元江普通野生稻后代鎘分布特點及鎘積累動態變化規律
2022-05-24 03:28:34曾民陳佳李娥賢殷富有王玲仙曾黎瓊郭蓉
生態環境學報 2022年3期
關鍵詞:水稻
曾民,陳佳,李娥賢,殷富有,王玲仙,曾黎瓊,郭蓉
1. 云南農業科學院生物技術與種質資源研究所/云南省農業生物技術重點實驗室/農業部西南作物基因資源與種質創制重點實驗室,云南 昆明 650223;
2. 云南農業科學院經濟作物研究所,云南 昆明 650205
水稻是中國種植面積最大、食用人口最多的糧食作物,水稻易吸收并在籽粒內富集鎘(Cd),這已成為中國人體鎘攝入的主要途徑(文典等,2021)。在中國中南及西南部分地區,稻米中的鎘含量能高達1—2 mg·kg-1,遠高于食品安全國家標準0.2 mg·kg-1(GB 2762—2012)(甄燕紅等,2008)。目前,中國土壤鎘污染面積約為1.4×104hm2,農田土壤超標位點約為7%(文典等,2020)。如何保證鎘污染農田的水稻生產安全是急需解決的難題,而種植籽粒低積累鎘的水稻品種是保障水稻食用安全最直接、最有效的方法(Zhao et al.,2015)。
水稻遺傳基因不同是造成水稻品種籽粒鎘含量差異的主要原因(Ueno et al.,2010;Sun et al.,2016)。水稻通過根系吸收鎘離子后,裝載到木質部并隨蒸騰作用轉運至地上部,然后再分配到莖葉中,最后通過韌皮部運輸進入籽粒(賀慧等,2014)。水稻在吸收運輸鎘的同時,還能通過各組織器官內的物質與鎘結合,例如纖維素、半纖維素、果膠、非蛋白質巰基及谷胱甘肽等,并將其大量儲存在細胞壁或液泡內,降低鎘活性,減少其進入籽粒的數量(嚴勛等,2019)。因此,水稻籽粒鎘含量不僅與水稻對Cd吸收及運送到籽粒的能力相關,還與限制并降低鎘活性相關(史靜等,2015)。因此,通過研究低/高鎘株系各器官內的鎘含量、亞細胞分布及生育期內各組織中鎘的積累動態變化,可以更好地了解不同水稻材料對鎘的吸收、轉運、積累存在差異的原因,以及如何限制鎘向籽粒轉運。
普通野生稻具有豐富的遺傳多樣性,擁有許多栽培稻消失或者不具備的優良性狀(楊慶文等,2004)。元江普通野生稻根系發達、枝葉茂盛、生命力強、穗型散、種子小(黃興奇等,2005)。這些性狀表明,元江普通野生稻具有很強的根系吸收、光合代謝及物質轉運能力。同時,其種子少,籽粒小,繁殖后代所消耗能量低,元江普通野生稻能儲存大量物質來應對惡劣的環境,并進化出多種生物及非生物抗性來提高其自身生存能力(黃興奇等,2005)。與現代栽培稻相比,野生稻植株體內富含更多的有益礦質元素和更少的有害重金屬(徐玲玲等,2005;徐靖等,2014)。