不同杉木林分類型植物多樣性及其土壤碳氮關系的研究
2022-05-24 03:28:12胡靚達周海菊黃永珍姚賢宇葉紹明喻素芳
生態環境學報 2022年3期
關鍵詞:植物
胡靚達,周海菊,黃永珍,姚賢宇,葉紹明,喻素芳
廣西大學林學院,廣西 南寧 530004
人工林作為森林資源的重要組分,對于森林的可持續經營意義重大。目前,中國人工林純林面積大,生態系統結構和功能簡單,易造成林產品單一、生物多樣性降低、病蟲害蔓延、地力衰退和生產力低下等諸多問題(周霆等,2008)。營造混交林在一定程度上可改善土壤理化性質、提升林分涵養水源功能(王青天,2014)、減少林木病蟲害發生(葉紹明等,2010),進而提高林分的生物、生境多樣性(Chianucci et al.,2016;Kooijman et al.,2019)及林分生產力,從而建立更穩定的森林生態系統。然而,混交樹種選擇可能會產生不同的經營效果,如杉木(Cunninghamia lanceolate)與木荷(Schima superba)以1∶1進行行間混交可促進杉木生長,混交林中的杉木樹高、胸徑皆大于杉木純林(李勇,2016),而與細葉青岡(Cyclobalanopsis gracilis)混交杉木的生長反而受到了抑制(劉雨暉,2018a);桉樹(Eucalyptus)與不同樹種混交對林下植物物種多樣性有顯著影響,尤其與望天樹(Parashorea chinensis)、格木(Erythrophleum fordii)和紅錐(Castanopsis hystrix)混交更具優勢(龐圣江等,2020)。混交林經營不僅增加了林下植被多樣性,還能提高土壤養分含量,維持地立平衡(劉世榮等,2018),如石亮等(2021)對樟子松(Pinus sylvestris var. mongolica)人工林混交改造后的土壤養分研究發現,38 a生的樟子松與樟子松、銀中楊(Populus alba)、色木槭(Acer mono)幼苗混交后,土壤養分均高于沙地樟子松純林,其中有機質、堿解氮含量在樟子松×色木槭、樟子松×銀中楊混交林達到最高,但有效磷含量則低于樟子松×樟子松混交林。
植被和土壤是一個密不可分的整體,植物通過根系下扎、分泌物、凋落物及調節小氣候等影響土壤性狀(王光州等,2021),同時植物個體之間以及植物種間競爭土壤資源也會對植物群落物種組成和生長產生影響(余軒等,2021)。生物多樣性是人工植被恢復重要評價指標,而植物多樣性是生物多樣性在植物水平上的表現形式(甄倩等,2020),是基于群落的物種數和個體數及其分布均勻程度的統計量(趙耀等,2018),它與土壤環境的協調程度反映出森林生態系統的穩定性。因此,了解植物多樣性與土壤養分因子的相互關系對評價人工林可持續經營水平有著重要意義。
