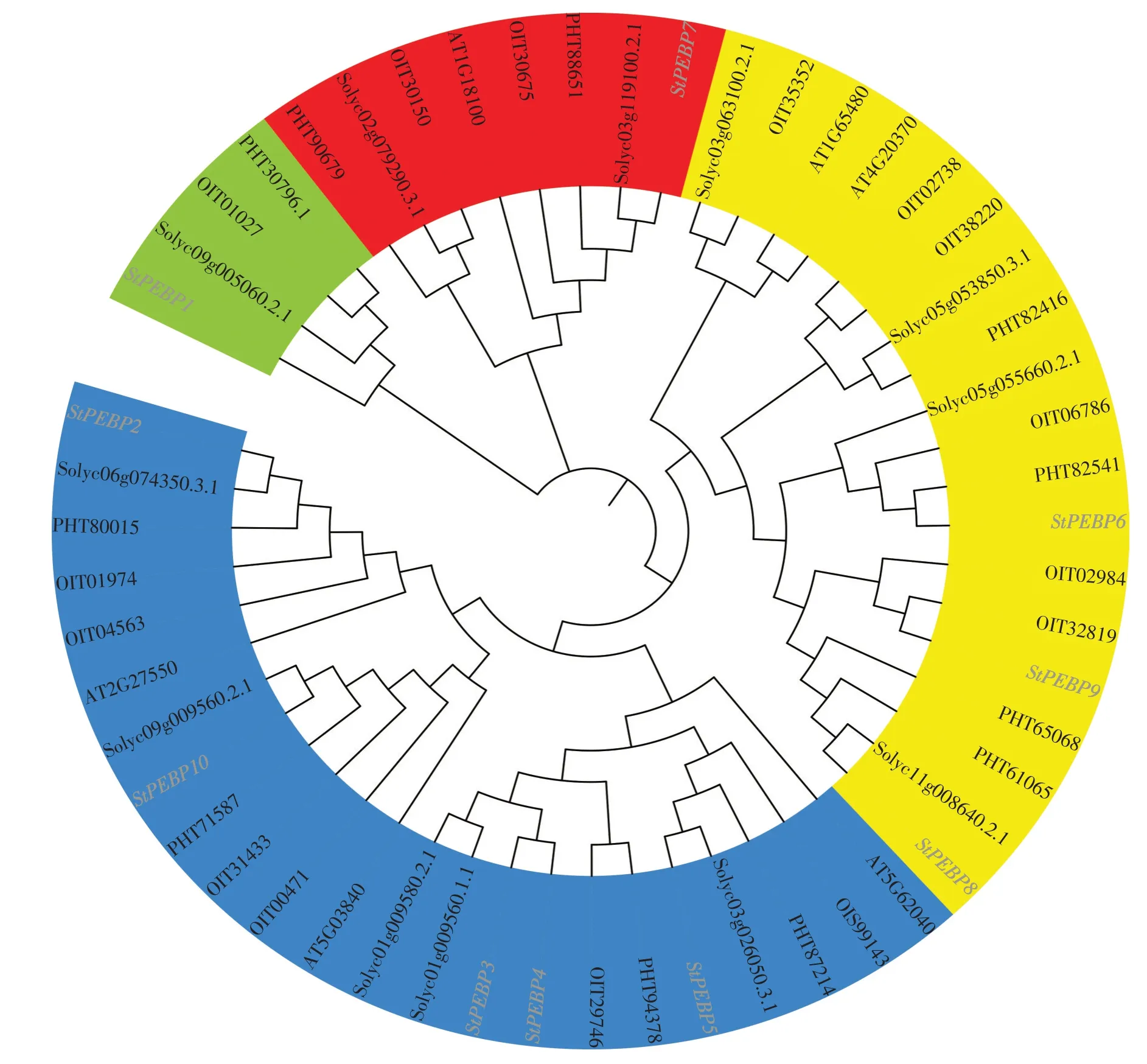
圖1 馬鈴薯、番茄、煙草、辣椒和擬南芥PEBP基因家族進化分析Figure 1 Relationship analysis of PEBP gene family in potato,tomato,tobacco,pepper and Arabidopsis thaliana
2.3 基因結構、motif以及domain預測
根據(jù)已鑒定的馬鈴薯StPEBP基因家族成員的蛋白序列和全基因組的注釋序列,對基因結構進行分析。除StPEBP1(PEBP-like)和StPEBP4(TFL1-like)包含2 個外顯子和1 個內含子,家族其他成員均包含4 個外顯子和3 個內含子(圖2)。結構域分析結果表明,鑒定出的10 個StPEBP基因均具有PEBP superfamily 保守結構域,證實他們均為StPEBP基因家族成員。motif 序列分析,除StPEBP4外所有基因均具有特征性的保守motif序列motif 5。除StPEBP1和StPEBP4外,其余所有基因均具有保守的基序motif 1、motif 2 和motif 3;除StPEBP1,StPEBP2和StPEBP4外,余下基因均具有保守的基序motif 1、motif 2、motif 3、motif 4和motif 5。StPEBP1屬 于PEBP-like 亞 家 族,StPEBP2和StPEBP4均屬于TFL1-like 亞家族,其在進化過程中不僅基因結構發(fā)生變異,基因序列也發(fā)生變異。由結果分析可知,除PEBP-like亞家族和TFL1-like 亞家族外,StPEBP家族基因中其他亞家族的基因序列功能較為保守。
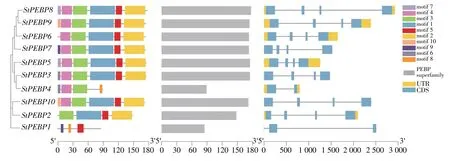
圖2 馬鈴薯StPEBP基因家族進化樹、motif序列與基因結構Figure 2 Phylogenetic tree,motif sequence and gene structure of StPEBP gene family in potato
