阿司匹林干預(yù)腦膠質(zhì)細(xì)胞瘤相關(guān)巨噬細(xì)胞表達(dá)VEGF、bFGF 和YKL-40 成血管生長(zhǎng)因子
2022-06-21 07:48:20陳麗麗王涵武蕾蕾展?jié)?/span>郭素芬于建渤吳影高丹
智慧健康 2022年8期
陳麗麗,王涵,武蕾蕾,展?jié)胤遥诮ú常瑓怯埃叩?/p>
(牡丹江醫(yī)學(xué)院附屬紅旗醫(yī)院 皮膚科,黑龍江 牡丹江 157011)
0 引言
腫瘤血管生成為腫瘤組織提供必需的營(yíng)養(yǎng)成分和氧氣,同時(shí)也是腫瘤組織擴(kuò)散和轉(zhuǎn)移的必要條件。靶向阻擊腫瘤血管生成一直是抗腫瘤轉(zhuǎn)移或抗腫瘤復(fù)發(fā)的熱點(diǎn)。但是,臨床研究發(fā)現(xiàn),應(yīng)用抗腫瘤血管生成類的藥物,卻沒有完全達(dá)到抗腫瘤轉(zhuǎn)移的預(yù)期效果[1]。
腫瘤微環(huán)境(tumor microenvironment,TME)包含腫瘤細(xì)胞和非腫瘤細(xì)胞、炎癥細(xì)胞以及多種促血管生成因子等,這些成分構(gòu)成了腫瘤生長(zhǎng)、轉(zhuǎn)移的局部微環(huán)境,TME對(duì)腫瘤血管的生成起到重要的促進(jìn)作用[2]。血液中的單核細(xì)胞/巨噬細(xì)胞,被炎癥因子募集到腫瘤組織中,經(jīng)細(xì)胞因子刺激極化為腫瘤相關(guān)巨噬細(xì)胞,該細(xì)胞能夠分泌基質(zhì)金屬蛋白酶(matrix metalloproteinase,MMP)、巨噬細(xì)胞源性腫瘤壞死因子(tumor necrosis factor,TNF)及白細(xì)胞介素-1(interleukin-1,IL-1)等細(xì)胞因子而促進(jìn)血管的生成[3],并有助于腫瘤細(xì)胞擺脫免疫系統(tǒng)的攻擊,促進(jìn)腫瘤生長(zhǎng)或轉(zhuǎn)移,是血管生成因子的主要來源[4]。
膠質(zhì)瘤是常見的顱內(nèi)原發(fā)腫瘤,具有新生血管較多并且雜亂的特點(diǎn)[5]。雖然目前以放化療或者手術(shù)治療為主,但是膠質(zhì)瘤具有易侵襲、轉(zhuǎn)移等特點(diǎn),治療效果并不十分理想。證據(jù)表明高腫瘤相關(guān)巨噬細(xì)胞浸潤(rùn)與腫瘤患者的疾病進(jìn)展和總生存率有關(guān)[6]。
研究發(fā)現(xiàn):膠質(zhì)瘤相關(guān)巨噬細(xì)胞與膠質(zhì)瘤的惡性程度密切相關(guān),即膠質(zhì)瘤級(jí)別越高,腫瘤相關(guān)巨噬細(xì)胞越多[7]。又發(fā)現(xiàn)膠質(zhì)瘤中腫瘤相關(guān)巨噬細(xì)胞與微血管密度呈正相關(guān),且級(jí)別越高越顯著[8]。目前關(guān)于膠質(zhì)瘤相關(guān)巨噬細(xì)胞的研究,多集中在膠質(zhì)瘤相關(guān)巨噬細(xì)胞對(duì)膠質(zhì)瘤細(xì)胞的凋亡、侵襲和遷移方面的影響,而膠質(zhì)瘤相關(guān)巨噬細(xì)胞是否通過影響成血管因子的表達(dá)而調(diào)控膠質(zhì)瘤的生長(zhǎng)和轉(zhuǎn)移研究卻較少。
