梵凈山東北部同域黔金絲猴與藏酋猴的時空分布格局*
2022-06-15 09:03:26李海波楊朝輝粟海軍胡燦實張明明
林業(yè)科學(xué) 2022年3期
王 丞 李海波 楊朝輝 畢 興 樊 涵 粟海軍 胡燦實 張明明
(1.貴州大學(xué)林學(xué)院 貴州大學(xué)生物多樣性與自然保護研究中心 貴陽 550025; 2.貴州梵凈山國家級自然保護區(qū)管理局 江口 554400; 3.貴州麻陽河國家級自然保護區(qū)管理局 沿河 565300; 4.貴州省興義市林業(yè)局 興義 562400; 5. 湖南張家界大鯢國家級自然保護區(qū)事務(wù)中心 張家界 427400)
明晰野生動物分布特點及影響因素一直是動物空間生態(tài)學(xué)研究的重點內(nèi)容, 這是因為物種往往不是隨機分布的, 而是選擇性地棲息在利于其生存繁衍的適生區(qū), 通常與氣候、地形、食物資源以及生物有機體的競爭、捕食等環(huán)境條件息息相關(guān)(Rosenzweig, 1981; 孫儒泳等, 2019)。由于物種對各類環(huán)境的喜愛度或耐受度不同, 環(huán)境因子的時空分布差異也就制約著物種分布(Ofstadetal., 2019; Kinapetal., 2021)。作為動物行為學(xué)研究的重要內(nèi)容, 日活動節(jié)律是了解野生動物的空間分布與環(huán)境關(guān)系的基礎(chǔ), 可深入了解野生動物的生存策略(Clutton-Brock, 1977)。因此, 生態(tài)環(huán)境條件變化對野生動物行為的影響, 以及它們?nèi)绾卧谶m宜棲息地內(nèi)合理安排活動時間, 并將能量收支成本最優(yōu)化, 對其生存維持具有重要意義。
當(dāng)2個或多個同域生存物種在時空上使用相同資源時, 必然產(chǎn)生種間資源爭奪(Hardin, 1960), 使劣勢物種可獲得資源變少, 甚于可能在特定分布區(qū)內(nèi)被優(yōu)勢物種排除(Wiszetal., 2013; Sanzoetal., 2014)。Schoener(1974; 1986)認為空間、時間或取食等機制差異可促使生態(tài)位分化和細分資源利用, 讓各物種占有獨特的環(huán)境資源, 以減少或避免直接競爭, 保障種群正常活動的同時也有效維護了種間關(guān)系。這既是維持生物多樣性和群落結(jié)構(gòu)穩(wěn)定的基礎(chǔ), 又是反向制約當(dāng)?shù)厣鷳B(tài)系統(tǒng)中各物種消長平衡的關(guān)鍵(Chesson, 2000)。Schreier等(2009)比較了全球同域靈長類的生態(tài)位分化, 發(fā)現(xiàn)其競爭共存方式主要是通過大環(huán)境下的棲息地選擇差異、食性分化以及共占棲息地的垂直空間、微環(huán)境資源或活動時間的選擇差異來實現(xiàn)。其中, 同域靈長類的棲息地選擇和食性分化是生態(tài)位分離、避免競爭、穩(wěn)定共存的最常見行為。同時, 生態(tài)選擇機制的不同也反映了物種的生理生態(tài)需求差異(Grueteretal., 2010; Zhouetal., 2018)。
黔金絲猴(Rhinopithecusbrelichi)僅分布于梵凈山保護區(qū)內(nèi), 種群數(shù)量約700余只(楊業(yè)勤等, 2002)。受當(dāng)?shù)芈糜位顒佑绊? 主要棲息于海拔1 500~1 900 m的常綠落葉闊葉混交林內(nèi)(Xiangetal., 2009; Guoetal., 2018)。廣泛分布于中國中部到南部的藏酋猴(Macacathibetana)(蔣學(xué)龍等, 1996), 環(huán)境適應(yīng)能力較強, 適應(yīng)于多種森林生態(tài)系統(tǒng), 從溫帶山地森林到亞熱帶濕潤常綠森林都有活動(熊成培, 1984; Zhaoetal., 1989), 在梵凈山東北部主要棲息于海拔800~1 900 m的常綠闊葉林和常綠落葉闊葉混林交林內(nèi)(楊炯蠡, 1994)。過去, 黔金絲猴的棲息面積較廣, 于梵凈山的東北部至西南部均見活動(楊業(yè)勤等, 2002), 隨著當(dāng)?shù)芈糜伍_發(fā)建設(shè), 大量游客涉足, 對生性機敏、害羞的黔金絲猴造成了極大影響, 導(dǎo)致其原有的適宜棲息地破碎化, 壓縮了活動空間(Guoetal., 2020)。因此, 明晰二者的時空分布關(guān)系非常重要, 是進一步研究資源分配和競爭模式的必要前提(Rosenzweig, 1981), 對它們的持續(xù)生存和保護管理具有重要意義。
在考慮到不影響和損害靈長類及其他野生動物種群正常活動的情況下, 本研究用紅外相機技術(shù)開展了長時間、全天候、多生境的同步定點監(jiān)測(O’Connelletal., 2010), 研究了梵凈山東北部同域分布的黔金絲猴和藏酋猴的時空共存機制, 以期揭示二者在梵凈山保護區(qū)的時空分布格局, 將有助于了解這2種靈長類活動與環(huán)境的關(guān)系, 并提高對二者種間關(guān)系的認識。

圖1 梵凈山國家級自然保護區(qū)紅外相機布設(shè)方案Fig. 1 The infrared camera locations in Fanjingshan National Nature Reserve表示紅外相機布設(shè)于2017年; 表示紅外相機布設(shè)于2020年。 indicates that the camera was deployed in 2017; indicates that the camera was deployed in 2020.