2.4 順式作用元件預測分析
使用PlareCare 對10 個StPEBP家族基因起始密碼子上游的2 000 bp 進行順式作用元件預測,并用TBtools 進行展示。結果表明,StPEBP家族基因啟動子區(qū)域與光響應相關的元件最多。同時,在StPEBP基因家族的啟動子中存在許多激素響應元件,如水楊酸響應元件、茉莉酸響應元件、生長素響應元件和脫落酸響應元件,其也可能在調控塊莖形成中發(fā)揮重要作用。StPEBP家族基因啟動子中也含有許多低溫響應元件、防御元件和應激反應元件,表明StPEBP家族基因可能在應激反應防御中發(fā)揮作用(圖3)。
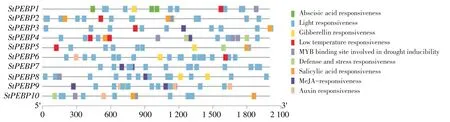
圖3 馬鈴薯StPEBP基因家族順式作用元件預測Figure 3 Prediction of cis-acting elements of StPEBP gene family in potato
2.5 選擇壓力分析
以擬南芥PEBP 基因家族為對照估算馬鈴薯StPEBP基因家族在進化過程中的選擇壓力(表3)。馬鈴薯StPEBP基因家族的所有基因Ka∕Ks 均小于1,受到純化選擇,馬鈴薯中StPEBP基因家族較擬南芥PEBP基因家族發(fā)生了進化。
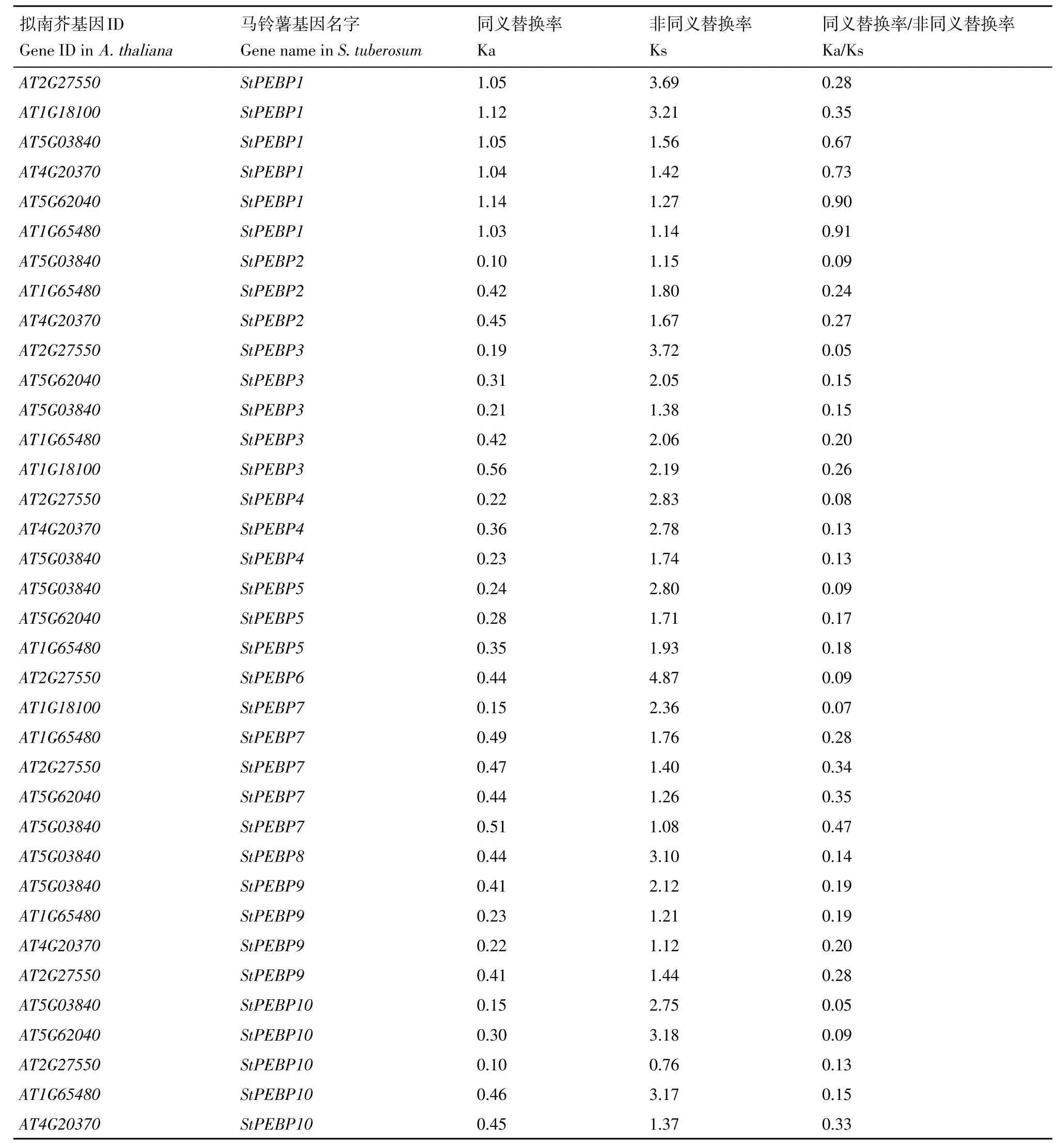
表3 馬鈴薯和擬南芥PEBP選擇壓力分析Table 3 Selective pressure analysis of PEBP in potato and Arabidopsis thaliana
2.6 染色體定位分析及共線性分析