環(huán)氧合酶-2(cyclooxygenase-2,COX-2)對(duì)腫瘤血管的生成有促進(jìn)作用。阿司匹林具有選擇性抑制COX-2的重要作用[9],在阿司匹林的干預(yù)下,是否可能影響膠質(zhì)瘤相關(guān)巨噬細(xì)胞分泌VEGF、bFGF以及YKL-40等成血管因子,是本課題所要解決的問題,研究結(jié)果將對(duì)揭示膠質(zhì)瘤相關(guān)巨噬細(xì)胞與膠質(zhì)瘤發(fā)生發(fā)展關(guān)系探尋靶點(diǎn),并為抗膠質(zhì)瘤做好研究基礎(chǔ)。
1 材料與方法
1.1 材料
1.1.1 細(xì)胞株
人白血病THP-1單核細(xì)胞(購(gòu)于中國(guó)科學(xué)院典型培養(yǎng)物保藏委員會(huì)細(xì)胞庫(kù));人腦星形膠質(zhì)母細(xì)胞瘤U-87MG細(xì)胞(由沈陽(yáng)萬類生物科技公司惠贈(zèng))。
1.1.2 主要試劑和儀器
RPMI-1640或DMEM高糖培養(yǎng)基和胎牛血清(美國(guó)Hyclone公司),佛波酯(Phorbol ester,PMA;美國(guó)sigma公司),人重組白細(xì)胞介素-4蛋白(Interleukin-4,IL-4;美國(guó)BD公司),免抗人CD163(武漢博士得公司),免抗人VEGF單克隆抗體、免抗人FGF2單克隆抗體和山羊抗免IgG(美國(guó)Proteintech公司),超敏型二步法檢測(cè)試劑盒(北京中衫金橋公司),濃縮型DAB試劑盒(北京中衫金橋公司)。人VEGF、bFGF和YKL-40的ELISA試劑盒(美國(guó)R&D公司)。日本OLYMPUS倒置顯微鏡,西班牙Biological超凈工作臺(tái)。
1.2 實(shí)驗(yàn)方法
1.2.1 細(xì)胞培養(yǎng)
含10%胎牛血清的RPMI-1640完全培養(yǎng)液培養(yǎng)THP-1細(xì)胞。含10%FBS的DMEM高糖完全培養(yǎng)基培養(yǎng)U-87MG細(xì)胞。
1.2.2 細(xì)胞誘導(dǎo)和分組
3×105/mL人白血病THP-1單核細(xì)胞,添加含PMA的RPMI-1640完全培養(yǎng)基,37℃,5%CO2培養(yǎng)24h后,更換含白細(xì)胞介素-4(Interleukin-4,IL-4)的DMEM高糖完全培養(yǎng)基;37℃,5%CO2培養(yǎng)48h,誘導(dǎo)為M2型巨噬細(xì)胞,單獨(dú)培養(yǎng)M2型巨噬細(xì)胞為M2組。單獨(dú)培養(yǎng)U-87MG細(xì)胞為U-87M組。U-87MG細(xì)胞與M2型巨噬細(xì)胞非接觸共同培養(yǎng)為共培養(yǎng)M2組,共培養(yǎng)細(xì)胞用2mmol/L、4mmol/L、8mmol/L阿司匹林進(jìn)行干預(yù)分別為2mM、4mM、8mM阿司匹林組。
1.2.3 非接觸共培養(yǎng)體系
3×105/mL人M2型巨噬細(xì)胞接種在孔徑0.4μm的Transwell培養(yǎng)板下室,DMEM高糖完全培養(yǎng)基培養(yǎng)。2×105/mL U-87MG細(xì)胞用DMEM高糖完全培養(yǎng)基重懸,接種至Transwell培養(yǎng)板上室;37℃,5%CO2培養(yǎng)12h后,待U-87MG細(xì)胞完全貼壁,Transwell培養(yǎng)板上室內(nèi)添加0mmol/L、2mmol/L、4mmol/L、8mmol/L阿司匹林,37℃,5%CO2培養(yǎng)24h。