1 研究地區(qū)概況
梵凈山國家級自然保護區(qū)(以下簡稱“梵凈山保護區(qū)”)位于貴州省東北部, 是武陵山脈的主峰, 地處云貴高原向湘西丘陵過渡的斜坡地帶, 地理位置為27°49′50″—28°1′30″N, 108°45′55″—108°48′30″E, 總面積419.00 km2(圖1)。作為長江上游森林生態(tài)區(qū)生物多樣性保護優(yōu)先區(qū)中具有最高保護優(yōu)先性的區(qū)域之一, 保護區(qū)內(nèi)自然植被保存較為完好, 動植物資源極其豐富, 為多種珍稀瀕危動植物提供了優(yōu)良棲息環(huán)境, 是黔金絲猴野外種群的唯一棲息地(吳波等, 2006)。
梵凈山保護區(qū)屬中亞熱帶季風(fēng)濕潤氣候, 溫暖濕潤, 雨熱同期, 形成了中亞熱帶濕潤區(qū)山地森林生態(tài)系統(tǒng), 年均氣溫變化在-1~21 ℃, 年降水量約1 300 mm(周政賢, 1990)。保護區(qū)地貌復(fù)雜, 山高谷深,高峻陡險,相對高差達2 000 m, 海拔范圍500~2 570 m。主要植被類型有: 1)低山次生的落葉闊葉林和針闊混交林, 受人為干擾較重, 有明顯次生特點, 主要分布于海拔1 300 m以下, 多是在原有地帶性植被消失后出現(xiàn)次生植物群落, 主要群落類型包括楓楊(Pterocaryastenoptera)林、赤楊葉(Alniphyllumfortunei)林和楓香(Liquidambarformosana)林等, 甚至存在一小部分人工針葉純林, 主要群落類型有杉木(Cunninghamialanceolata)林、馬尾松(Pinusmassoniana)林等; 2)常綠闊葉林, 為該地區(qū)典型的原生植被群落, 主要分布于海拔1 400 m以下, 群落類型有栲樹(Castanopsisfargesii)林、米櫧(Castanopsiscarlesii)林等; 3)中山常綠落葉闊葉混交林, 主要分布于海拔1 300~1 900 m, 群落類型有光葉水青岡(Faguslucida)林、貴州青岡(Cyclobalanopsisargyrotricha)林等; 4)高山矮林灌叢和針葉林, 主要分布于海拔1 800 m以上, 主要群落類型包括高山柏(Sabinasquamata)灌叢、大花黃楊(Buxushenryi)灌叢、鐵杉(Tsugachinensis)林、黃山松(Pinustaiwanensis)林等(周政賢, 1990; 楊海龍等, 2010)。
2 研究方法
2.1 紅外相機布設(shè)方案
基于梵凈山保護區(qū)的矢量邊界 進行1 km2的網(wǎng)格化, 紅外相機密度為1臺· km-2。根據(jù)梵凈山保護區(qū)黔金絲猴與藏酋猴的歷史分布研究, 主要選擇在黔金絲猴的主要分布區(qū)及與藏酋猴同域的梵凈山東北部布設(shè)相機(圖1)。在紅外相機實際布設(shè)中, 監(jiān)測點選取綜合考慮了覆蓋主要生境和海拔、通行可達性等因素, 具體布設(shè)于有野生動物活動痕跡的林間開闊地。2017年預(yù)先布設(shè)41臺, 2020年增加25臺, 共計布設(shè)紅外相機66臺, 整個期間連續(xù)監(jiān)測。
紅外相機(獵科LTL-6210 MC、東方紅鷹E1B)設(shè)置為拍照模式、連拍3張、時間間隔10 s、中靈敏度, 固定于離地高0.5~2.0 m的樹干, 鏡頭盡量與地面平行, 避開陽光直射。同時, 記錄相機的GPS位點、海拔、林型以及水源距離等基本生境信息。
2.2 數(shù)據(jù)處理