為了進一步分析其基因復制情況,首先對StPEBP基因家族進行染色體定位分析(圖4),StPEBP基因家族在染色體上分布并不均勻,僅分布在1、3、5、6、9 和11 號染色體上,1、3、9、11 號染色體上含有2 個StPEBP基因,5、6 號染色體上各含有1 個StPEBP基因。此外,進一步分析了馬鈴薯基因組內StPEBP基因家族的復制情況(圖4A),馬鈴薯與茄科植物番茄和辣椒之間的PEBP基因的共線性情況(圖4B,圖4C)。馬鈴薯基因組內存在兩對重復基因,即StSP6A和StSP3D、StPEBP2和StPEBP4。將馬鈴薯分別與茄科植物番茄和辣椒之間PEBP 家族基因進行共線性分析,發(fā)現(xiàn)番茄和馬鈴薯間存在11 對共線性基因(圖4B,表4),辣椒和馬鈴薯之間存在8 對共線性基因(圖4C,表4)。結果表明,馬鈴薯StPEBP家族與番茄PEBP 家族共線性較馬鈴薯StPEBP家族與辣椒PEBP家族同源性高,并且在進化過程中更為保守。
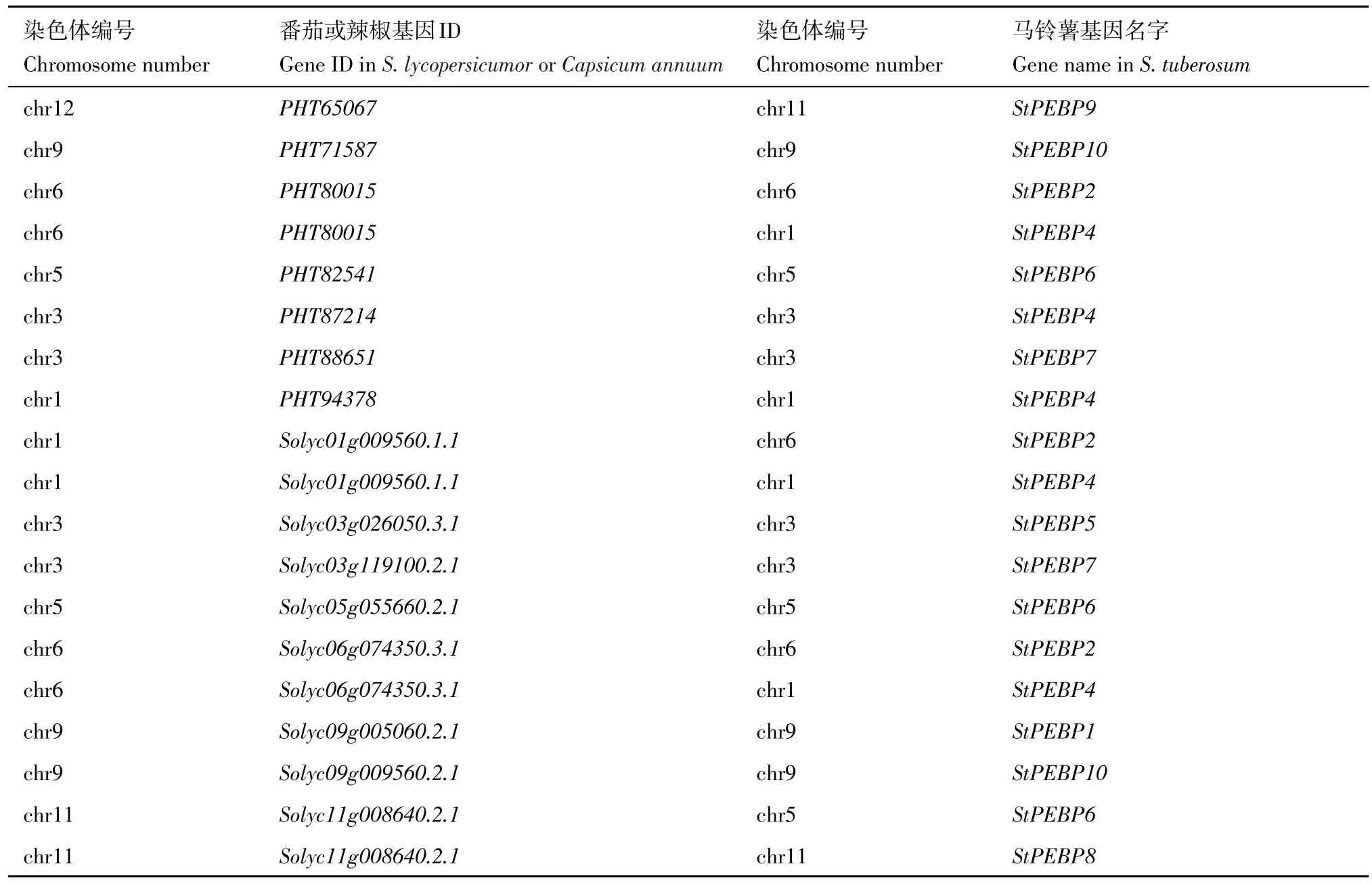
表4 馬鈴薯StPEBP和番茄及辣椒共線性基因Table 4 Synteny analysis of StPEBP gene family in potato,tomato and pepper
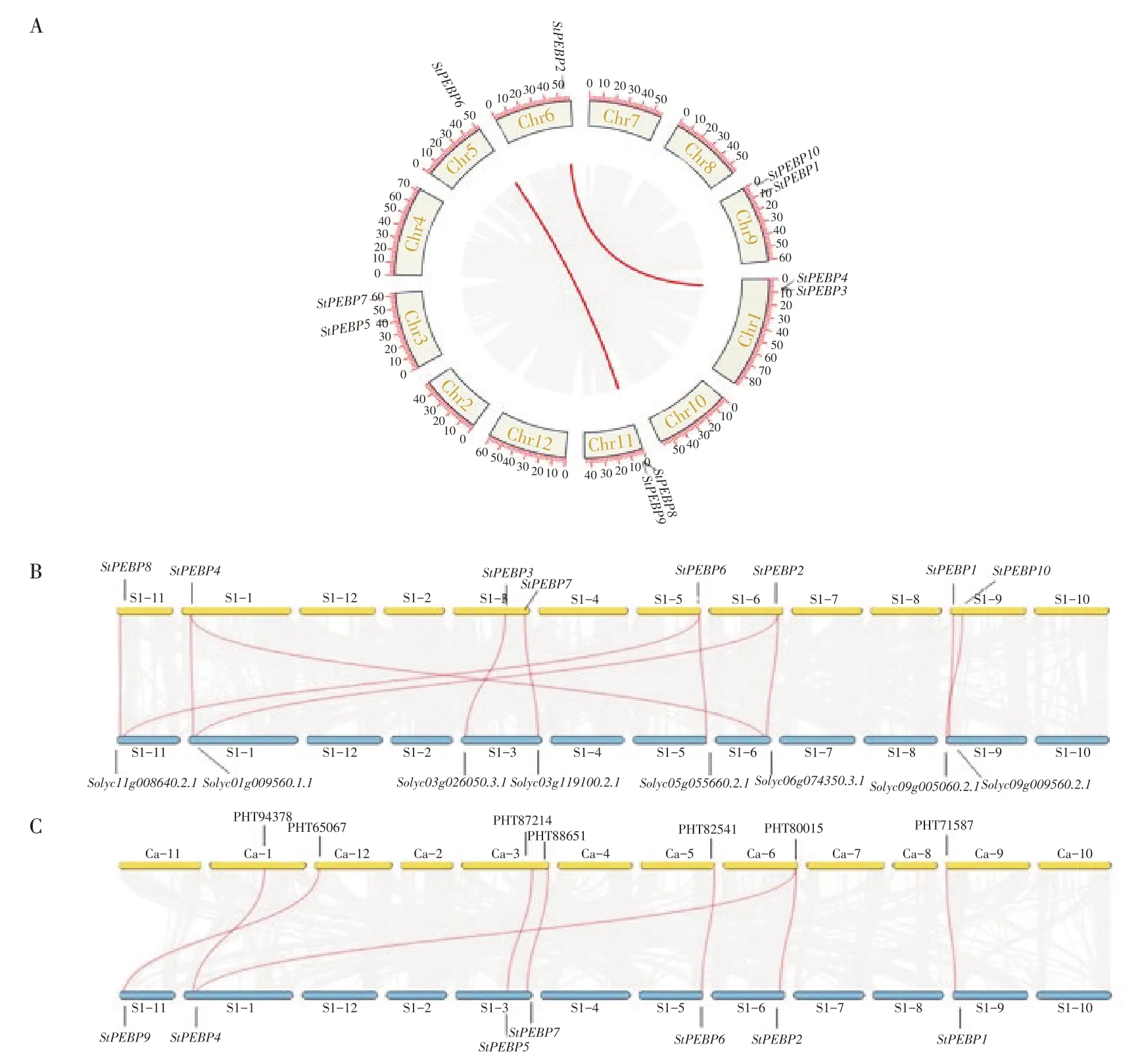
圖4 馬鈴薯PEBP基因家族共線性分析Figure 4 Synteny analysis of PEBP gene family in potato
2.7 StPEBP 基因家族成員的組織表達分析
在Potato Genomics Resource 數(shù)據(jù)庫中StPEBP基因家族組織表達的轉錄組數(shù)據(jù)表明,7 個StPEBP家族基因在馬鈴薯的15 個組織中均呈現(xiàn)出高表達水平,而一些基因在1 或2個組織中表達較低(圖5)。例如,StPEBP3在葉片和愈傷組織中分別表現(xiàn)出低表達水平,而StSP3D(StPEBP8)在葉柄中表現(xiàn)出低表達水平,StSP5G(StPEBP9)在果實內部表現(xiàn)出低表達水平。但是大部分基因在馬鈴薯的各個組織中均高表達,因此,推斷StPEBPs基因家族也可能參與馬鈴薯植株的發(fā)育。