杉木是中國南方亞熱帶地區的主要用材樹種之一,因其生長快、適應性強、經濟效益高、材質好等特點而被廣泛種植(李萌等,2020)。由于杉木純林經營引發系列生態問題,嚴重制約林地的可持續利用(明安剛,2017)。目前,已有大量試驗證明杉木人工混交林可以有效促進土壤中的養分循環(姜俊等,2019),改善土壤的理化性質(徐海東等,2020),增強土壤肥力,促進林木生長(唐學君等,2019),提升林分的涵養水源功能等(張莉,2019)。相較于杉木純林,混交林更有利于提高植物多樣性,維持生態系統穩定(黃冬梅,2017;劉雨暉,2018)。同時,前人研究發現土壤有機碳含量和有機碳密度在很大程度上依賴于植被群落多樣性(劉林馨等,2018),土壤全氮含量與植物多樣性指數呈顯著正相關關系(劉俊杰等,2021),植物多樣性與土壤碳氮含量之間關系密切。然而,不同樹種與杉木混交后植物多樣性與土壤碳、氮變化的關系鮮見報道。因此,本文以位于廣西壯族自治區憑祥市的中國林業科學研究院熱帶林業研究中心伏波實驗場的杉木×紅錐混交林、杉木×大葉櫟(Quercus griffithii)混交林以及杉木純林為研究對象,旨在探討紅錐、大葉櫟與杉木混交后對植被多樣性特征及其土壤碳、氮含量的影響,并分析土壤碳、氮含量及碳氮比變化與植被多樣性的相關性,為杉木人工林的可持續經營提供科學依據。
1 研究方法
1.1 研究區概況
試驗地位于廣西壯族自治區憑祥市中國林業科學研究院熱帶林業研究中心的伏波實驗場,該試驗區屬南亞熱帶季風型半濕潤-濕潤氣候,年日照時數1218—1620 h,年平均氣溫20.5—21.7 ℃,年均降水量 1200—1500 mm。干濕季節明顯,4—9月為雨季,10月至翌年3月為旱季,雨熱同季的氣候特點對植物生長十分有利。土壤為花崗巖發育的山地紅壤(盧立華等,2008)。林下主要灌木有三椏苦(Evodia lepta)、水東哥(Saurauia tristyla)、杜莖山(Maesa japonica)、粗糠柴(Mallotus philippensis)、中平樹(Macaranga denticulata)、九節(Psychotria rubra)、黃毛榕(Ficus esquiroliana)、潺槁木姜子(Litsea glutinosa)等,草本大多為半邊旗(Pteris semipinnata)、烏毛蕨(Blechnum orientale)、扇葉鐵線蕨(Adiantum flabellulatum)、芒萁(Dicranopteris dichotoma)、淡竹葉(Lophatherum gracile)等。
1.2 樣地設置
試驗林均為 1993年營造的杉木純林,初植密度為2500 plant·hm-2。1999年和2003年分別進行了強度約為30%的透光伐和強度約為40%的撫育間伐,2008年實施近自然化改造,對杉木純林進行疏伐,近自然改造林杉木保留密度為600 plant·hm-2,疏伐后在林下以 400 plant·hm-2的密度分別均勻套種紅錐、大葉櫟,構建杉木混交林,并設置疏伐后保留密度為1000 plant·hm-2的杉木純林作為對照。在杉木×紅錐混交林(MCC)、杉木×大葉櫟混交林(MCQ)和杉木純林(PCL)中選取自然環境條件基本一致的典型區域,各設立3塊20 m×20 m的固定試驗樣地,同一林分中各樣地間距離20 m以上,即每種林分設3個重復。2018年10—11月在固定樣地進行每木調查,樣地基本情況見表1。

表1 不同類型林分樣地基本情況Table 1 Basic situation of different stand types on the ground
1.3 樣地調查