首先, 挑出所有可識別黔金絲猴與藏酋猴的照片, 將在30 min內(nèi)拍攝到單種靈長類的所有照片定義為1次獨立有效照片, 并記為1次有效日活動時間(O’Brienetal., 2003)。由于紅外相機監(jiān)測時間跨度大、監(jiān)測時長不均一, 且未嚴格按照靈長類家庭群體的家域進行相機布設(shè)。因此, 在評估二者的種群空間分布和重疊情況時, 未考慮不同家庭群的相互競爭與結(jié)構(gòu)組成, 以分布于研究區(qū)域內(nèi)的生存種群作為評估單元。同時, 考慮到時間跨度影響, 使用相對豐富度指數(shù)(relative abundance index, RAI)作為種群分布的評估指標(biāo)(李晟等, 2016)。其原理是以同一物種單位時間內(nèi)所獲有效照片數(shù)為基礎(chǔ), 即拍攝率, 計算方法為:
RAI =Ai/N× 1 000。
其中,Ai為紅外相機所拍攝到的第i個動物物種的獨立照片數(shù),N為Ai位點的總相機日, 單臺紅外相機持續(xù)工作24 h定義為1個相機日。
對存在空間重疊的相機位點, 使用“空間相遇時間差”來評估二者在同一空間的相遇時間變化情況。“空間相遇時間差”被定義為, 在同一相機位點上乙物種(黔金絲猴)在某一時刻出現(xiàn)后, 在此時刻前后出現(xiàn)的甲物種(藏酋猴)與乙物種(黔金絲猴)的時間差值,即 甲物種出現(xiàn)時刻與乙物種出現(xiàn)時刻之差。空間相遇時間差為正值, 表示甲物種出現(xiàn)晚于乙物種, 反之負值表示甲物種出現(xiàn)早于乙物種。
在以上各項數(shù)值的統(tǒng)計分析中, 對非參數(shù)的2個獨立樣本的差異使用Mann-Whitney U檢驗進行比較; 對非參數(shù)的多個獨立樣本的差異采用Kruskal-Wallis H檢驗進行比較; 對連續(xù)變量使用線性回歸進行相關(guān)性分析。差異顯著水平為P= 0.05。
利用整個監(jiān)測期間的數(shù)據(jù)評估日活動節(jié)律, 以獲悉監(jiān)測區(qū)域內(nèi)黔金絲猴與藏酋猴的整體日活動水平及其季節(jié)性變化。日活動節(jié)律描述方法使用核密度估計(Kernel density estimation)(Ridoutetal., 2009), 方法是將1天視為1個24 h的圓循環(huán), 每次有效活動時間作為連續(xù)時間循環(huán)中的隨機樣本, 描述一種靈長類在某個特定時間段內(nèi)被發(fā)現(xiàn)的概率, 以時間為橫軸, 以該時間點上物種被發(fā)現(xiàn)的概率為縱軸, 最終呈現(xiàn)該物種在某個特定時間點活動的概率曲面圖。同時, 所構(gòu)成的曲線下的面積積分值為1。
為比較2種靈長類的日活動節(jié)律差異, 采用重疊指數(shù)比較兩者核密度曲線之間的相似程度, 以估計二者的日活動節(jié)律對稱重疊程度(Δ), 其取值范圍從0(無重疊)到1(完全重疊)(Ridoutetal., 2009)。同時, 利用Wald檢驗比較2個物種的日活動節(jié)律差異, 差異顯著水平為P= 0.05。以上所有處理結(jié)果在R軟件v4.0.2的“activity”和“overlap”包中進行。
3 結(jié)果與分析
3.1 空間分布
在近4年的紅外相機連續(xù)監(jiān)測中, 累計有效工作日43 028天, 共記錄到靈長類照片7 613張, 獨立有效照片1 417張, 其中藏酋猴1 049張、黔金絲猴368張。在66個監(jiān)測相機位點中, 藏酋猴在65個位點被記錄到, 分布占比達到98.48%, 可見藏酋猴在研究區(qū)域內(nèi)普遍分布, 從監(jiān)測位點的最低海拔700 m至最高海拔2 400 m的有林地均有活動; 黔金絲猴在49個位點被記錄到, 分布占比達74.24%, 主要分布于研究區(qū)域中北部, 且不分布于索道附近(圖2)。可見, 黔金絲猴的分布相對較窄, 完全重疊于藏酋猴的分布范圍內(nèi), 且二者在中北部的多個位點空間交互明顯。

圖2 梵凈山黔金絲猴與藏酋猴的空間分布Fig. 2 Spatial distribution of Rhinopithecus brelichi and Macaca thibetana in Fanjingshan Reserve