圖5 StPEBP基因家族在馬鈴薯不同組織中的表達分析Figure 5 Expression analysis of StPEBP gene family in different tissues of potato
為了進一步研究StPEBP家族基因在馬鈴薯不同組織中的表達模式,進行了qRT-PCR。StPEBP1在小薯中表達水平最高,其次莖尖、根和大薯中表達水平較高,在其他組織中表達水平較低。StPEBP2僅在根和莖尖表達水平較高,在其他組織中表達水平較低。StPEBP3在小薯中表達水平最高,在花蕾和莖中表達水平較高,其次是大薯,在其他組織中表達水平較低。StPEBP4在花蕾中表達水平最高,其次是葉片,在其他組織部位表達水平較低。StPEBP5和StPEBP7在花蕾中表達水平最高,其次是莖,在其他組織中表達水平相似且較低。StSP3D(StPEBP8)在莖中表達水平最高,其次是葉片,在莖尖、花蕾和匍匐莖表達量相似且較高,在其他組織部位的表達水平較低(圖6)。
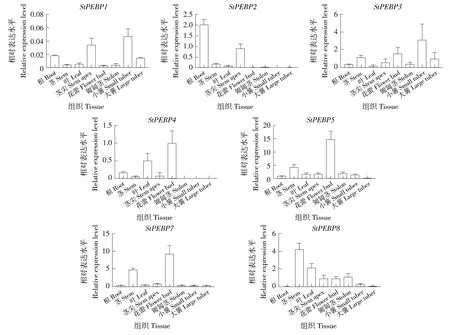
圖6 馬鈴薯StPEBP基因家族在不同組織中的表達模式Figure 6 Expression patterns of StPEBP gene family in different tissues of potato
2.8 StPEBP 基因家族在塊莖發(fā)育中的表達分析
為了探索StPEBP 基因家族在塊莖發(fā)育過程中的作用,使用qRT-PCR 檢測其在不同發(fā)育階段的匍匐莖和塊莖中的表達水平(圖7)。隨著匍匐莖彎鉤角度從60°到120°變化StPEBP1 表達量增加,在匍匐莖彎鉤角度為150°時StPEBP1 表達量急劇下降,但其在小薯中表達量很高,大薯中表達量極低。