新疆野生阿魏菇原生質體的再生與單核菌株交配型測定
2022-06-15 02:24:48努爾孜亞亞力買買提郝敬喆祁顥萱裴龍英孫春花賈文捷賈培松
新疆農業科學 2022年5期
努爾孜亞·亞力買買提,羅 影,郝敬喆,祁顥萱,裴龍英,孫春花,賈文捷,賈培松
(1. 新疆農業科學院植物保護研究所/農業部西北荒漠綠洲作物有害生物綜合治理重點實驗室,烏魯木齊 830091;2. 新疆農業大學,烏魯木齊 830091;3. 新疆理工學院,新疆阿克蘇 843000;4.青河縣農業技術推廣中心,新疆阿勒泰 836200)
0 引 言
【研究意義】阿魏菇(Pleurotusferulae) 具有極高的食藥用價值,是一種有良好市場前景的野生食用菌[1]。我國阿魏菇野生資源僅分布在新疆北疆地區的戈壁阿魏灘上,生長在阿魏屬植物根莖部,分布量稀少[2-3]。近些年阿魏屬植物分布面積極度萎縮[4-5],以阿魏屬植物為營養的野生阿魏菇,亟待保護[6-7]。目前,阿魏菇雖已大量栽培,但是由于國內人工栽培時間較久,商品阿魏菇存在種性不穩定、菌種退化等問題[8],加強對阿魏菇新品種進行選育、種質資源的保護迫在眉睫。【前人研究進展】當前核心問題是怎樣提純復壯菌種[9]。優化培養基法、出菇組織再分離法是傳統的菌種提純復壯方式。其中,優化培養基法操作簡單、耗費時間短,但效果不明顯;出菇組織再分離法效果好,但周期長、過程復雜。然而原生質體單核化技術提純復壯菌絲比上述方式效果更好。原生質體技術是現代農業生物技術的一個重要分支[10-11],原慧等[12]研發出新疆野生阿魏菇(P.ferulae)的原生質體高效分離與再生工藝,所得阿魏菇原生質體恢復能力較強;樊曉琳等[13]通過該技術成功地對雙孢蘑菇As2796實現復壯;譚琦等[14]將香菇野生種7號、栽培種Le l當做親本,通過此項技術對申香8號展開選育;吳小平等[15]同樣利用改技術獲得了24個優良的靈芝菌種。江玉姬等[16]證實,原生質體單核化技術是對白色金針菇品種具改良作用的新工藝。交配型測定是控制菌種單孢雜交或單雙雜交的交配、結實等關鍵遺傳過程的決定步驟,2個異宗聯合的菌株間交配因子的異同差異,是判別2個菌株親緣關系遠近和遺傳距離的重要指標[10]。拮抗現象是反映菌株間遺傳差異以及同緣關系的主要性狀[17-18],是物種之間為保持種群遺傳物質穩定而限制外源基因侵入的本能反應,該現象在自然界普遍存在以防止不同個體間的融合[19]。【本研究切入點】雖然食用菌育種領域已普遍應用原生質體單核化技術,但是,目前應用原生質體單核化技術對新疆野生阿魏菇菌種進行提純復壯還缺乏系統深入的研究。需研究利用原生質體單核化技術獲得具有優良性狀的阿魏菇(Pleurotusferulae)新菌種。【擬解決的關鍵問題】研究對原生質體技術獲得的阿魏菇單核菌株進行交配型測定,分析阿魏菇原生質體單核菌株與親本菌株在生長性狀方面的差異,篩選阿魏菇雜交菌株并進行拮抗驗證,獲得性狀優良的阿魏菇新菌種,為新疆野生阿魏菇新品種的選育及產業化奠定理論與技術基礎。
1 材料與方法
1.1 材 料
供試的3個親本野生阿魏菇菌株PL001、PL176、PL163均由新疆農業科學院植物保護研究所食用菌實驗室提供。
1.2 方 法
1.2.1 供試試劑與培養基