藏酋猴分布的海拔差異不顯著(Kruskal-Wallis test,χ2= 5.22, df = 4,P= 0.27), 其RAI與海拔不具線性關(guān)系(F= 0.51,P= 0.48), 表明其適應(yīng)多段海拔; 黔金絲猴分布的海拔差異極顯著(Kruskal-Wallis test,χ2= 17.78, df = 4,P= 0.001), 其RAI與海拔呈線性相關(guān)(F= 7.19,P= 0.009), 在1 600~1 900 m相對豐富度最高, 表明其更喜歡在該海拔范圍棲息(圖3A)。比較黔金絲猴與藏酋猴在不同海拔段的相對豐富度后發(fā)現(xiàn), 在1 300 m以下(P< 0.01)和1 900 m以上(P< 0.01)的2個海拔范圍呈極顯著差異, 在1 300~1 900 m海拔范圍未有顯著差異。
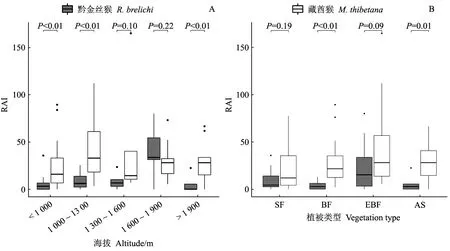
圖3 梵凈山黔金絲猴與藏酋猴在不同海拔與植被類型的RAI值Fig. 3 RAI of R. brelichi and M. thibetana at different altitudes and vegetation types in Fanjingshan ReserveSF:低山次生林;EF:常綠闊葉林;EBF:常綠落葉闊葉混交林;AS:高山矮林灌叢。SF: Secondary forest; EF: Broad-leaved evergreen forests; EBF: Evergreen and deciduous broad-leaved forests; AS: Alpine shrub.
藏酋猴分布的植被類型差異不顯著(Kruskal-Wallis test,χ2= 3.63, df = 3,P= 0.30), 說明其適應(yīng)多種森林植被類型; 黔金絲猴分布的植被類型差異極顯著(Kruskal-Wallis test,χ2= 12.65, df = 3,P= 0.005), 說明其更喜歡出現(xiàn)于中山常綠落葉闊葉林, 較少出現(xiàn)于其余3種森林植被類型(圖3B)。比較黔金絲猴與藏酋猴在不同植被類型中的相對豐富度后發(fā)現(xiàn), 在常綠闊葉林(P< 0.01)和高山矮林灌叢中(P= 0.01), 二者分布呈顯著差異; 在低山次生林(P= 0.19)和中山常綠闊葉林中(P= 0.09), 二者分布未有顯著差異, 二者均較少出現(xiàn)于低山次生林。

圖4 梵凈山黔金絲猴與藏酋猴在同一位點的相遇時間差Fig. 4 Encounter time difference betweenR. brelichi and M. thibetana at the same camera site in Fanjingshan Reserve
3.2 同一空間的相遇時間
統(tǒng)計黔金絲猴與藏酋猴的共現(xiàn)位點后發(fā)現(xiàn), 二者15天內(nèi)相遇的位點有45個, 占總監(jiān)測位點數(shù)的68.18%, 占黔金絲猴分布總位點數(shù)的91.84%, 表明二者的棲息空間重疊度極高。對同一棲息地的利用時間差表現(xiàn)為, 于黔金絲猴出現(xiàn)的前后, 藏酋猴出現(xiàn)在同一位點的時間不存在顯著差異(P= 0.48), 平均5~6天會出現(xiàn)在同一位點, 表明二者對同一棲息地的利用有一定的時間分布規(guī)律(圖4)。四季變化對黔金絲猴與藏酋猴在同一棲息地的相遇時間無顯著影響(Kruskal-Wallis test,χ2= 3.31, df = 3,P= 0.35), 平均相遇時間的變化范圍為5天(冬季)至7天(春季)(圖5)。

圖5 梵凈山黔金絲猴與藏酋猴在不同季節(jié)的相遇時間差Fig. 5 Encounter time difference betweenR. brelichi and M. thibetana in different seasons in Fanjingshan Reserve
3.3 日活動節(jié)律