1.2.4 人M2 型巨噬細(xì)胞的鑒定
1×104THP-1細(xì)胞種植于附有玻片的24孔板內(nèi),然后經(jīng)PMA和人重組IL-4誘導(dǎo)THP-1細(xì)胞至貼壁并融合達(dá)70%以上;4%多聚甲醛固定30min;3%BSA工作液封閉,室溫放置20min;添加免抗人CD163抗體(1:200),4℃孵育過夜;0.01mmol/l PBS磷酸鹽緩沖液清洗3次,每次3min;山羊抗免IgG(1:300),37℃孵育40min;0.01mmol/l PBS磷酸鹽緩沖液清洗3次,每次2min;滴加DAB工作液,蘇木素輕度復(fù)染3min。顯微鏡觀察并照相。
1.2.5 免疫組化法檢測(cè)VEGF、bFGF 的表達(dá)
取上述各組中單獨(dú)培養(yǎng)的細(xì)胞和共培養(yǎng)M2型巨噬細(xì)胞及阿司匹林作用的M2型巨噬細(xì)胞,經(jīng)固定、沖洗、封閉等處理后加入VEGF、bFGF一抗,4℃過夜;余步驟同M2型巨噬細(xì)胞的鑒定。
1.2.6 ELISA
3.5×104各組細(xì)胞,37℃,5%CO2培養(yǎng)12h,提取各組細(xì)胞的上清液。使用人VEGF、bFGF地ELISA試劑盒檢測(cè)各組細(xì)胞VEGF、bFGF的含量。
1.3 觀察指標(biāo)
需要與結(jié)果2.1,2.2,2.3對(duì)應(yīng)。
1.4 統(tǒng)計(jì)學(xué)分析
2 結(jié)果
2.1 THP-1細(xì)胞形態(tài)學(xué)特征
THP-1從血液中被募集到組織中,被細(xì)胞因子激活,轉(zhuǎn)化為TMAs。THP-1細(xì)胞經(jīng)2-3次傳代,狀態(tài)穩(wěn)定良好的情況下,細(xì)胞懸浮呈圓形,大小均勻,折光度較好。在培養(yǎng)液中添加含濃度為100ng/mL PMA的RPMI-1640完全培養(yǎng)基24h,細(xì)胞完全貼壁,形態(tài)由圓形轉(zhuǎn)變?yōu)槁褕A形或梭形有隆起。添加含濃度為20ng/mL IL-4的RPMI-1640完全培養(yǎng)基24h,部分細(xì)胞伸出偽足,形態(tài)變長(zhǎng)。48h后,細(xì)胞形態(tài)明顯改變,偽足延長(zhǎng)變細(xì),圖1。
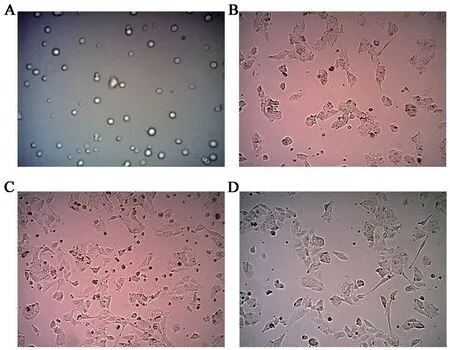
圖1 各組THP-1細(xì)胞培養(yǎng)24h的形態(tài)學(xué)特征(×100)
2.2 M2型巨噬細(xì)胞的表型鑒定
免疫組化結(jié)果表明:與THP-1細(xì)胞誘導(dǎo)之前CD163表達(dá)(0.37%)相比,THP-1細(xì)胞經(jīng)PMA及IL-4逐級(jí)誘導(dǎo)后出現(xiàn)高表達(dá)的M2型巨噬細(xì)胞特異性抗原CD163(93.7%),兩組數(shù)據(jù)相比具有顯著差異(P<0.01),圖2。

圖2 THP-1細(xì)胞經(jīng)PMA及IL-4誘導(dǎo)后CD163的表達(dá)(×400)
2.3 ELISA檢測(cè)各組細(xì)胞VEGF、bFGF和YKL-40含量