研究還發現,水稻對鎘的吸收和轉運是依靠其他礦質元素的離子通道,尚未發現吸收和轉運鎘的專用通道(劉仲齊等,2019)。因此從生物學性狀的角度分析,元江普通野生稻根系應具有較強的鎘吸收能力,同時也應該具有很強的抵抗鎘毒害能力。元江普通野生稻滲入系是具有普通野生稻基因和性狀的后代群體,是創新性水稻種質資源,是進行基因挖掘和理論研究的優良群體材料(柯學等,2018)。元江普通野生稻滲入系在籽粒鎘含量方面存在極顯著差異,可從中篩選出多份鎘低積累株系及多份高積累株系(曾民等,2021)。目前對元江普通野生稻滲入系后代株系對鎘吸收、轉運和積累特征均不清楚,因此本文以元江普通野生稻滲入系群體中籽粒鎘含量低的株系(低鎘株系)和籽粒鎘含量高的株系(高鎘株系)為研究材料,分析鎘在這兩類株系各器官內的分配和亞細胞分布差異以及鎘積累的動態變化特點,從而揭示元江普通野生稻滲入系后代中低鎘株系與高鎘株系間對鎘的吸收和分配差異,為闡明元江普通野生稻滲入系鎘積累的機理提供了理論基礎。
1 材料與方法
1. 1 試驗材料
以前期開展的元江普通野生稻滲入系低鎘積累材料篩選試驗中獲得的籽粒鎘含量低的株系(GJ71、GJ91)及籽粒鎘含量高的株系(GJ11、GJ110)為試驗材料,并選取籽粒鎘含量居中的株系(GJ38、GJ114)作為對照材料。
1.2 試驗方法
大田試驗:試驗地為個舊市倘甸鄉水旱輪作農田,前茬作物為蠶豆,農田存在Cd 污染,主要是由污水灌溉導致,現周邊已無污染源(曾民等,2019)。采用隨機區組排列,3次重復,在水稻播種前平整土地、劃定小區,對小區地塊進行多次深耕深翻處理,每個小區面積30 m2(6 m×5 m),株距為15 cm,行距為20 cm。四周設置2 m的保護行,相鄰小區間隔40 cm。于2020年3月下旬育秧,4月移栽,10月收獲。土壤為褐色土,其基本理化性質為:pH值6.52,有機質21.97 g·kg-1,堿解氮1.24 g·kg-1,有效磷 12.2 mg·kg-1,速效鉀 92.7 mg·kg-1,土壤總Cd質量分數為1.01 mg·kg-1。田間管理按當地大田常規方式進行管理。
1.3 樣品采集、處理與分析
1.3.1 樣品采集與處理
在不同生育期(分蘗期、孕穗期、抽穗期、灌漿期、乳熟期及完熟期)采集田間水稻樣品各 10株,用不銹鋼剪刀將根、莖、葉分離,先用自來水小心清洗水稻植株的根系、莖、葉,然后再用蒸餾水清洗,自然風干,粉碎過0.15 mm篩網,保存于自封袋中備用;待水稻成熟后,進行測產并采集植株樣品,每個小區隨機采集 10株植株樣,將根、莖、葉分離,先用自來水小心清洗水稻植株的根系及其莖、葉,然后再用蒸餾水清洗,稻谷風干后用礱谷機脫殼,再用粉碎機將籽粒和谷殼粉碎,過0.15 mm篩網,保存于自封袋中備用。
