辣椒細胞質雄性不育系小孢子敗育的細胞學形態鑒定
2022-06-28 01:17:28周坤華陳學軍黃月琴袁欣捷
江西農業學報 2022年4期
雷 剛,周坤華,陳學軍,黃月琴,袁欣捷,方 榮
(江西省農業科學院 蔬菜花卉研究所,江西 南昌 330200)
0 引言
辣椒(Capsicum annuum L.)是我國栽培面積最大的蔬菜作物之一,年種植面積超過210萬hm2,總 產 量6400多 萬t[1]。辣 椒 果 實 中 含 有 豐 富 的辣椒素、辣椒堿、維生素、辣椒色素、有機酸、蛋白質、糖和礦物質等多種功能性成分,除了應用在食品行業外,還被用于飼料、化妝品、工業染料、醫藥及軍事等行業[2-3]。辣椒屬于常異花授粉植物,具有較強的雜種優勢,F1代雜交種子比常規種早期增產達50%左右,全生育期產量增加30%~50%[4]。但目前人工雜交制種不僅存在成本高、授粉效率低、種子純度難以保證等問題,而且親本材料容易流失[5-7],利用細胞質雄性不育系(cytoplasmic male sterile,CMS)進行雜交制種可有效解決這一難題。
有關辣椒CMS小孢子敗育細胞學的研究已有諸多報道,小孢子敗育的時期因材料的不同而異,敗育現象從造孢細胞時期之后每個階段都有發生[8]。掌握敗育發生的時期和成因對于雄性不育的轉育和在生產上的應用意義重大。本研究通過對辣椒CMS B351A及其保持系B351B的小孢子發育的細胞學形態進行比較和鑒定,確定不育發生的時期,推測其發生的細胞學機制,進而為B351A花藥敗育發生的分子生物學研究和不育性狀育種應用提供參考和理論指導;同時,對小孢子發育時期與對應花蕾外觀形態特征的相關性進行分析,旨在確定從花蕾的形態特征來判斷其小孢子發育時期的標準,為辣椒小孢子培養接種材料和分子生物學研究材料的便捷選取提供依據。
1 材料與方法
1.1 試驗材料
供試辣椒細胞質雄性不育系材料B351A(S/rfrf)及其保持系B351B(N/rfrf),由江西省農業科學院蔬菜花卉研究所茄果類蔬菜課題組發現并選育。B351A的細胞質來源于2007年在試驗田發現的細胞質雄性不育植株,其花藥為紫色、干癟皺縮,開裂后無花粉,柱頭外露,雌蕊發育正常;其保持系B351B的花藥也為紫色、飽滿,開裂后花粉量大(圖1)。

圖1 辣椒雄性不育材料B351A(A)及其保持系材料B351B(B)的花器官形態
材料于2020年2月中旬播種,每個材料播100粒種子,苗期晝夜溫度設為(28±2)℃/(15±2)℃,光周期12 h,光照強度180 μmol/(m2·s),相對濕度(65±5)%。3月下旬定植于試驗溫室,管理同常規生產。
1.2 試驗方法
1.2.1 花蕾的測量及形態描述 于盛花期早上8:00~9:00采集不同大小(縱徑2.00~8.00 mm)的花蕾。花蕾的測量采用游標卡尺,分別測量花蕾縱徑,即花蕾基部至花冠伸出點/花冠頂端的長度;橫徑,花萼中部位置橫切面的直徑;花冠長度,花冠漏出花萼部分的長度。對花蕾外觀形態,如花萼形態、花冠形態、色澤等進行描述。
1.2.2 花藥小孢子發育的細胞學觀察 (1)固定:取辣椒雄性不育材料B351A及其保持系B351B經過測量的花蕾,用鑷子小心剝去花萼、花冠露出花藥后迅速置于電鏡固定液中(2.5%戊二醛,0.1 mol/L磷酸緩沖液,pH值為7.0~7.5)固定,并用真空泵抽氣直至沉底,室溫放置2 h,后轉入4 ℃冰箱,隨后用0.1 mol/L磷酸緩沖液PB(pH值為7.4)漂洗3次,每次15 min。