StPEBP2隨著匍匐莖彎鉤角度從60°到120°變化表達量增加,匍匐莖彎鉤角度達到150°后StPEBP2表達量下降并保持穩(wěn)定。StPEBP3在大薯中表達量最高,其次在彎鉤角度為150°的匍匐莖和小薯中表達量較高,在匍匐莖發(fā)育的其他階段表達量極低。StPEBP5在小薯中表達量最高,在匍匐莖其他發(fā)育階段表達量極低。在彎鉤角度為150°的匍匐莖中,StSP6A(StPEBP6)的表達量明顯增加,小薯和大薯中StSP6A的表達量也較高,在其他時期表達量較低。StPEBP7在匍匐莖彎鉤角度為120°時表達量最高,在大薯中的表達量類似于匍匐莖彎鉤角度為150°時的表達量,低于匍匐莖彎鉤角度為120°時表達量。StSP3D(StPEBP8)在匍匐莖彎鉤角度為30°時表達量最高,其次在匍匐莖彎鉤角度為90°和小薯中表達量較高,其他時期表達量較低。StSP5G(StPEBP9)在匍匐莖彎鉤角度為120°時表達量最高,在匍匐莖彎鉤角度為30°和90°時表達量較高,其他時期表達量較低。StPEBP10在匍匐莖彎鉤角度為60°時表達量最高,在匍匐莖彎鉤角度為90°時和大薯中表達量較高。
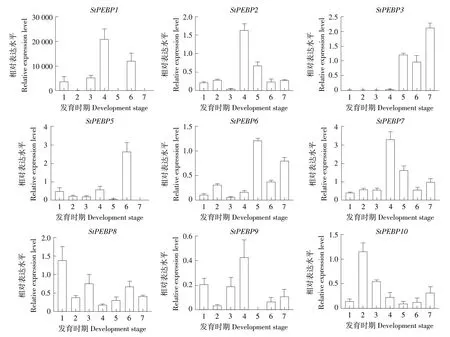
圖7 馬鈴薯StPEBP基因家族在匍匐莖發(fā)育過程中的表達模式Figure 7 Expression pattern of potato StPEBP gene family during stolon development in potato
3 討 論
PEBP 基因家族不僅在開花調節(jié)通路中發(fā)揮重要作用,還參與了植物種子萌發(fā)和非生物脅迫響應等調控過程[23]。在馬鈴薯中,StPEBP基因家族成員StSP5G、StSP6A和StSP3D已鑒定與馬鈴薯塊莖誘導及開花有關,但其他成員在塊莖形成中的作用依然未知。因此本試驗在鑒定馬鈴薯DM v3.4 版本中StPEBP家族基因具有10 個成員后,并分析了其在塊莖發(fā)育中可能發(fā)揮的功能。同時,進行了StPEBP基因家族的保守基序、基因和蛋白結構、順式作用元件、染色體定位、共線性關系及組織表達分析。