黔金絲猴與藏酋猴為完全晝行性物種, 但二者明顯擁有不同的日活動節(jié)律, 全年數(shù)據(jù)分析結(jié)果呈現(xiàn)極顯著差異(P< 0.01), 重疊度為0.76(圖6)。黔金絲猴呈現(xiàn)晨昏型的日活動節(jié)律, 呈雙峰型, 一天中活動時間跨度約13 h, 活動高峰期在晨間7: 00—10: 00和黃昏16: 00—18: 00時段, 日中活動強度較低, 活動強度最低時段在13: 00—15: 00; 藏酋猴日活動節(jié)律呈單峰型, 一天中活動時間跨度約14 h, 日出后活動逐漸增強, 高峰在9: 00—12: 00, 午后逐漸減弱。
比較黔金絲猴與藏酋猴在不同季節(jié)的日活動節(jié)律后發(fā)現(xiàn), 二者均呈現(xiàn)季節(jié)性變化, 重疊程度不同[冬季(0.64)<春季(0.68)<夏季(0.75)<秋季(0.82)],二者的季節(jié)性日活動節(jié)律均呈極顯著差異(P< 0.01)。春季, 二者均呈現(xiàn)雙峰型, 其中黔金絲猴的2個活動高峰最明顯, 與藏酋猴的活動高峰時段形成錯峰現(xiàn)象(圖7A)。夏季, 黔金絲猴呈多峰型, 藏酋猴的呈單峰型(圖7B), 其中黔金絲猴增加了午間前后的活動強度, 使日活動時間分配更均勻; 藏酋猴從日出后活動強度逐漸增加, 于10: 00—12: 00出現(xiàn)活動高峰, 之后活動強度逐漸下降直至天黑。秋季, 黔金絲猴呈雙峰型, 藏酋猴呈單峰型(圖7C)。冬季與春季相似, 二者形成錯峰現(xiàn)象(圖7D), 其中, 黔金絲猴2個活動高峰在8: 00—10: 00和16: 00—18: 00; 藏酋猴從日出開始活動強度逐漸增加, 12: 00—14: 00達到高峰, 之后迅速下降。

圖6 梵凈山黔金絲猴與藏酋猴的日活動節(jié)律Fig. 6 Daily activity patterns of R. brelichi and M. thibetana in Fanjingshan Reserve
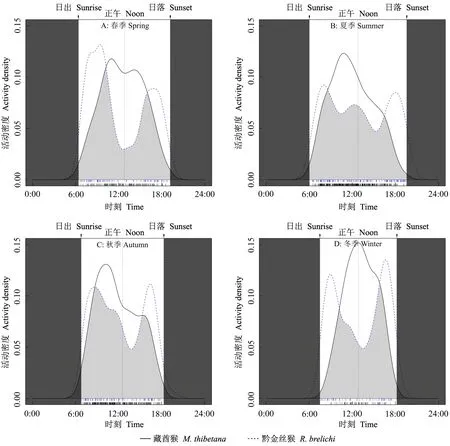
圖7 梵凈山黔金絲猴與藏酋猴的日活動節(jié)律及其季節(jié)變化Fig. 7 Daily activity patterns and seasonal variation of R. brelichi and M. thibetana in Fanjingshan Reserve
4 討論
4.1 時間關(guān)系
黔金絲猴與藏酋猴的日活動節(jié)律明顯不同, 呈現(xiàn)極顯著差異(P< 0.01), 很好地錯開了活動時間的競爭。雖然存在日活動節(jié)律重疊度的季節(jié)變化(冬春夏秋依次增大), 秋季的高重疊可能會引起競爭, 但筆者認為這種日活動節(jié)律差異和季節(jié)變化更多來自內(nèi)源適應(yīng), 與自身取食特征和對棲息環(huán)境的溫、光和食物資源等變化的響應(yīng)密切相關(guān)(尚玉昌, 2014)。此外, 秋季的棲息地內(nèi)分布著大量優(yōu)質(zhì)果實資源, 可選食物豐富, 同域靈長類對特定空間的競爭壓力相對減少, 可通過取食不同主要食物維護共存(Singhetal., 2011; Zhouetal., 2018)。因而, 高度相似的日活動節(jié)律變得不再重要, 更多地在于如何適應(yīng)該環(huán)境條件的季節(jié)變化, 使其日活動時間得到合理有效的分配(Schoener, 1974)。
受地球公轉(zhuǎn)影響, 滇金絲猴(R.bieti)的日活動時間在短日照的冬季比長日照的夏季縮短2 h, 且推遲1 h開始活動(Dingetal., 2004)。其他晝行性靈長類也表現(xiàn)出相似行為(黃中豪等, 2007; Hanyaetal., 2018)。Dunbar(1992)認為, 晝長是有效制約晝行性動物日活動時間分配和長度的重要因素, 如何在晝間獲得一天內(nèi)所需能量, 合理分配其活動時間尤為重要。黔金絲猴與藏酋猴在不同季節(jié)對晝長的反應(yīng)相似, 相較于夏季, 在冬季呈現(xiàn)為日活動時間縮短和日出活動時間推遲。可見, 黔金絲猴與藏酋猴的日活動時長同樣受晝長制約。