(2)后固定:用1.0%的鋨酸·0.1 mol/L磷酸緩沖液PBS(pH值為7.4)室溫固定5 h,隨后用0.1 mol/L磷酸緩沖液(pH值為7.4)漂洗3次,每次15 min。
(3)脫水與透明:將固定好的組織分別用30%、50%、70%、80%、90%、95%、100%的酒精進行脫水,每次1 h。將脫水后的組織材料置于無水乙醇∶丙酮為3∶1、1∶1和1∶3(v∶v)的溶液中分別透明0.5 h,最后置于丙酮溶液中1 h。
(4)滲透與包埋:利用丙酮∶812包埋劑=3∶1(v∶v)滲 透3 h,丙 酮∶812包 埋 劑=1∶1(v∶v)滲透過夜,丙酮∶812包埋劑=1∶3(v∶v)滲透3 h,純812包埋劑滲透6 h;將純812包埋劑倒入包埋板,將樣品插入包埋板后37 ℃烤箱過夜,隨后轉60 ℃烤箱聚合48 h。
(5)切片:采用徠卡的超薄切片機Leica UC7切片,切片厚度1~2 μm。
(6)染色:將甲苯胺藍染液放入烤箱60 ℃加熱,將切片置于染液中浸染2 min,隨后用蒸餾水洗2 min,95%酒精分化,在光鏡下控制顏色,烤干后用中性樹膠封片。最后采用日立HT7700電子透射電鏡進行觀察并拍照。
2 結果與分析
2.1 不育系與保持系小孢子發育過程的細胞學形態觀察比較
2.1.1 花粉母細胞時期(S1) 孢原細胞經過分裂形成了纖維層、中層、絨氈層和花粉母細胞。絨氈層細胞位于花藥壁的最內層,排列整齊緊密,染色很深,造孢細胞則位于絨氈層的內側,體積較大(圖2A)。此時期,不育系與保持系花藥在細胞結構和形態上沒有明顯的差異(圖2B)。
2.1.2 四分體時期(S2) 此時保持系花藥中層細胞消失,其他各層細胞層次分明,絨氈層細胞體積大,細胞核大且著色深;花粉母細胞經過 2次分裂形成染色較深的胼胝質包裹的四面體結構(圖2C)。而不育系花藥室內未見四分體結構,可見一些形狀不規則的花粉母細胞,中層細胞還清晰可見,部分絨氈層細胞明顯液泡化且體積較大,擠壓花藥室,有的絨氈層細胞已經降解成為深色條塊(圖2D)。
2.1.3 單核期-單核靠邊期(S3) 保持系小孢子細胞質中出現液泡,細胞核被擠壓靠近細胞壁;絨氈層細胞已經開始退化(圖2E)。此時不育系絨氈層已經完全降解成為深色條帶;花藥室受擠壓變得狹小,未完成減數分裂的花粉母細胞也完全降解,與絨氈層細胞降解殘渣粘連在一起形成深色條帶(圖2F)。
2.1.4 成熟花粉粒時期(S4) 此時保持系的花粉粒變的很圓,著色深,有的花粉粒可以看到萌發孔;絨氈層細胞幾乎全部降解,僅有部分細胞殘留物在花藥室內壁上(圖2G)。而不育系花藥室內看不到成熟的花粉粒,僅可見解體的花粉母細胞和絨氈層細胞粘連在一起,形成著色很深的條帶(圖2H)。

圖2 辣椒胞質雄性不育系B351A和保持系B351B小孢子發育過程
2.2 B351A和B351B小孢子發育各時期對應花蕾外觀特征