系統(tǒng)發(fā)育分析揭示了馬鈴薯中StPEBP家族基因與擬南芥中PEBP 家族基因亞家族分類并不一致,擬南芥中PEBP 家族基因分為3 個亞家族,分別是TFL1-like、MFT-like 和FT-like[24],而馬鈴薯中StPEBP家族基因分為4 個亞家族,分別是TFL1-like、MFT-like、FT-like 和PEBP-like。通過系統(tǒng)發(fā)育分析結果表明,PEBP-like 亞家族與StPEBP家族基因其他亞家族親緣關系較遠。并且在馬鈴薯中,只有StPEBP1屬于PEBP-like 亞家族,該亞家族并未在擬南芥、蘋果和水稻中發(fā)現(xiàn),但在茄科植物煙草、辣椒和番茄中各鑒定出1 個PEBP-like 亞家組基因,推測PEBP-like 亞家組應該是在進化過程中茄科植物所特有的亞家族,可能具有一定功能,今后進行深入探究。此前,有報道稱MFT-like 亞家族是FT-like 亞家族和TFL1-like 亞家族的祖先。在苔蘚植物和石松屬植物中,沒有FT-like 亞家族和TFL1-like 亞家族同源基因,但分別有4 和2 個MFT-like 亞家族同源基因,表明FT-like 亞家族和TFL1-like 亞家族基因是隨種子植物的進化而出現(xiàn)的[3],因此推測馬鈴薯中FT-like 亞家族和TFL1-like 亞家族基因也是在進化過程中而出現(xiàn)的。在馬鈴薯和番茄中,已經(jīng)報道具有影響馬鈴薯結薯和番茄果實成熟功能的基因均屬于FT-like 亞家族[6],由此猜測PEBP 基因家族中FT-like 亞家族基因是影響茄科植物果實成熟和開花時間的重要基因,對于茄科植物果實發(fā)育具有重要的研究價值。