黔金絲猴與藏酋猴的日活動節(jié)律均為完全晝行性, 但黔金絲猴活動呈明顯晨昏性, 藏酋猴則呈單高峰特點。Clutton-Brock(1974;1977)認為引起靈長類日活動節(jié)律不同的重要因素是白天溫度和日照強度的變化, 以及與自身的消化生理和形態(tài)特征, 并最終表現(xiàn)出一定的節(jié)律性適應(yīng)特征。黔金絲猴與葉食性靈長類的日活動節(jié)律相似(Huangetal., 2003; 黃乘明等, 2018; 張龍等, 2019), 即在經(jīng)過夜間較長時間的不進食后, 急需補充營養(yǎng), 會隨日出開始積極尋找和采集食物, 出現(xiàn)第1個活動高峰; 午間普遍會增加休息減少活動, 因為所食樹葉富含纖維素, 休息利于樹葉營養(yǎng)在腸胃中的發(fā)酵、分解和吸收(美國國家學(xué)院國家研究委員會, 2011); 為迎接漫漫長夜的到來, 日落前普遍又再一次進行高強度取食, 以保障進入夜宿地后的熱量維持。藏酋猴單峰型日活動節(jié)律與獼猴類相似(周岐海等, 2007; Hanyaetal., 2018), 因其腸道短、食性雜, 多取食高糖、高水、低纖維的食物(美國國家學(xué)院國家研究委員會, 2011), 大多沒有明顯晨昏活動特點, 而是在日出后活動逐漸增強, 日間持續(xù)保持一定強度的活動, 常在一定時段出現(xiàn)明顯單高峰或不明顯的多高峰。
季節(jié)性的溫度變化是影響靈長類日活動節(jié)律季節(jié)差異的重要因素, 其帶來的環(huán)境資源變化迫使靈長類改變生存適應(yīng)策略, 尤其在嚴寒冬季和酷熱夏季表現(xiàn)更明顯(Nakagawaetal., 2010; 黃乘明等, 2018)。冬季面臨低溫、食物匱乏的困境, 靈長類生存壓力劇增, 為減少機體損耗、高效獲取每日所需基本營養(yǎng), 它們常通過節(jié)能保溫的生存策略度過寒冬(黎大勇, 2010; Nakagawaetal., 2010; 黃乘明等, 2018)。藏酋猴與黔金絲猴為躲避冬季寒冷, 會傾向于選擇高溫時段活動(圖7D), 藏酋猴表現(xiàn)為日活動開始時間變晚, 活動高峰由上午推遲至午間。這種躲避低溫的越冬策略普遍存在于多種靈長類中, 如日本獼猴(M.fuscata)在冬季偏向于午后至下午活動(Hanyaetal., 2018), 熊猴(M.assamensis)的覓食、移動活動于11: 00后保持著較高比例, 并一直持續(xù)到日落前(周岐海等, 2007), 黑葉猴(Trachypithecusfrancoisi)的大部分覓食、移動活動也分配于下午時段(Zhouetal., 2007)。這歸因于日間的積溫變化, 日間高溫時段常集聚在中午到下午, 因而在評估低溫能量損耗與覓食利益之后, 靈長類可能采取能量最大化的活動策略, 以減少不必要的能量損耗(Schoener, 1971)。黔金絲猴表現(xiàn)為晨、昏某時段活動的高度聚集, 以及午間進行長時間休息, 目的是保持體溫和對上午獲得食物營養(yǎng)的較高效率的發(fā)酵、分解和吸收(Huangetal., 2003; Zhouetal., 2007; 黎大勇, 2010)。夏季大多數(shù)靈長類會在林下或洞穴中休息納涼減少活動, 以躲避正午至午后時段的高溫侵襲(Dingetal., 2004; 黃中豪等, 2007; 黃乘明等, 2018)。為躲避高溫, 藏酋猴日活動集中于上午, 午后活動強度逐漸減弱。然而, 黔金絲猴日活動節(jié)律呈現(xiàn)多峰型特點, 增強了午間活動, 筆者認為這種日活動策略可能與食物資源的豐度與種類有關(guān), 夏季豐富的花、果給它們提供了高質(zhì)量、易消化的食物, 減少了休息消化時間, 增加了活動時間的寬度與自由度(Schoener, 1971; Dingetal., 2004)。另外, 這還可能與安裝的相機位置有關(guān), 相較于樹冠層或裸露區(qū), 林下溫度明顯更低、更涼爽(Huangetal., 2003), 使得黔金絲猴在氣溫高的時段更易在林下被發(fā)現(xiàn)。總的來說, 它們的日活動差異主要與當(dāng)?shù)貧夂颦h(huán)境和食物資源有關(guān), 普遍呈現(xiàn)既適應(yīng)當(dāng)?shù)丨h(huán)境又滿足自身需求的變化(Nakagawaetal., 2010; 黎大勇等, 2013; 黃乘明等, 2018)。