將3種不同類型林分的20 m×20 m樣地分成4個10 m×10 m的中樣方,分別調查各個中樣方里的喬木(DBH≥1 cm)樹種種類、數量、胸徑、樹高和冠幅;在各個中樣方(10 m×10 m)的左上角設置1個5 m×5 m的小樣方,調查并記錄小樣方內喬木幼樹幼苗(H≥30 cm,DBH<1 cm)及所有灌木種類、數量、平均高度和覆蓋度;同時再設置1個1 m×1 m的微型樣方,調查并記錄微型樣方內所有草本植物的種類、數量、平均高度和覆蓋度。
1.4 土壤樣品采集與分析
在每個20 m×20 m樣地內按“S”形設置5個采樣點,分別在0—20 cm土層采集原狀土壤1 kg。將各個樣地內土壤樣品分別混合均勻。將每個混合樣品沿自然結構輕輕掰開,并過2 mm篩除去動植物殘體和小石塊等,置于實驗室內自然風干。風干后的土壤用磨土機粉碎后過<0.2 mm篩,測定土樣的有機碳(SOC)和全氮(TN)含量,其中有機碳采用高溫外熱重鉻酸鉀氧化-分光光度法,全氮采用消煮法并使用連續流動分析儀測定(魯如坤,2000)。
1.5 數據處理與統計分析
1.5.1 植物多樣性及重要值測定
統計樣方內物種組成并分析樹種重要值和多樣性。重要值可作為判斷群落中優勢樹種的衡量指標(Naeem et al.,1997)。其計算公式分別為:喬木重要值 (Iv1)=(相對多度+相對顯著度+相對頻度)/3,灌木 (草本) 重要值 (Iv2)=(相對多度+相對蓋度+相對頻度)/3。植物多樣性利用豐富度指數(Rrichness index,S)、香農維納指數(Shannon-Wiener index,H′)、辛普森指數(Simpsonindex,D)、均勻度指數(Pielou index,E)進行分析(Magurran,1988)。
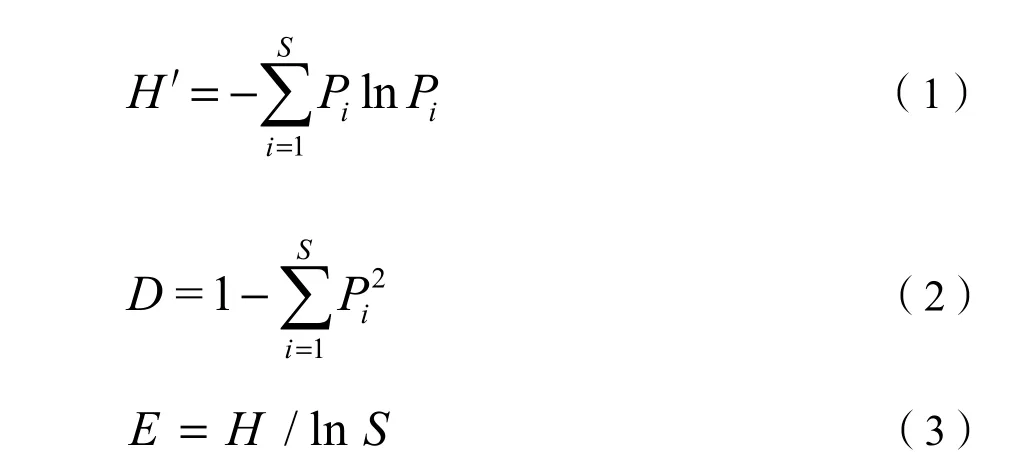
式中:
S——總物種數;
Pi——種的個體數占群落中總個體數的比例。
1.5.2 數據統計分析
應用 Excel 2007進行數據計算和制圖,SPSS 19.0對數據進行單因素方差(One-way ANOVA)分析和Pearson相關性分析,并采用鄧肯法進行多重比較(P<0.05)。
2 結果與分析
2.1 不同林分類型群落的物種組成及重要值
經調查發現(表 2),3種林分類型喬木層物種數表現為PCL>MCQ>MCC。其中,MCC物種數為10種,除杉木和紅錐外,重要值超過5%的樹種還有香梓楠和中平樹;MCQ物種數為14種,除杉木和大葉櫟外,格木、三椏苦、黃毛榕、紅錐也占據一定的優勢;PCL物種數為18種,除主要造林樹種杉木外,黃毛榕和毛黃肉楠的重要值也都在5%以上。此外,在3種林分類型中,水東哥均有分布,但重要值均低于5%。
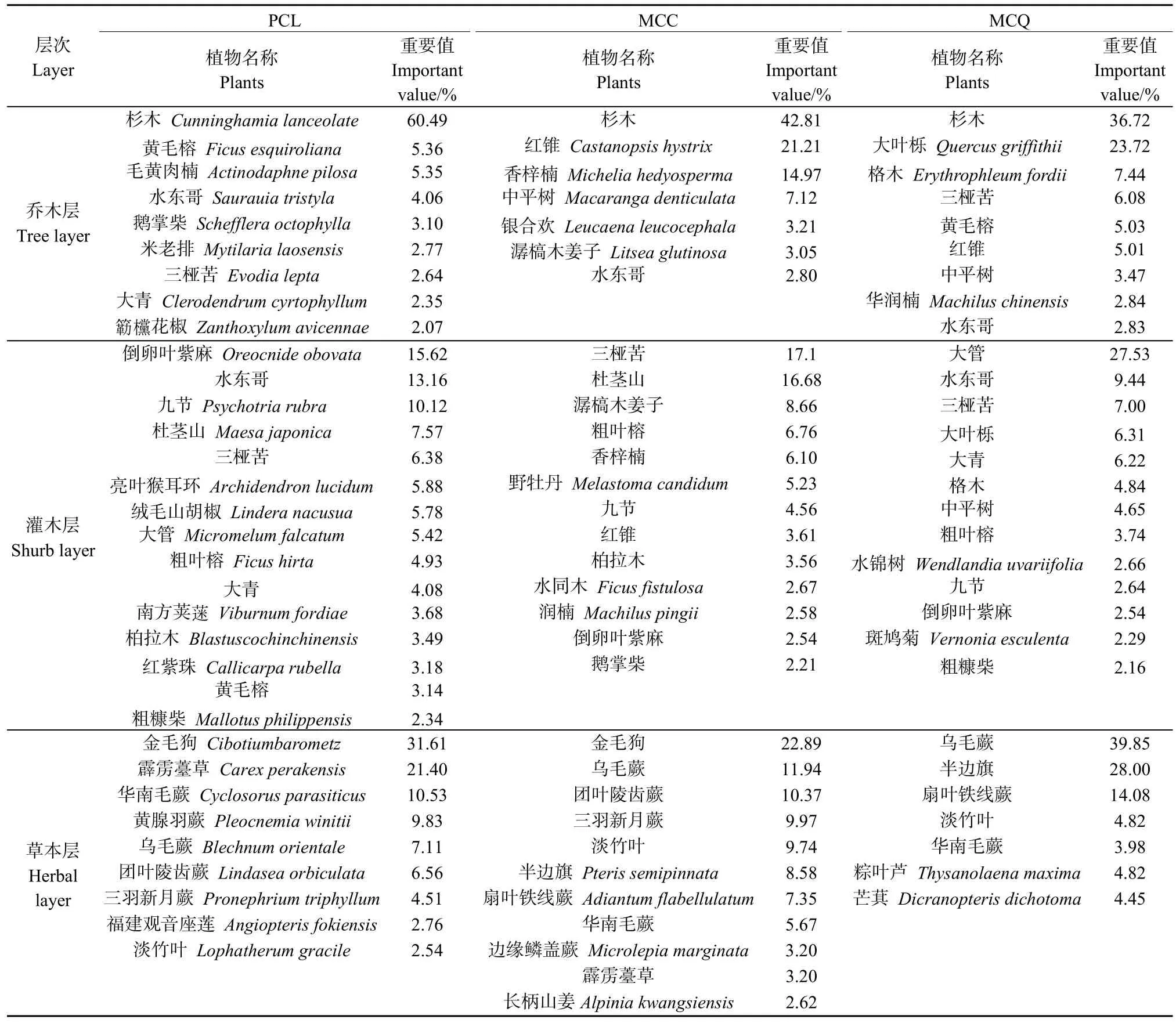
表2 不同林分類型物種組成及重要值Table 2 Species composition and important value of plants in different stand types