1.3.2 Cd的亞細胞組分提取與消解
在分蘗期和灌漿期,分別采集田間種植的不同水稻株系植株各10 株,用去離子水清洗3次,各稱取根、莖、葉鮮樣2.0 g用于亞細胞組分提取(付鑠嵐等,2017)。提取步驟如下:將樣品剪成約 2—3 mm的小段,在預冷研缽內加入液氮研磨,研磨成粉末后置4 ℃冰箱保存。采用Tris-HCl緩沖液(0.154 g·L-1DTT,0.25 mol·L-1蔗糖溶液,pH 7.4)作為亞細胞提取液,樣品和提取劑以1∶10的比例充分混合,用漏斗和尼龍布過濾到離心管中,將尼龍布上的殘渣(細胞壁)裝入三角瓶中,濾液裝入離心管,在15000 r·min-1下離心40 min,將上清液和沉淀分別倒入三角瓶中,分別為可溶性部分和細胞器部分。最后分別向所用三角瓶中加4∶1的硝酸和高氯酸的混合酸10 mL(可溶性部分和細胞器)和20 mL(細胞壁),待充分消化,消煮至澄清,蒸餾水定容后放入4 ℃冰箱保存待用。采用石墨爐原子吸收光譜儀(美國Thermo Elemental公司生產)測定Cd含量。
1.3.3 樣品消解與測定
植物樣品的消解與測定:稱取0.2 g處理好的樣品于50 mL離心管中,加入2 mL優級純硝酸,混合均勻,室溫下放置一夜后,用微波消煮爐消煮,程序是55 ℃ 10 min;75 ℃ 10 min;95 ℃30 min,采用標準樣品(GBW 08502,大米粉標準物質)同樣消煮。消煮后的樣品用2%的硝酸溶液稀釋至40 mL,過0.45 μm 濾膜,4 ℃保存待測。消解液中的重金屬濃度用石墨爐原子吸收光譜儀測定。
土壤樣品的消解與測定:稱取0.2 g土壤樣品于石英玻璃管中,加入 5 mL 王水(HCl∶HNO3=3∶1),混勻后室溫下平衡過夜,再將土壤樣品采用開放式消煮爐消解,消解程序是100 ℃ 1 h,120 ℃ 1 h;保持溫度在140 ℃,直到溶液清澈,將消解后的樣品置于通風櫥揮發酸,然后用 2%的硝酸溶液定容到20 mL,過0.45 μm濾膜后備用。空白和標準樣品(GBW 07405,土壤標準物質)同時消煮后用石墨爐原子吸收光譜儀測定Cd含量。
1.4 數據分析與統計
試驗數據利用Microsoft Excel 2007進行均值、標準差計算和繪圖,利用SPSS 19.0進行方差分析和LSD多重比較,采用Pearson相關系數法進行相關性分析,采用非參數檢驗中K獨立樣本檢驗不同株系差異性。
2 結果與分析
