基因c06orf223通過PI3K/AKT/mTOR信號轉(zhuǎn)導(dǎo)促進大腸癌進展的分子機制研究
2022-05-27 14:02:46焦璐熊鷹
河北醫(yī)藥 2022年10期
焦璐 熊鷹
我國大腸癌的發(fā)病率和病死率均保持上升趨勢。2015中國癌癥統(tǒng)計數(shù)據(jù)顯示,我國結(jié)大腸癌發(fā)病率、病死率在全部惡性腫瘤中均位居第5位,其中新發(fā)病例37.6萬,死亡病例19.1萬[1]。雷帕霉素靶蛋白(Mammalian target of rapamycin,mTOR)是PI3K/AKT通路下游的一種重要的絲氨酸-蘇氨酸蛋白激酶,其有整合細胞外多種信號刺激的作用,在體內(nèi)參與多條信號通路的傳導(dǎo)[2],其mTOR與細胞的生長以及凋亡等存在密切的關(guān)系。基因c06orf223與多種腫瘤的轉(zhuǎn)移存在密切的相關(guān)性,c06orf223基因的高表達與腫瘤細胞的增殖、侵襲以及遷移的形成相關(guān)[3,4]。PI3K(磷脂酰肌醇3-激酶)通過肌醇脂質(zhì)磷脂酰肌醇4,5-二磷酸[PI(4,5)P2]的磷酸化,形成第二信使分子磷脂酰肌醇(3,4,5)-三磷酸[PI(3,4,5)P3],其招募并激活包含pleckstrin同源結(jié)構(gòu)域的蛋白質(zhì),導(dǎo)致下游信號事件對細胞凋亡至關(guān)重要擴散、生存和移徙。Akt亞型有一個N端PH(pleckstrin homology)結(jié)構(gòu)域和一個激酶結(jié)構(gòu)域,這兩個結(jié)構(gòu)域由39個氨基酸的鉸鏈區(qū)分開。有催化活性的Akt調(diào)節(jié)多種底物的功能,這些底物參與細胞的存活、生長、增殖、代謝和蛋白質(zhì)合成,其是細胞存活的關(guān)鍵介質(zhì),其失活與各種應(yīng)激誘導(dǎo)的病理性細胞死亡和退行性疾病有關(guān)。PI3K/AKT/mTOR信號通路在細胞周期進程、細胞生長、存活、肌動蛋白重排和遷移以及細胞內(nèi)囊泡轉(zhuǎn)運等生物學過程中起著重要作用。本研究通過選取24例新鮮的大腸癌組織,從而探究基因c06orf223通過PI3K/AKT/mTOR信號轉(zhuǎn)導(dǎo)促進大腸癌進展的機制。
1 資料與方法
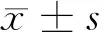
1.1 一般資料 選擇2019年1月至2020年9月于我院進行手術(shù)切除的24例新鮮的大腸癌組織及其癌旁組織(全層腸壁),其中選擇的癌旁組織距離癌組織約10 cm。患者中有男12例,女12例;年齡平均(46.82±5.42)歲;TNM分期均為Ⅲ~Ⅳ期57例。患者在手術(shù)切除前未做化療和放療治療。實驗所需的正常人腸上皮細胞系FHC均購自美國ScienCell研究實驗室。大腸癌組織組(n=24)、癌旁組(n=24)、正常人腸上皮細胞組(n=24),其中大腸癌組織組又分為空白對照組(n=12)和抑制組(n=12),其中空白對照組不抑制PI3K/AKT/mTOR信號通路。抑制組則加入AKT抑制劑Honokiol。AKT抑制劑Honokiol濃度為100 μg/ml,與細胞數(shù)的比例是1∶0.01 μg/ml。
1.2 方法 收集的組織標本用無菌PBS洗滌,迅速放置于液氮冷凍,然后放在-80℃保存。將購買的細胞復(fù)蘇后置于含有100 U/ml鏈霉素、100 U/ml青霉素、10%體積的胎牛血清的DMEM培養(yǎng)基中,于37℃,5.0%的CO2的細胞培養(yǎng)箱中培養(yǎng),以備后續(xù)實驗。
1.2.1 PI3K/AKT/mTOR信號通路在組織和細胞中激活情況的比較:采集患者大腸癌組織及其癌旁組織各5 g,研磨后3 000 r/min的條件下離心處理10 min,取上清液5 ml,Western blot檢測蛋白的表達水平;提取總RNA并進行RT-PCR,20 g/L瓊脂糖凝膠電泳50 min,紫外燈下用凝膠成像系統(tǒng)成像,用UVI圖像分析處理系統(tǒng)進行吸光度掃描,以目的基因片段/GAPDH片段的條帶灰度值比值來表示mTOR、PI3K與AKT mRNA相對含量。
1.2.2 過表達c06orf223促進PI3K/AKT/mTOR信號激活比較:取培養(yǎng)細胞轉(zhuǎn)染過表達的c06orf223質(zhì)粒,添加到培養(yǎng)基中,將細胞添加到上述培養(yǎng)基中,輕微晃動培養(yǎng)板,放置于培養(yǎng)箱中培養(yǎng)。按照1.2.1方法測定mTOR、PI3K與AKT蛋白和mRNA的相對含量。