4.2 空間關(guān)系
黔金絲猴的空間分布明顯嵌套于藏酋猴分布范圍內(nèi), 且不出現(xiàn)于風(fēng)景旅游區(qū)。過去, 旅游風(fēng)景區(qū)部分范圍曾是黔金絲猴由梵凈山東北部向西南部游走的途經(jīng)區(qū)域(楊業(yè)勤等, 2002), 但隨當(dāng)?shù)芈糜伍_發(fā), 生性機敏、害羞的黔金絲猴活動空間受到極大的負面影響。Guo等(2020)經(jīng)過2年實地踏查, 發(fā)現(xiàn)梵凈山南部未見黔金絲猴任何活動痕跡。同時, 筆者的研究也表明黔金絲猴不再出現(xiàn)于梵凈山風(fēng)景旅游區(qū)范圍, 意味著它們的活動路線可能改變或遷移受阻。因此, 相比于原有棲息活動范圍, 環(huán)境現(xiàn)狀可能壓縮了黔金絲猴的分布空間或造成分布破碎化。進一步還可能影響種群間交流, 威脅到種群遺傳繁衍: 在梵凈山東北部獲取的128只野外個體線粒體DNA變異分析研究中, 發(fā)現(xiàn)黔金絲猴遺傳多樣性已處于極低狀態(tài), 種內(nèi)受脅程度緊迫(Panetal., 2011)。另外, 由于藏酋猴在梵凈山保護區(qū)廣泛分布且環(huán)境適應(yīng)能力較強, 種群生存更具優(yōu)勢, 可能也會阻礙黔金絲猴的種群發(fā)展, 增加黔金絲猴的生存阻力。
雖然筆者發(fā)現(xiàn)黔金絲猴與藏酋猴的空間分布高度重疊, 但二者不會同框出現(xiàn), 存在一定規(guī)律的利用時間差異, 交互利用時間相差平均5~6天, 有效降低在同一空間相遇的可能競爭。同時, 黔金絲猴與藏酋猴棲息地選擇明顯不同, 黔金絲猴主要活動于1 600~1 900 m的中山常綠落葉闊葉混交林中(圖3), 這一特定活動范圍與其他學(xué)者研究結(jié)果一致, 主要原因在于其主要食物來源于該區(qū)域的森林, 以及遠離低海拔的人類活動干擾(Xiangetal., 2012; Guoetal., 2020)。藏酋猴則廣泛分布于整個調(diào)查區(qū)域, 僅在植被較單一的次生林中活動較低(圖3B), 與在其他分布區(qū)一樣, 表現(xiàn)出極強的環(huán)境適應(yīng)能力(蔣學(xué)龍等, 1996; 熊成培, 1984; Zhaoetal., 1989)。因此, 空間上合理有效的劃分自需資源和利用時間差, 無疑減小了二者的空間相遇機會和可能競爭。在其他地區(qū), 與本研究親緣關(guān)系類似的同域靈長類也呈現(xiàn)明顯的棲息地選擇差異, 如云南白馬雪山自然保護區(qū)的獼猴(M.mulatta)與滇金絲猴(R.bieti), 前者更喜歡中低海拔的常綠闊葉林, 后者更喜歡中高海拔的常綠落葉闊葉混交林(Grueteretal., 2010); 分布于四川唐家河自然保護區(qū)的藏酋猴與川金絲猴(R.roxellanae), 前者主要棲息于海拔1 200~1 800 m的常綠落闊混交林和落葉闊葉林內(nèi), 后者主要在海拔1 800~2 900 m的落葉闊葉林和針闊混交林內(nèi)(白洢露, 2019)。Grueter等(2010)認為同域物種的生態(tài)選擇差異一定程度上反映了種間競爭的存在與分化, 也預(yù)示著它們不同的生理生態(tài)需求。經(jīng)過歷史的長期適應(yīng)進化, 不同靈長類的腸胃消化系統(tǒng)各有差異, 意味著它們的食物選擇存在一定變化(美國國家學(xué)院國家研究委員會, 2011)。