灌木層主要由喬木的幼樹幼苗(H≥30 cm,DBH<1 cm)及叢生灌木組成。3種林分類型中灌木層物種數為 MCQ>MCC>PCL。不同林分灌木層優勢種不同,其中,MCC植物物種數為23種,優勢種為三椏苦、杜莖山,重要值分別為 17.10%和16.68 %;MCQ的植物物種數為24種,主要優勢種為大管,重要值為27.53%,水東哥、三椏苦、中平樹和大青(Clerodendrum cyrtophyllum)的重要值都在5%—10%之間;PCL的植物物種數為18種,優勢種為倒卵葉紫麻、水東哥、九節,重要值分別為15.62%、13.16%、10.12%。
草本層的優勢種主要為蕨類及禾本科植物。MCC草本層物種數為 13種,優勢種為金毛狗(Cibotiumbarometz)、烏毛蕨、團葉陵齒蕨(Lindaseaorbiculata),重要值分別為 22.89%、11.94%、10.37%;MCQ草本層物種數為7種,優勢種為烏毛蕨、半邊旗、扇葉鐵線蕨,重要值為 39.85%、28.00%、14.08%;PCL草本層物種數為11種,優勢種為金毛狗、霹靂薹草(Carex perakensis)、華南毛蕨(Cyclosorus parasiticus),重要值為31.61%、21.40%、10.53%。
2.2 不同林分類型植物多樣性
對MCC、MCQ、PCL林分喬木層、灌木層、草本層以及群落的植物多樣性指數進行分析發現(圖1),喬木層Shannon-Wiener指數、Simpson指數和Pielou均勻度指數表現為MCC和MCQ顯著高于PCL(P<0.05),MCC和MCQ的豐富度指數與PCL差異不顯著(P>0.05)。在3種林分類型中,MCQ的豐富度指數、Shannon-Wiener指數和Simpson指數最高,Pielou均勻度指數最高的是MCC。草本層、灌木層和群落中,MCC、MCQ的植物多樣性指數均與PCL無顯著差異。MCC的3種群落層次植物多樣性指數均無顯著差異;在MCQ中,豐富度、Shannon-Wiener和Simpson指數表現為灌木層>喬木層>草本層,且差異顯著,Pielou均勻度指數以灌木層最高,且灌木層和草本層顯著高于喬木層;PCL中,植物多樣性指數均以灌木層最高,灌木和喬木層的豐富度指數顯著高于草本層,灌木層Shannon-Wiener和Simpson指數與喬木層、草本層存在顯著差異,喬木層Pielou均勻度指數顯著低于灌木和草本層。

圖1 不同林分類型植物多樣性指數比較Figure 1 Comparative study on the indexes of plant species diversity in different forest types
2.3 不同林分類型土壤碳氮含量的變化
不同林分土壤有機碳含量以MCQ最高,且顯著高于 PCL(P<0.05),MCC土壤有機碳含量高于PCL,但差異不顯著(P>0.05);全氮含量由高到低依次為 MCQ、PCL、MCC;碳氮比由高到低依次為MCC、PCL、MCQ;MCC、MCQ的全氮含量及碳氮比均與PCL無顯著差異(圖2)。

圖2 不同林分類型土壤有機碳、全氮含量以及碳氮比Figure 2 Soil organic carbon and total nitrogen content of different stand types
2.4 植物多樣性與土壤碳氮含量的相關分析