1.2.3 下調(diào)c06orf223抑制PI3K/AKT/mTOR信號激活的比較:取培養(yǎng)細胞轉(zhuǎn)染的c06orf223 siRNA質(zhì)粒,添加到培養(yǎng)基中,將細胞添加到上述培養(yǎng)基中,輕微晃動培養(yǎng)板,放置于培養(yǎng)箱中培養(yǎng)。按照1.2.1方法測定mTOR、PI3K與AKT蛋白和mRNA的相對含量。
1.2.4 抑制PI3K/AKT/mTOR信號降低大腸癌細胞増殖能力的比較:將對數(shù)生長期的培養(yǎng)細胞加入96孔細胞培養(yǎng)板中,同時添加100 μl的細胞培養(yǎng)液和AKT抑制劑,將96孔板置于37℃,5.0%的CO2培養(yǎng)箱中培養(yǎng),當培養(yǎng)時長達到0、24、48、72 h時取出細胞培養(yǎng)板。然后在每孔中添加10 μl的MTT溶液,于37℃環(huán)境中反應(yīng)4 h。取上清溶液并添加150 μl的DMSO溶液,孵育10 min。在470 nm波長下測定OD值,OD值越大則表示細胞的增殖能力越強。
1.2.5 抑制PI3K/AKT/mTOR信號降低大腸癌細胞侵襲能力的比較:細胞侵襲能力通過Transwell小室進行測定。取100 μl濃度為3×105個/ml的培養(yǎng)細胞添加到Transwell小室上室中,于下室中添加500 μl的含血清的培養(yǎng)基和AKT抑制劑。分別在培養(yǎng)0、24、48、72 h后用濃度為4.0%的多聚甲醛溶液固定細胞20 min,通過結(jié)晶紫溶液染色20 min。于放大倍數(shù)為400的顯微鏡下觀察細胞的侵襲數(shù)目。
2 結(jié)果
2.1 PI3K/AKT/mTOR信號通路在組織和細胞中激活情況比較 3組PI3K/AKT/mTOR信號通路相關(guān)蛋白表達情況比較,正常腸細胞系最高,大腸癌組織最低,差異有統(tǒng)計學意義(P<0.05)。見表1,圖1、2。
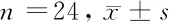
表1 PI3K/AKT/mTOR信號通路在組織和細胞中激活情況比較

圖1 組織和細胞中PI3K/AKT/mTOR通路mRNA表達
2.2 過表達c06orf223促進PI3K/AKT/mTOR信號激活的比較 大腸癌細胞中轉(zhuǎn)染c06orf223,Western blot檢測檢測細胞中PI3K/AKT/mTOR信號相關(guān)蛋白表達變化。結(jié)果發(fā)現(xiàn),過表達大腸癌細胞中的PI3K、AKT和mTOR的磷酸化水平較大腸癌細胞升高(P<0.05)。見表2。

圖2 組織和細胞中PI3K/AKT/mTOR通路蛋白表達
表2 過表達c06orf223促進PI3K/AKT/mTOR信號激活比較
2.3 下調(diào)c06orf223抑制PI3K/AKT/mTOR信號激活比較 大腸癌細胞中轉(zhuǎn)染下調(diào)c06orf223,Western blot檢測檢測細胞中PI3K/AKT/mTOR信號相關(guān)蛋白表達變化。結(jié)果發(fā)現(xiàn),下調(diào)基因大腸癌細胞中的PI3K、AKT和mTOR的磷酸化水平較大腸癌細胞降低(P<0.05)。見表3。
表3 下調(diào)c06orf223抑制PI3K/AKT/mTOR信號激活比較
2.4 抑制PI3K/AKT/mTOR信號大腸癌細胞増殖能力比較 大腸癌細胞經(jīng)過1 μm的AKT信號抑制劑處理以后,經(jīng)Transwell小室檢測抑制組細胞増殖能力較空白對照組下降(P<0.05)。見表4。
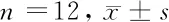
表4 抑制PI3K/AKT/mTOR信號大腸癌細胞増殖能力比較
2.5 抑制PI3K/AKT/mTOR信號大腸癌細胞侵襲能力比較 大腸癌細胞經(jīng)過1 μm的AKT信號抑制劑處理以后,經(jīng)Transwell小室檢測抑制組細胞侵襲數(shù)目較空白對照組降低(P<0.05)。見表5。
表5 抑制PI3K/AKT/mTOR信號大腸癌細胞侵襲能力比較
3 討論