2.1 元江普通野生稻滲入系株系Cd 含量
由表1可知,不同滲入系株系各組織Cd含量為根>莖>葉>谷殼>籽粒,Cd主要集中在滲入系株系根部。高鎘株系(GJ11、GJ110)根部 Cd質量分數分別為24.2、22.4 mg·kg-1,幾乎是低鎘株系(GJ71、GJ91)的 2倍。高鎘株系籽粒和谷殼的Cd含量也顯著高于低鎘株系。低鎘株系GJ91莖、葉中的鎘質量分數分別為1.77、1.75 mg·kg-1,顯著高于其他株系,表明GJ91能將地上部的鎘更多的轉運并儲存在莖、葉內。低鎘株系GJ71 與GJ91在莖、葉、谷殼中的鎘含量均存在顯著差異性,這表明 2份低鎘株系對轉運到地上部的鎘的分配方式存在差異。
高鎘株系總Cd積累量顯著高于其他株系(見表 1),其中 GJ11 積累量最多,為 268.01 μg·kg-1,GJ71 Cd積累量最少,為93.15 μg·kg-1,兩者相差2.88倍。高鎘株系的籽粒和根中的Cd積累量顯著高于低鎘株系,其中GJ110籽粒Cd積累量最多,為 18.94 μg·kg-1,GJ71 籽粒 Cd 積累量最少,為 0.47μg·kg-1,而GJ91莖部Cd積累量最多。通常,以地上部與地下部 Cd含量比值(S∶R)表示水稻對 Cd的轉運能力(李江遐等,2017),本文采用水稻籽粒與根部的鎘含量的比值(G∶R)表征水稻將 Cd轉運到籽粒的能力,比值越小表示積累能力越強。高鎘株系(GJ11、GJ110)根/籽粒比值分別是 1∶43.21和 1∶32.94,遠低于低鎘株系(GJ71、GJ91)的根/籽粒的 1∶67.5和 1∶471,這表明高鎘株系將鎘運輸到籽粒的能力遠高于低鎘株系,其中GJ110的運輸能力最強,GJ71最弱。
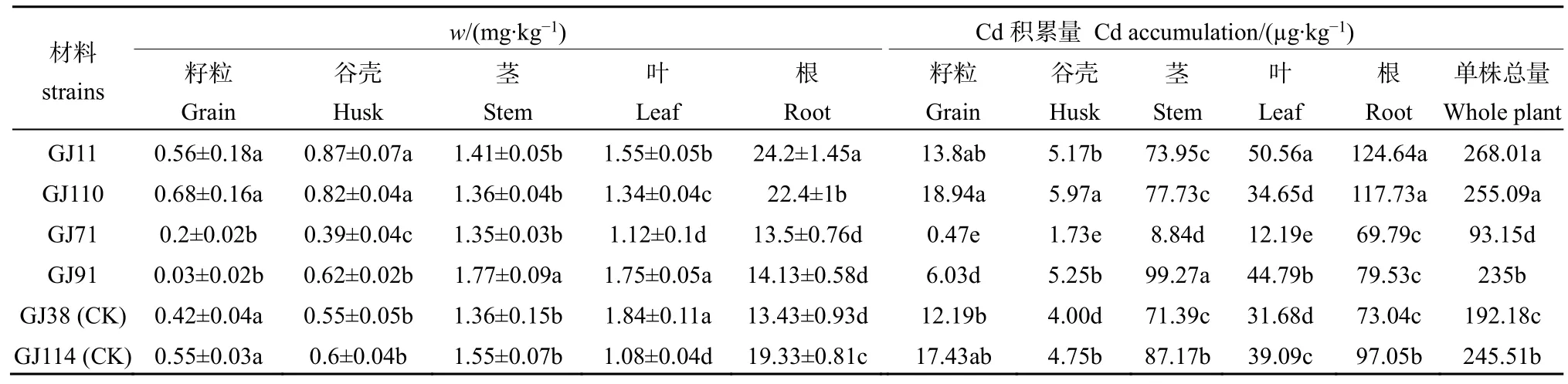
表1 水稻完熟期不同器官Cd質量分數Table 1 The Cd content in different organs of six strains during maturing stage
2.2 元江普通野生稻滲入系株系籽粒Cd相關性分析
對滲入系株系籽粒鎘含量與根、莖、葉、稻殼鎘含量進行相關性分析(圖 1),結果發現,籽粒鎘含量與谷殼、根、莖中的鎘含量存在正相關性,相關系數分別為0.737、0.739、0.519,其相關性順序為谷殼>根>莖。
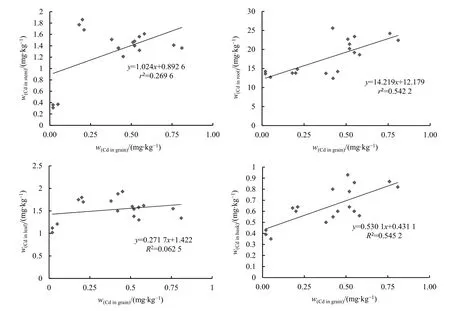
圖1 籽粒鎘含量與其他器官鎘含量的相關性分析Figure 1 Correlation analysis of Cd content in grains and other organs