通過電鏡觀察不同大小花蕾對應小孢子的發育時期,從表1可見,小孢子發育的時期與花蕾的縱徑、橫徑、花冠長度及花萼、花冠色澤和狀態有密切的關系。無論是不育系還是保持系,在花粉母細胞時期花萼緊緊包裹花冠;四分體時期花萼微微張開,漏出淺綠色的花冠,從S1到S2花蕾縱徑增加不明顯,但橫徑增加相對較大;單核靠邊期(S3)花萼處于半抱合狀態,花冠長度接近花萼長度,從S2到S3花蕾縱徑和橫徑都明顯增加;花粉成熟期花萼向外張開,花冠露出部分達花萼長度的1/2,中部鼓起,顏色黃白色,從S3至S4花蕾縱徑和花冠長度增加最為明顯,增加的長度主要是花冠長度增加所致,橫徑變化不明顯(表1)。
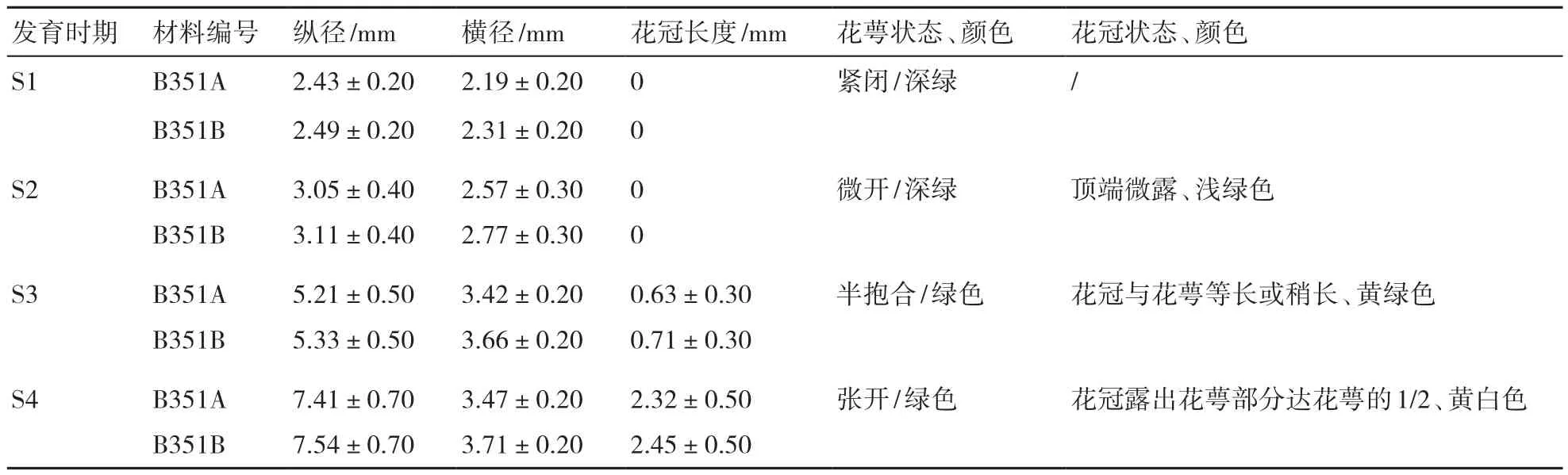
表1 花藥不同發育時期對應花蕾的外觀形態和參數
3 討論與結論
3.1 辣椒胞質雄性不育系B351A小孢子發生敗育的時期
在被子植物中,一般單子葉植物小孢子敗育多發生在雙核期或靠近雙核期,而雙子葉植物多發生在四分體時期或小孢子早期[9]。就辣椒而言,花粉敗育發生在花粉母細胞時期[10-13]、花粉母細胞減數分裂期[14-17]、四分體時期[18-22]、單核期[23-25]等的都有相關報道,這可能是基因型和不育類型的差異所致。本研究發現,在花粉母細胞時期,2個材料花藥的細胞結構沒有明顯的差異(圖2A、2B),但在四分體時期,B351A的絨氈層細胞退化明顯,花藥室內無四分體形成;隨著花藥繼續發育,花粉母細胞逐漸發生分解、退化(圖2F、2H),最終導致無小孢子產生而發生敗育。因此,不育系B351A小孢子敗育發生在花粉母細胞時期至四分體時期之間,即花粉母細胞減數分裂時,這與吳鶴鳴等[14-17]的研究結果相似,皆是在形成四分體前絨氈層細胞的異常或提前降解,導致無四分體或產生畸形的四分體結構而引起敗育。絨氈層是花藥壁的最內層結構,毗鄰小孢子,為小孢子發育提供所需的營養物質、生長素和酶,并適時降解,為小孢子發育提供空間。絨氈層細胞與高等植物的花粉發育有著密切的關系,其結構異常,提早或延后降解都會導致花粉敗育[26]。本研究中,B351B的絨氈層在四分體時期后開始降解,到花粉成熟期已完全降解,而B351A花藥絨氈層在減數分裂期就提前降解,解體的細胞包裹著花粉母細胞,導致花粉母細胞營養不能減數分裂形成四分體結構,沒有小孢子產生而發生敗育。
3.2 辣椒小孢子發育時期與花蕾外觀形態的關系
前人研究發現,辣椒花蕾色澤、縱徑、橫徑,花藥色澤、長、寬,花萼長及花瓣長等外觀指標和形態特征與小孢子發育時期有密切關系,可作為通過外觀形態判斷小孢子發育時期,進行田間取樣的依據[27-29]。本研究對辣椒CMS B351A及其保持系B351B的小孢子發育時期與花蕾外觀形態特征進行了分析,研究結果表明,小孢子各發育時期與花蕾的一些外部形態特征關系密切。花粉母細胞時期或之前,花萼緊緊包裹花冠是最明顯的特征;從花粉母細胞時期到四分體時期,花蕾橫徑增加最為明顯,可能是此時花藥縱向生長速度較快;四分體到單核期,花蕾縱徑和橫徑都明顯增加;當花冠長度接近花萼長度時,小孢子處于單核靠邊期,但單核靠邊期之后,花蕾橫徑幾乎沒變化,花冠的快速伸長導致了縱徑大幅的增加。因此,可以根據花萼狀態判斷是否處于花粉母細胞期前后,根據花冠長與花蕾縱徑比判斷是否處于單核靠邊期,這與張菊平等[27-30]報道的對應花蕾狀態的小孢子發育有些許差異,這可能與所用的材料基因型不同有關。筆者認為,辣椒花蕾大小和外觀形態不僅受基因型的影響,受環境影響也很大,研究結果對供試材料具有一定的指導意義,但不具有普適性。