2%Lywallzyme(即溶壁酶,取自廣東微生物研究所):2%溶壁酶中加入0.6 M無菌甘露醇(mannitol)。
PDA 固體培養基:馬鈴薯 200 g,葡萄糖20 g,瓊脂20 g;
PDA 液體培養基:馬鈴薯 200 g,葡萄糖15 g;
MYG 再生培養基:麥芽糖 10 g,葡萄糖4 g,酵母膏4 g,0.6 M甘露醇,瓊脂粉20 g。。
以上培養基皆用水定容到1 000 mL,酸堿度自然,于121℃下,行20 min滅菌處理,再待用。
1.2.2 原生質體制備與再生
將供試菌株分別接種至PDA培養基平板純化,生長7 d后挑取5×5 (mm)菌塊接種至PDA培養基中央,在25℃、暗環境中,實施7 d培養。將新生菌絲挑至PDA液體培養基內,25℃靜止培養 7 d,待出現菌絲球后利用篩網過濾菌絲,分別用無菌水、0.6 M甘露醇各沖洗1次,用濾紙吸干水分,放入10 mL離心管中,添加0.6 M甘露醇3 mL,當溫度為4℃,轉速為3 000 r/min時,5 min離心處理,棄上清液,將沉淀物加入2%溶壁酶3 mL,30℃水浴1.5~4.5 h,間隔30 min輕輕搖動離心管,鏡檢菌絲破壁情況。待酶解停止,將菌絲體混合液移至無菌且配備棉塞的注射器,經過濾將斷裂菌絲清理掉,在3 000 r/min速度下,對濾液實施10 min離心,吸取上清,同時將沉淀收集起來,通過0.6 M甘露醇稀釋。借助移液槍將0.1 mL的原生質體稀釋液均勻涂布在再生培養基內,于25℃下暗培養,得到原生質體再生菌落。
1.2.3 單核菌株獲得
利用單孢分離方法[20],在顯微鏡下挑選菌落小、生長緩慢的單菌落接種至PDA固體斜面培養基上,等到菌落長到1 cm,顯微鏡下觀察是否存在鎖狀聯合,無鎖裝聯合則為單核菌株。
1.2.4 菌株交配型測定及單核菌株篩選
選取3株單核菌株為測試菌株,分別與其它單核菌株進行配對,接種于PDA平板上并于25℃避光培養。7 d后顯微觀察菌絲鎖狀聯合有無,再根據交配反應結果進行分析。確定交配類型后,根據各個單核菌株的生長速度進行篩選,以每個交配型中生長速率最快的作為育種材料。
1.2.5 雜交菌株獲得
將不同親本菌株中篩選出的不同交配型的單核菌株進行兩兩配對,在PDA固體培養基上進行雜交。挑取有鎖狀聯合的菌絲接種到 PDA平板上培養2~3 d,挑取新鮮的菌絲進行純化,獲得雜交菌株。
1.3 數據處理
應用Excel 2010、SPSS 19.0軟件對數據進行處理和方差分析(AVNOA)。
2 結果與分析
2.1 酶解時間對原生質體產量的影響
研究表明,供試菌株PL001、PL163和PL176在酶解1.5 h時均開始釋放原生質體,此時產生的原生質體較大,40×單視野中個數為(3.1~4.4)×105,數量較少;2.5 h之后,觀察到體積較小的原生質體,此時單視野中原生質體個數為(5.9~11.6)×105,隨著酶解時間的延長,原生質體釋放量越來越大。當酶解3.5 h時,40×目鏡單視野中原生質體個數為(10.8~20.6)×105,原生質體釋放量達到最大。當酶解時間延長至4.5 h,原生質體的再生數量開始降低,40×單視野中原生質體個數為(2.9~10.3)×105,隨著酶解時間的延長,原生質體釋放量越來越低。阿魏菇菌絲釋放原生質體數量隨時間延長呈先升高后降低的趨勢,阿魏菇原生質體的酶解最佳時間為3.5 h。圖1