2.3 元江普通野生稻滲入系株系Cd含量動態變化規律分析
從圖2可知:生育期內,各株系根部Cd含量變化趨勢一致,均表現為緩慢上升,且高鎘株系鎘含量均高于低鎘株系。各株系莖部的鎘含量在抽穗期前高鎘株系高于低鎘株系,抽穗期后低鎘株系高于高鎘株系,其中 GJ71鎘含量在抽穗期后快速上升,到乳熟期后又快速下降,GJ91則是在乳熟期后快速上升。從各株系葉部鎘含量來看,低鎘株系GJ91在乳熟期后鎘含量快速上升,高于高鎘株系。
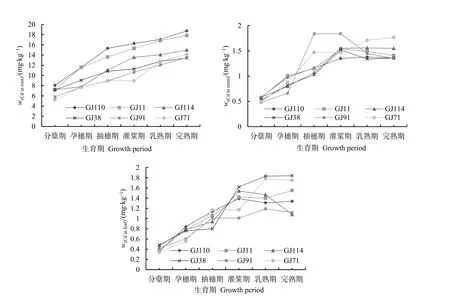
圖2 不同生育期水稻各器官內Cd積累動態變化Figure 2 Dynamic changes of Cd accumulation in different organs of six rice strains at different growth stages
2.4 元江普通野生稻滲入系株系Cd含量的亞細胞分布
不同株系各器官鎘含量亞細胞分布(見表 2)均是細胞壁>可溶部分>細胞器,Cd在細胞壁分布最多,約占總量的39.47%—60.39%。分蘗期細胞壁Cd含量順序為根>莖>葉,灌漿期細胞壁Cd含量為根>葉>莖。從分蘗期到灌漿期,不同株系各器官內分布在細胞壁、細胞器、可溶部分的鎘含量均增加,鎘含量增加量為細胞壁>可溶部分>細胞器。
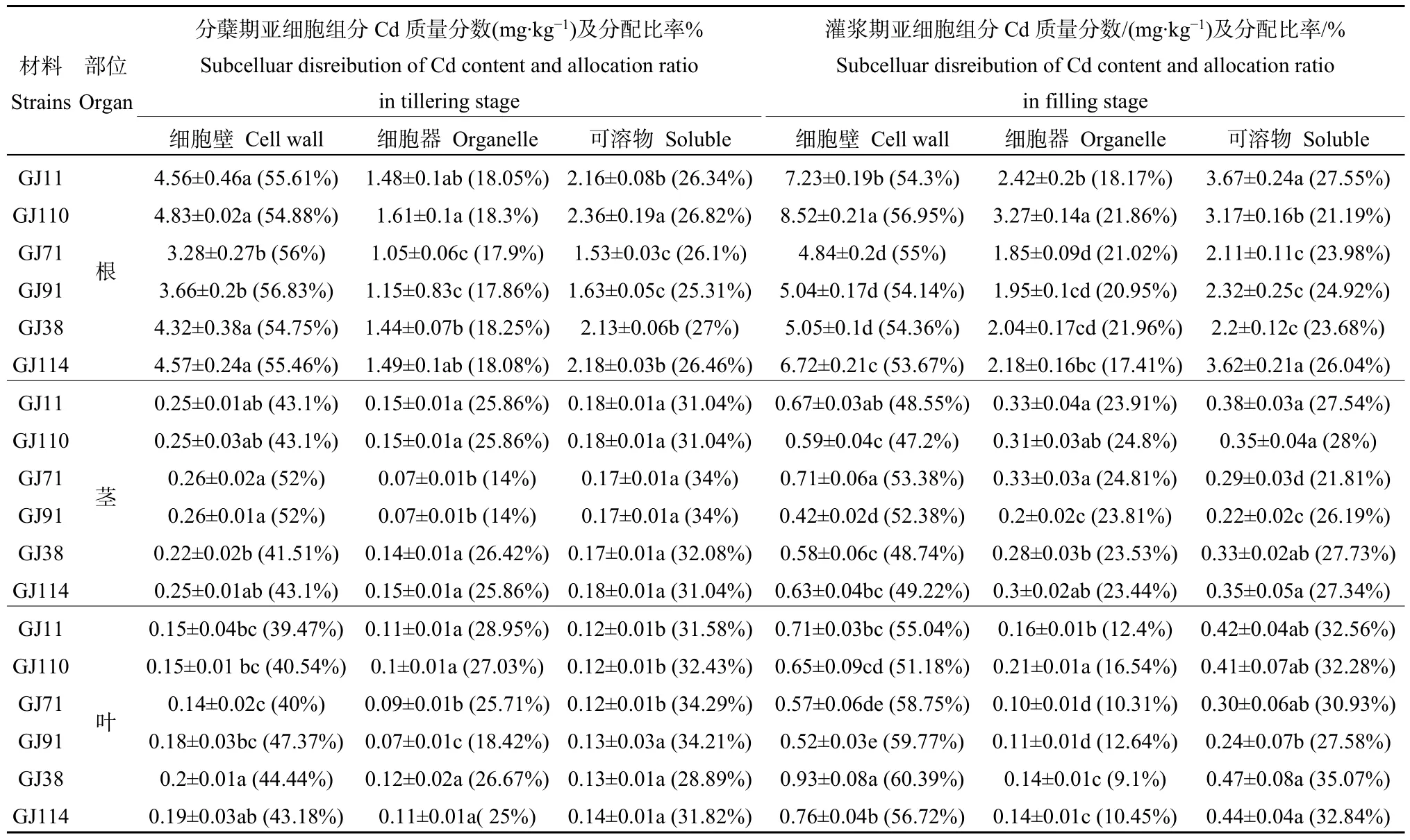
表2 滲入系水稻材料的不同時期亞細胞分布Table 2 Subcelluar disreibution of Cd content in introgression lines rice at different stage
低鎘株系與高鎘株系根部細胞壁、細胞器及可溶部分的Cd含量均存在顯著差異。分蘗期,低鎘株系與高鎘株系的莖和葉部中細胞器的Cd含量存在顯著差異;灌漿期,低鎘株系與高鎘株系的莖部可溶部分的Cd含量和葉部細胞壁的Cd含量均存在顯著差異。
3 討論
滲入系株系中存在葉部鎘含量超過了莖部的現象,這與以往研究得出的水稻莖部鎘含量高于葉部結論不同(李坤權等,2003),并且不同株系籽粒鎘含量與葉片鎘含量之間沒有明顯的相關性,也與以往研究得出的高積累水稻品種根系、莖和葉Cd含量均高于低積累水稻品種(付鑠嵐等,2017)不一致,這可能與元江普通野生稻滲入系株系保留野生稻遺傳特性有關。當各滲入系株系籽粒成熟時,其植株生長依然旺盛,葉片未見枯萎,光合作用和新陳代謝旺盛,依然能將鎘運輸到葉片組織,所以滲入系株系葉部鎘含量普遍偏高。