對3種林分植物多樣性指數與土壤碳、氮含量進行相關性分析(表 3),發現喬木層Shannon-Wiener指數與土壤有機碳含量的正相關關系達到顯著水平(P<0.05);灌木層Simpson指數與土壤有機碳含量表現為顯著正相關關系,Shannon-Wiener指數、Simpson指數與全氮含量呈極顯著正相關(P<0.01);豐富度指數及與碳氮比呈顯著負相關,Shannon-Wiener指數與碳氮比為極顯著負相關,草本層和群落的植物多樣性指數均與土壤有機碳、全氮含量及碳氮比無顯著相關關系(P>0.05)。

表3 植物多樣性指數與土壤碳氮含量相關分析Table 3 Correlation analysis between plant species diversity indices of tree and chemical properties
3 討論
群落的物種多樣性是群落結構和功能復雜性的度量,能較好地反映群落的穩定性(高賢明等,2001)。研究表明,營造針闊混交林,可提升林地空間資源的利用率,增強樹種多樣性,進而增強林地抗逆性,提高生態系統的穩定性(Jactel et al.,2018)。本研究中,從林分整體群落來看,植物多樣性指數以MCQ最高,MCC次之,PCL最低。可見,營造混交林有利于增強植物群落物種多樣性,提高抵御不良環境或外部干擾的能力(黃冬梅,2017;康永祥等,2010)。
喬木層兩種混交林香濃維納指數、辛普森指數、Pielou均勻度指數均顯著高于PCL(P>0.05),豐富度指數無顯著差異(P<0.05),說明與混交林相比,PCL喬木層物種組成較為單一,分布不均。混交林灌木層、草本層的植物多樣性指數與PCL的差異不顯著,與李海霞等(2010)、馮健等(2021)研究結果相似。灌木層植物多樣指數以MCQ最高,原因是混交樹種大葉櫟樹冠濃密,林下透光度較小,更有利于耐蔭灌木及喬木幼樹幼苗的生長。草本層香濃維納指數、辛普森指數、Pielou均勻度指數以PCL最低,這是由于杉木純林地表針葉枯落物較多且難以分解,導致土壤養分下降,使一些草本植物難以在杉木純林林下定居(王玉杰等,2020)。草本層均以禾本科和蕨類植物為主要優勢種,一方面,禾本科草本植物起源于熱帶森林或林緣開放的生境,對環境適應能力強;另一方面,林下接受光照的面積有限,促進了蕨類等耐蔭植物的生長(張涵丹等,2021)。在3種林分類型中,不同群落層次植物多樣性指數以灌木層最高,因為灌木層的生態環境分化程度大于喬木層和草本層,不僅具有真正的灌木種類,還包括喬木的幼樹幼苗(吳迪等,2017)。此外,杉木純林及其混交林優勢種作用明顯(杉木、紅錐、大葉櫟),其他樹種生長處于劣勢,均勻度指數較低(吳迪等,2017),致使喬木層的均勻度指數低于灌木層和草本層。
土壤有機碳含量的變化與樹種組成、植被覆蓋度、凋落物累積與分解程度、土壤微生物、根系在土壤中的分布等因素有關(董云中等,2014)。本研究中PCL有機碳含量低于MCC和MCQ,說明營造杉闊混交林有利于提高土壤有機碳含量,與目前大部分研究結果一致(黃宇等,2005;楊玉盛等,2003;郭家新,2008)。