單獨(dú)培養(yǎng)的M2型巨噬細(xì)胞及與U87MG細(xì)胞共同培養(yǎng)的M2型巨噬細(xì)胞均見VEGF,bFGF和YKL-40的表達(dá),但共培養(yǎng)的M2型巨噬細(xì)胞VEGF,bFGF和YKL-40表達(dá)明顯增多,數(shù)據(jù)相比具有統(tǒng)計(jì)學(xué)有意義(P<0.01)。與共同培養(yǎng)的M2型巨噬細(xì)胞相比較,當(dāng)在共培養(yǎng)體系中加入2mm、4mm、8mm不同濃度阿司匹林后,阿司匹林組VEGF,bFGF和YKL-40含量逐漸降低,數(shù)據(jù)相比具有統(tǒng)計(jì)學(xué)有意義(P<0.01)。當(dāng)阿司匹林濃度達(dá)8mm時(shí),共培養(yǎng)體系M2型巨噬細(xì)胞VEGF,bFGF和YKL-40含量較2mm和4mm阿司匹林干預(yù)的M2型巨噬細(xì)胞明顯最低,數(shù)據(jù)相比具有統(tǒng)計(jì)學(xué)有意義(P<0.01),表1。
表1 ELISA檢測(cè)各組細(xì)胞VEGF、bFGF含量(,n=16,pg/mL)

表1 ELISA檢測(cè)各組細(xì)胞VEGF、bFGF含量(,n=16,pg/mL)
注:*P<0.01,與U-87M組和M2組比較;#P<0.01,與共培養(yǎng)M2組比較
3 討論
膠質(zhì)瘤是一種頑固、常見的顱內(nèi)原發(fā)腫瘤,具有新生血管較多并且雜亂的特點(diǎn)[5]。臨床目前主要進(jìn)行放化療或者手術(shù)治療,但膠質(zhì)瘤具有易侵襲、轉(zhuǎn)移等特點(diǎn),導(dǎo)致以上治療效果一直不理想,因此我們需要不斷地尋找新的治療方法,為臨床治療膠質(zhì)瘤提供新思路。
在腫瘤組織中可以發(fā)現(xiàn)各種免疫細(xì)胞。血液中的單核細(xì)胞/巨噬細(xì)胞,被炎癥因子募集到腫瘤組織中,經(jīng)細(xì)胞因子刺激極化為腫瘤相關(guān)巨噬細(xì)胞(TAMs)。從功能表型上,巨噬細(xì)胞大致分為:經(jīng)典(M1)型和替代(M2)型巨噬細(xì)胞。M1型巨噬細(xì)胞能夠被γ-干擾素等誘導(dǎo),主要抗感染,抑制腫瘤細(xì)胞生長(zhǎng)。M2型巨噬細(xì)胞分泌多種因子,有助于腫瘤細(xì)胞擺脫免疫系統(tǒng)的攻擊,從而促進(jìn)腫瘤生長(zhǎng)轉(zhuǎn)移。研究發(fā)現(xiàn),M2型巨噬細(xì)胞經(jīng)腫瘤誘導(dǎo)更接近TAMs。有資料顯示:TAMs在腫瘤微環(huán)境中扮演重要角色,TAMs是血管生成因子的主要來源[10],如M2型巨噬細(xì)胞可以釋放血管內(nèi)皮生長(zhǎng)因子(Vascular endothelial growth factor,VEGF)、基質(zhì)金屬蛋白酶(Matrix metalloproteinase,MMP)等直接促進(jìn)血管的生成[3],還可以釋放巨噬細(xì)胞源性腫瘤壞死因子(Tumor necrosis factor,TNF)及白細(xì)胞介素-1(Interleukin-1,IL-1)等促進(jìn)黑色素瘤中白細(xì)胞介素-8(Interleukin-8,IL-8)和VEGF的釋放,從而間接促進(jìn)腫瘤的血管生成[11]。越來越多的證據(jù)表明高TAMs浸潤(rùn)與腫瘤患者的疾病進(jìn)展和總生存率有關(guān)。