圖1 阿魏菇菌株釋放出原生質體Fig.1 The protoplasts released by Pleurotus ferulae
2.2 原生質體再生與單核菌株獲得情況
研究表明,獲得了462個原生質體萌發單菌落,獲得單核菌株50個。從 PL01菌株原生質體中分離獲得156株單個原生質體再生菌株,其中20株無鎖狀聯合,原生質體單核化率為12.8%;從 PL163菌株原質體中分離獲得74 株單個原生質體再生菌株,其中20株無鎖狀聯合,原生質體單核化率為27.02% ;從PL176 菌株原生質體中分離獲得175株單個原生質體再生菌株,其中13株無鎖狀聯合,原生質體單核化率為12.03%。表1

表1 原生質體單核菌株的數量以及單核獲得比例Table1 The quantity of monocytic strains and the rate of monokaryogenesis
2.3 單核菌株交配型確定及單核菌株篩選
研究表明,在所得2類交配型單核菌株的占比方面,同一親本菌株存在區別,PL01、PL163這2個親本菌株的2類交配型的占比皆是1∶3, PL176此親本菌株的2類交配型占比是1∶4。表2

表2 交配型分類結果Table 2 The classification of mating type
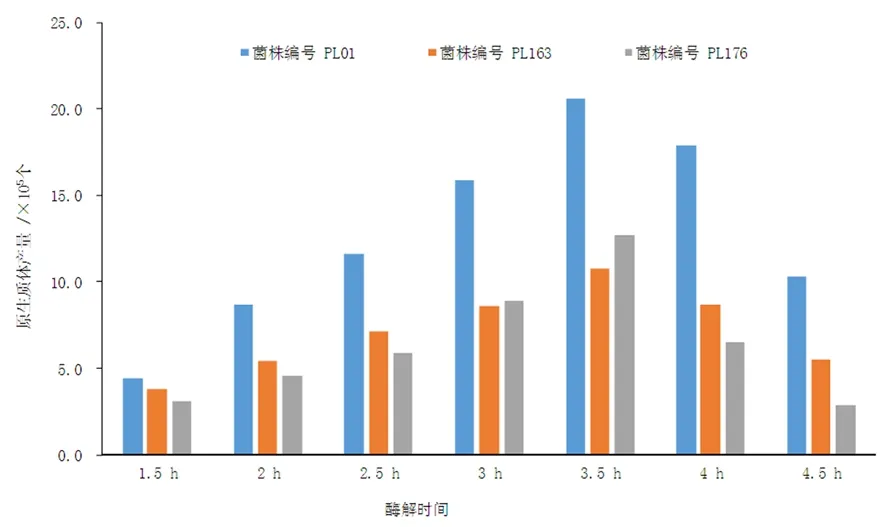
圖2 原生質體產量隨不同酶解時間的變化 (個 /mL)
Fig.2 The production of Ganoderma lucidum protoplast in different time(ind /mL)
不同交配型單核菌株的菌落形態及生長速度差異大,親本菌株生長速度快于單核菌株,親本PL01、PL163和PL176日長速分別為5.28、6.20和5.99 cm。在長速上,不同單核菌株之間的生長速度差異較大,根據單核菌絲的生長速度,每種交配型選擇單核菌株 PL01-P16、PL01-P49、PL163-7、PL163-24、PL176-23、PL176-39作為雜交育種材料。表3,圖4

注:A:單核菌絲;B:雙核菌絲,可見鎖狀聯合

圖4 親本菌株與單核菌株菌落形態Fig.4 Colony morphology of parent strain and monocyte strain

表3 親本菌株與單核菌株的生長速度Table 3 The growth speed of mononuclear strains
2.4 雜交菌株的獲得與驗證
研究表明,共得到12株雜交菌株。在雜交菌株與親本菌株之間出現明顯的隆起拮抗帶,拮抗帶是不同菌株生長對抗的典型現象,純化后獲得的雜交菌株與親本菌株是不同的菌株個體。表4,圖5