藏酋猴相比于黔金絲猴, 它們胃內(nèi)壁更簡單、光滑, 小腸更短, 不易分解纖維含量較高的葉組織(美國國家學(xué)院國家研究委員會, 2011; Zhouetal., 2014), 藏酋猴更多以高蛋白、高糖分和低纖維的植物組織為食(Zhaoetal., 1991)。然而, 黔金絲猴擁有作為食物初級儲存和發(fā)酵的前囊, 類似于反芻動物的前胃結(jié)構(gòu), 可通過發(fā)酵的方式降解葉片纖維素, 能有效地吸收和利用其中的營養(yǎng), 最終造就黔金絲猴更多以葉為食(Zhouetal., 2014)。因此, 對于生態(tài)位劃分明顯的同域物種, 相互之間是可以友好共存的。
在梵凈山的物種RAI中, 藏酋猴(28.23)占比最高, 甚至超過了野豬(Susscrofa)(11.82)2倍之多(張明明等, 2019)。可見, 性膽大、適應(yīng)能力強的藏酋猴在梵凈山較為常見, 受旅游和人類影響程度很低, 種群優(yōu)勢明顯, 據(jù)楊業(yè)勤等(2002)統(tǒng)計, 梵凈山藏酋猴的種群數(shù)量達2 000余只, 是當(dāng)時黔金絲猴數(shù)量的近3倍。迫于旅游和人類活動影響, 黔金絲猴的空間分布明顯受限(圖2)(Guoetal., 2020), 且近20年來種群數(shù)量一直維持著700余只(楊業(yè)勤等, 2002; 郭艷清等, 2017), 生存現(xiàn)狀岌岌可危。雖然二者擁有不同的取食特性和空間選擇, 但在種群分布、數(shù)量和環(huán)境適應(yīng)能力等方面, 黔金絲猴受限因素明顯更多。隨著藏酋猴數(shù)量不斷擴增和發(fā)展, 處于受限空間內(nèi)的黔金絲猴與藏酋猴的種間關(guān)系可能會惡化, 可能出現(xiàn)類似于西黑冠長臂猿(Nomascusconcolor)與印支灰葉猴(T.crepusculus)不對稱的惡性競爭(Chenetal., 2020), 最終不利于黔金絲猴小種群復(fù)壯和擴增。因此, 為了解二者種間關(guān)系的現(xiàn)狀和未來變化, 還需長期深入調(diào)查, 從而較為全面地獲悉在人類活動和旅游共同影響背景下的這2個物種的競爭與共存關(guān)系。
5 結(jié)論
近4年的紅外相機監(jiān)測研究表明, 黔金絲猴日間活動為晨昏型, 在梵凈山東北部主要分布于遠離旅游索道、海拔1 600~1 900 m范圍的常綠落葉闊葉林中。藏酋猴日間活動為單峰型, 在梵凈山東北部具較強適應(yīng)能力, 調(diào)查范圍內(nèi)均有分布, 無明顯的海拔、植被類型選擇偏好。時空關(guān)系上二者的生態(tài)位劃分明顯, 有效避免了競爭和沖突的發(fā)生, 可友好共存。但迫于人類活動干擾影響, 黔金絲猴的空間分布明顯受限, 不利于其種群發(fā)展, 對現(xiàn)環(huán)境下的二者關(guān)系的發(fā)展趨勢還需要長期深入調(diào)查, 才能較全面獲悉在人類活動干擾背景下的競爭與共存關(guān)系。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18
少先隊活動(2022年5期)2022-06-06 03:45:04
音樂探索(2022年2期)2022-05-30 21:01:37
家庭科學(xué)·新健康(2022年3期)2022-05-10 00:32:13
中老年保健(2021年2期)2021-08-22 07:31:10
小天使·一年級語數(shù)英綜合(2019年8期)2019-08-27 02:23:00
科學(xué)大眾(中學(xué))(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
小學(xué)科學(xué)(學(xué)生版)(2018年7期)2018-08-13 09:33:04
海峽姐妹(2018年3期)2018-05-09 08:20:40