VEGF和bFGF是最常見的成血管因子,其促進(jìn)血管形成的機(jī)制已有大量報(bào)道,如它們可以促進(jìn)內(nèi)皮細(xì)胞有絲分裂,也可以動(dòng)員內(nèi)皮祖細(xì)胞遷移并摻入到成熟內(nèi)皮細(xì)胞中而發(fā)揮促血管生成作用[12]。YKL-40是一種分泌型糖蛋白,通常由多種細(xì)胞如巨噬細(xì)胞,軟骨細(xì)胞和血管平滑肌細(xì)胞表達(dá)[13]。大量證據(jù)表明YKL-40不僅是急性及慢性炎癥標(biāo)志物,而且參與細(xì)胞的增殖、分化、凋亡、血管生成及組織重構(gòu)等過程[14]。
阿司匹林是經(jīng)典的非甾體抗炎藥,具有選擇性抑制環(huán)氧合酶-2(cyclooxygenase-2,COX-2)的作用。已有研究表明COX-2對(duì)腫瘤血管的生成有促進(jìn)作用。COX-2對(duì)腫瘤血管形成的機(jī)制為:①促進(jìn)血管內(nèi)皮類生長(zhǎng)因子的表達(dá)如VEGF;②前列腺素類產(chǎn)物如血栓素A2、前列腺素E2、前列環(huán)素能促進(jìn)內(nèi)皮細(xì)胞的遷移及血管形成;③通過上調(diào)Bcl-2的表達(dá)或Akt基因的活性,促進(jìn)血管內(nèi)皮細(xì)胞的形成。Kale和Gan的研究證實(shí):M2型巨噬細(xì)胞可以產(chǎn)生COX-2。那么,在阿司匹林的干預(yù)下,是否可能通過抑制COX-2來影響TAMs成血管因子VEGF、bFGF以及YKL-40的表達(dá)呢?這也是本課題所要解決的問題。
鑒于以上原因,實(shí)驗(yàn)中我們采用共培養(yǎng)系統(tǒng)首先設(shè)計(jì)了常見的VEGF、bFGF以及YKL-40影響血管形成因子研究。結(jié)果發(fā)現(xiàn):與單獨(dú)培養(yǎng)的M2型巨噬細(xì)胞比,同U87MG膠質(zhì)瘤細(xì)胞共培養(yǎng)的M2型巨噬細(xì)胞表達(dá)和分泌VEGF、bFGF以及YKL-40明顯增加。結(jié)果顯示:與U87MG膠質(zhì)瘤細(xì)胞共培養(yǎng)的M2型巨噬細(xì)胞可以高表且與單獨(dú)培養(yǎng)的M2型巨噬細(xì)胞比具有明顯的統(tǒng)計(jì)學(xué)意義。以上這些成血管因子的變化說明膠質(zhì)瘤細(xì)胞可能參與誘導(dǎo)M2型巨噬細(xì)胞大量表達(dá)和分泌VEGF、bFGF以及YKL-40,但具體的機(jī)制還不清楚。當(dāng)阿司匹林濃度達(dá)8mm時(shí),共培養(yǎng)體系中的M2型巨噬細(xì)胞表達(dá)VEGF較空白對(duì)照組的M2型巨噬細(xì)胞明顯減少。因此,如果采用阿司匹林可以起到重要的抗血管新生作用,阿司匹林不但可以直接抑制腫瘤細(xì)胞成血管因子的表達(dá),同時(shí)也可以對(duì)腫瘤微環(huán)境中M2型巨噬細(xì)胞成血管因子的生成起到抑制作用,從而為腫瘤的抗血管治療提供更多可靠依據(jù),也可為阿司匹林的進(jìn)一步藥用價(jià)值開發(fā)帶來新希望。
這項(xiàng)研究初步探索了阿司匹林干預(yù)腦膠質(zhì)細(xì)胞瘤相關(guān)巨噬細(xì)胞表達(dá)VEGF、bFGF和YKL-40成血管生長(zhǎng)因子的影響,關(guān)于阿司匹林干預(yù)血管新生的靶點(diǎn)機(jī)制研究還需要借助基因組學(xué)、代謝組學(xué)等手段深入探索。