通過對馬鈴薯StPEBP家族基因的motif 分析,發(fā)現(xiàn)10 個StPEBP家族基因除StPEBP4外均具有保守的基序motif 5,這說明motif 5 是馬鈴薯StPEBP基因家族中最保守的序列,也可能是StPEBP家族基因發(fā)揮其生物學功能的基序。同時,分析了4 個亞家族的基序保守性,大部分StPEBP家族基因均具有motif 1~5,F(xiàn)T-like 亞家族和MFT1-like 亞家族基序是保守的,均具有motif 1~5,而PEBP-like亞家族和TFL1-like亞家族基序發(fā)生變化,StPEBP1(PEBP-like)僅有motif 5,StPEBP2(TFL1- like)缺 少motif 4,StPEBP4(TFL1-like)卻僅有motif 3 和motif 4。在本試驗中,PEBP-like 亞家族基因(StPEBP1)和TFL1 亞家族基因(StPEBP4)的內含子和外顯子數(shù)量也發(fā)生了變化,推測可能是在進化過程中PEBP-like亞家族和TFL1-like 亞家族發(fā)生變異,而其他亞家族結構在進化過程中非常保守。
共線性分析結果顯示StSP6A(StPEBP6)和StSP3D(StPEBP8)是重復基因,序列比對發(fā)現(xiàn)StSP6A和StSP3D氨基酸序列一致性高達70%。已有文獻報道[25,26],F(xiàn)T-like 亞家族和TFL1-like亞家族在植物中功能相反,主要是由于含有LGRQTVYAPGWRQN的氨基酸片段和三聯(lián)體LYN的關系,同時FT-like亞家族含有LGRQTVYAPGWRQN氨基酸片段在植物中是保守的并具有功能的。序列比對中同樣發(fā)現(xiàn)馬鈴薯StSP6A中具有LGRQTVYAPGWRQN 氨基酸片段中的一部分TVYAPGWRQN 片 段,StSP3D具 有TVYAPNWRQN片段,與StSP6A中TVYAPGWRQN 片段相比,僅有一個氨基酸序列改變,G 改變?yōu)镹,在馬鈴薯StSP6A和StSP3D中已被鑒定與馬鈴薯結薯和開花有關,因此推斷StPEBP家族中具有LGRQTVYAPGWRQN 氨基酸片段的基因,對于研究馬鈴薯結薯和開花意義較大。