表4 雜交菌株編號Table 4 hybrid strain
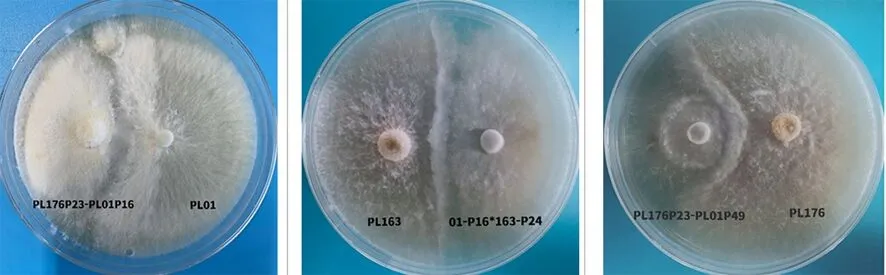
圖5 親本菌株與雜交菌株的拮抗實驗Fig.5 Antagonism test between parental strain and hybrid strain
3 討 論
目前利用原生質體單核化技術對新疆野生阿魏菇菌種進行提純復壯研究較為缺乏。制備原生質體的關鍵是有效去除細胞壁。然而,由于溶壁酶濃度、酶解時間、水浴溫度等諸多因素都會影響原生質體制備效率。酶解時間過短,生成的原生質體數量少但體積大;酶解時間過長,則原生質體的再生能力減弱。原慧等[12]研發新疆阿魏菇,35℃條件下酶解3 h,獲得原生質體數量最多,原生質體再生能力隨酶解時間的縮短而增強,但是原生質體細胞壁也隨著酶解時間的長短而受到不同程度的降解,因此,確定合適的酶解時間十分關鍵。研究發現,3株野生阿魏菇在酶解3.5 h時,原生質體釋放量最大,濃度達20.6×105個/ mL,因此,可將阿魏菇原生質體酶解時間控制在3.5 h。
野生阿魏菇菌齡的培養時間關系到原生質體獲得率高低,培養時間短原生質體釋放少,培養時間長則菌絲細胞壁難以充分酶解。不同菌類的最佳培養時間不同[8],白靈菇最佳培養時間為4~7 d[7],原慧等[12]阿魏菇菌絲選擇最佳培養時間均為8 d,阿魏菇菌絲最佳培養時間為5~7 d。3株野生阿魏菇菌株共獲得50個單核菌株。其中親本菌株P L163、PL01和PL176原生質體單核化得率分別為25.68%、12.8%和10.8%,獲得率較低,不同菌株獲得原生質體單核化率有著明顯差異,可能與原生質體制備時酶解時間不同有關。
研究3株野生阿魏菇菌株獲得2種交配型單核菌株非等比例分配,可能是由2種交配型的再生率不同導致[21]。研究表明,不同單核菌株的生命力強弱、細胞質對核的選擇作用以及某些與交配型基因連鎖的致死效應因子也同時影響著交配型單核菌株的分配比例[22],獲得更多樣本量的單核體以及控制好挑取時間可提高獲得2種交配型單核菌株的數量[23]。原生質體是遺傳轉化的廣泛受體之一,是多種植物的基因改造中都被作為接受外源基因的良好受體。原生質體單核化方法在研究細胞融合、突變與篩選等方面的應用普遍,利用原生質體單核化技術進行基因修飾已成為食用菌育種的有效途徑之一[24]。
4 結 論
3株野生阿魏菇在酶解3.5 h時原生質體釋放量最大,共獲得50個再生單核菌株。親本菌株PL163、PL01、PL176原生質體單核化率分別為 25.68%、12.8%、10.8%,3株阿魏菇菌株的原生質體再生能力在相同酶解條件下存在差異,獲得率普遍較低。每個親本菌株與單核菌株在菌絲長速、菌落形態方面均具有明顯差異,但無一致規律。3株親本阿魏菇原生質體單核菌株均獲得2種交配型,但獲得比例不同。3株阿魏菇野生菌株在原生質體再生單核獲得率、長速、形態等方面具有豐富的多樣性,獲得雜交菌株。