在氣候環境基本一致的情況下,植被類型、產量和植物殘體的分解過程是有機質的重要控制因子(Ganuza et al.,2003),針葉林凋落物中含有大量難分解的有機質,影響了土壤養分的歸還速率,且針葉林林下枯落物層較厚,腐殖質層較薄,既不利于枯落物的分解,又阻擋種子插入土壤和植被發育,導致植被生長較差;而針闊混交林植被組成豐富,腐殖質層較厚,闊葉樹種往往具有更大的凋落物產量和分解速率,使其養分歸還量和歸還速率提高,從而提高土壤中有機碳的含量(孫浩等,2020)。兩種混交林土壤全氮含量與PCL差異不顯著,與前人的研究結果相似(肖石紅等,2018;馮健等,2021)。由于MCQ中含有固氮樹種——格木,通過自身固氮減少對土壤的氮吸收,并可能將固定的氮素轉移到鄰近植物,降低土壤氮消耗(Yao et al.,2019),而且格木具有一定的優勢(在喬木層和灌木層的重要值分別為為7.44%和4.84%,其在喬木層的重要值僅次于杉木及大葉櫟),故土壤全氮含量最高。3種林分類型碳氮比的變化范圍為 14.28—17.51,均高于中國土壤碳氮比均值11.9和世界土壤碳氮比均值13.33(郭冬艷,2013),可能是存在氮受限,影響土壤微生物分解能力(曹小玉等,2019)。
本研究中,土壤有機碳含量與喬木、灌木層的多樣性指數呈正相關關系,且與 Shannon-Wiener指數及灌木層Simpson指數的相關性達到顯著水平(P<0.05);全氮含量與喬木層表現為正相關關系,與灌木層Shannon-Wiener指數、Simpson指數呈極顯著正相關(P<0.01),灌木層豐富度指數和Shannon-Wiener指數與碳氮比呈顯著負相關關系,說明全氮、有機碳與杉木林植物多樣性相互作用、相互影響。因此,適當施加氮肥,可促進喬木、灌木層植物多樣性的提高(舒韋維等,2021)。地上凋落物和地下根系的分解是土壤養分的重要來源(謝君毅等,2021)。喬木、灌木層植物多樣性指數與土壤碳、氮含量相關性高于草本層,相較于草本,灌木和喬木的根系更為發達,凋落物更多,且該研究區域相對較開闊,呈現偏陽性的環境,有利于動、植物殘體在微生物作用下分解(程瑞梅等,2010),故喬木、灌木層對土壤有機碳、氮含量的影響更大。群落植物多樣性指數與土壤有機碳、全氮含量及碳氮比無顯著關系(P>0.05),因為植物生長需要消耗、利用土壤中的有機碳和全氮,而植物的調落物分解以及根系分泌小分子物質會改善土壤的有機碳和全氮含量,導致土壤中碳、氮含量變化與植物多樣性關系不顯著。此外,隨著人工林系統的生長發育,及林下植被群落的不斷演替更新,植被層對養分的消耗及改良作用之間的關系變得更加復雜,也是土壤碳、氮含量與植被多樣性相關性不顯著的主要原因之一(王春香,2014)。
4 結論
杉木與紅錐、大葉櫟混交都不同程度地提高了林分植物多樣性,有利于杉木人工林可持續發展,且 MCQ植物多樣性最高,抵御不良環境和外部干擾的能力最強(表 1)。兩種混交林土壤有機碳均高于PCL,以MCQ最高。喬木、灌木層植物多樣性指數與土壤有機碳、氮含量及碳氮比的相關性表明有機碳、氮與杉木林植物多樣性相互作用、相互影響。喬木、灌木層植物多樣性指數與土壤碳、氮含量相關性高于草本層,群落植物多樣性指數與土壤有機碳、全氮含量及碳氮比無顯著關系,可見植物多樣性及土壤碳、氮含量不僅與混交樹種的選擇有關,同時也可能受其他因素影響。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