通過表達模式分析發(fā)現(xiàn),當匍匐莖彎鉤角度為150°時,StSP6A(StPEBP6)的表達量急劇增加,在小薯和大薯中表達量也較高。這再次證實了StSP6A在塊莖形成過程中發(fā)揮了重要的作用。除StPEBP6外,StPEBP1隨著匍匐莖發(fā)育表達量增加,并且在小薯中高表達,StPEBP3和StPEBP5表達量也在塊莖中高表達。推測,StPEBP1、StPEBP3和StPEBP5也可能在馬鈴薯塊莖形成的調控中發(fā)揮作用。通過qRT-PCR檢測了StPEBP1和StPEBP3在不同組織中的表達模式,發(fā)現(xiàn)StPEBP1和StPEBP3同樣在塊莖中高表達,這進一步證實了之前的推測,即StPEBP1和StPEBP3可能在馬鈴薯塊莖誘導途徑中具有重要意義,但其功能有待進一步研究。
本試驗利用生物信息學和qRT-PCR技術分析了馬鈴薯StPEBP家族基因在塊莖誘導中的功能。除StSP5G和StSP6A外,通過表達模式分析和共線性分析猜測StPEBP1、StPEBP3和StSP3D在馬鈴薯塊莖形成中發(fā)揮重要功能,這些基因可作為候選基因用于進一步研究PEBP家族基因在塊莖誘導中的功能。
猜你喜歡
現(xiàn)代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業(yè)技術(2016年15期)2016-12-01 05:31:22
當代經(jīng)濟研究(2016年5期)2016-12-01 03:12:05
現(xiàn)代農業(yè)(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫(yī)藥現(xiàn)代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