大腸癌是最常見的消化道惡性腫瘤之一,在我國發(fā)病率和病死率均呈上升趨勢[5-7]。我國最常見的是大腸癌其發(fā)病率目前占到了消化道癌癥總發(fā)病率的85%,臨床上主要表現(xiàn)為便秘、腹瀉和便血等癥狀,嚴重者可及機生命[8-10]。磷脂酰肌醇3激酶(P13K)是一種位于胞質(zhì)的脂質(zhì)激酶;AKT,又稱蛋白激酶B,作為PI3K下游的一個重要的靶激酶,其是c-AKT表達編碼的一種絲氨酸/蘇氨酸蛋白激酶[11]。mTOR可以通過AKT發(fā)生直接或者間接的磷酸化反應(yīng),從而激活,磷酸化mTOR則不僅能夠通過上調(diào)細胞周期素加速細胞周期,同時還可以通過清除泛素蛋白抑制自噬的發(fā)生[12]。基因c06orf223是近年來發(fā)現(xiàn)的與大腸癌發(fā)生發(fā)展存在相關(guān)行的基因,屬于鼠源性基因啟動子的轉(zhuǎn)錄抑制因子,該基因間接調(diào)控著PI3K/AKT/mTOR信號通路,最終抑制細胞的增殖以及侵襲能力[13]。本研究結(jié)果顯示,過表達的c06orf223基因會促進PI3K/AKT/mTOR信號激活,而通過抑制PI3K/AKT/mTOR信號會降低大腸癌細胞侵襲和增殖能力。基因c06orf223可以通過調(diào)控PI3K/AKT/mTOR信號通路降低大腸癌細胞的増殖和侵襲能力。
本研究表明,PI3K/AKT/mTOR信號通路對大腸癌細胞的增殖和侵襲能力的形成至關(guān)重要,PI3KAkt信號通路會引發(fā)一系列的癌細胞的相關(guān)變化。PI3KAkt信號通路與Wnt/β-catenin信號通路存在較大的聯(lián)系,因為Wnt/β-catenin信號通路是大腸癌細胞增殖和分化過程中重要的信號通路,該通路功能的強弱程度與癌細胞的形成量的多少有關(guān)[14]。當AKT的T308和Ser473位點被3-磷酸肌醇依賴性蛋白激酶1和mTORC2激活并磷酸化后,AKT便能夠立即從胞質(zhì)內(nèi)轉(zhuǎn)移到細胞膜上,進而被活化成P-AKT,而活化后的P-AKT蛋白能夠再次通過調(diào)控多種細胞轉(zhuǎn)錄因子,從而發(fā)揮自身的作用,例如能夠促進細胞的增殖和抵抗細胞凋亡現(xiàn)象的發(fā)生,同時還能促進細胞的侵襲轉(zhuǎn)移等生物學作用[15]。磷酸化的AKT復(fù)合體可激活mTORC1,mTORC1磷酸化后又可激活其直接作用底物磷酸化核糖體蛋白S6激酶和真核生物始動因子4E結(jié)合蛋白1,進行下一級聯(lián)反應(yīng),控制細胞翻譯、核糖體生物合成、代謝等重要的生理功能。
腫瘤的發(fā)生發(fā)展是由于不同的基因在不同時期的表達以及功能出現(xiàn)紊亂而引起的復(fù)雜的病理性過程。許多促癌基因、抑癌基因以及與癌癥發(fā)展存在相關(guān)性的基因的激活或者異常失活均是腫瘤發(fā)生的基礎(chǔ)。因此,在研究腫瘤通路方面尋找新的相關(guān)基因?qū)τ陉U明腫瘤的發(fā)病機制以及診斷、預(yù)防和治療均有重要意義。有研究發(fā)現(xiàn)了AKT與c06orf223可能相互作用的證據(jù)并且其功能上可能與c06orf223協(xié)同,表明c06orf223可能為調(diào)控AKT的表達的重要[16-18]。并且通過研究與c06orf223直接和間接作用的蛋白所在信號通路,預(yù)測出c06orf223可能參與的PI3K/AKT/mTOR信號通路,且與大腸癌的發(fā)生發(fā)展密切相關(guān)[19,20]。
綜上所述,基因c06orf223能夠通過降低大腸癌細胞侵襲和增殖能力,進而緩解大腸癌的發(fā)生發(fā)展,其機制可能與PI3K/AKT/mTOR信號通路有關(guān)。而相關(guān)的基因功能序列以及作用靶位點仍有待進一步研究。
猜你喜歡
發(fā)明與創(chuàng)新(2022年30期)2022-10-03 08:40:56
鴨綠江(2021年35期)2021-04-19 12:24:18
動漫星空(興趣百科)(2020年12期)2020-12-12 05:31:40
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
中國生殖健康(2019年3期)2019-02-01 06:12:26
人大建設(shè)(2018年6期)2018-08-16 07:23:10
文理導(dǎo)航·科普童話(2017年5期)2018-02-10 19:42:14
無人機(2017年10期)2017-07-06 03:04:36
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
海軍航空大學學報(2015年3期)2015-11-11 17:20:00