分蘗期后是水稻干物質積累的主要時期,灌漿期是干物質從莖葉向稻谷轉移的重要時期,也是水稻籽粒蛋白質與氨基酸合成的關鍵時期(吳文革等,2007)。此時,莖葉中蛋白組和其他有機質將被分解成氨基酸等小分子物質,并作為合成蛋白質的原料運輸到籽粒部位,而Cd可能也會隨OsLCT1和OsNRAMP1等轉運蛋白轉運至籽粒(Uraguchi et al.,2011;Tiwari et al.,2014)。本研究中,高鎘株系灌漿期后莖葉中鎘含量出現下降,這可能是由于鎘隨氨基酸等活性物質向籽粒運輸,而低鎘株系莖部鎘含量上升及 GJ91葉鎘含量上升,可能是由于鎘的積累速度大于鎘向籽粒轉運的速度,也可能是存在某種機制阻礙莖葉部鎘向籽粒轉運。
水稻中Cd主要與有活性基團的物質結合:這些物質包括細胞壁上纖維素、半纖維素、果膠、金屬硫蛋白和植物螯合肽等,細胞可溶部分中的有機酸和氨基酸等,以及細胞器膜上的活性物質(Siebers et al.,2013)。研究表明,在鎘脅迫下,植物首先是通過細胞壁上的纖維素、半纖維素和果膠等大分子物質與鎘結合,當植物細胞壁上的重金屬結合位點飽和后,鎘才會與大量的有機配位體結合形成穩定態的螯合物,并被轉運至液泡中儲藏,從而降低細胞質中自由重金屬離子的濃度(Wang et al.,2015)。本研究發現,從分蘗期到灌漿期,各器官分布在細胞壁、細胞器、可溶部分的鎘含量均有所增加,由此推測鎘可能是按照一定的比例分布在細胞壁、細胞器和可溶部分上,而不是與細胞壁上的大分子物質先結合至飽和后,再轉運到可溶物和細胞器上。因為水稻的根系在分蘗期后已發育完全,如果根系已經飽和,那么灌漿期細胞壁上的鎘含量就不應該再增加。
4 結論
(1)高鎘株系對鎘的吸收及轉運鎘到籽粒的能力均顯著高于低鎘株系。低鎘株系GJ71與GJ91間地上部對鎘的分配存在顯著差異。低積累株系GJ91莖葉的鎘含量顯著高于其他株系,它能將更多鎘分配并儲存在莖葉中,從而降低籽粒鎘含量。高鎘株系總Cd積累量顯著高于低鎘株系,低鎘株系GJ71總Cd積累量顯著低于其他株系。
(2)低/高鎘株系根部Cd 含量積累動態變化趨勢一致,莖和葉的Cd含量積累的動態變化分別從孕穗期和抽穗期開始有所不同,低鎘株系的鎘含量在灌漿期后上升,而高鎘株系的鎘含量在灌漿期后下降或維持不變。
(3)低/高鎘株系各組織器官鎘含量的亞細胞分布為細胞壁>可溶部分>細胞器,細胞壁Cd含量約占總量的40%—60%。從分蘗期到灌漿期,各組織細胞壁、細胞器和可溶部分的鎘含量均增加。
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
