黍子矮稈突變體‘87’表型及對赤霉素敏感性分析
2022-05-19 00:40:46郭英杰劉曉婕許晨晨朱學(xué)海
郭英杰 劉曉婕 張 帥 劉 洋 張 萍 許晨晨 朱學(xué)海*
(1.張家口市農(nóng)業(yè)科學(xué)院 黍子研究所,河北 張家口 075032;2.中國農(nóng)業(yè)大學(xué) 農(nóng)學(xué)院,北京 100193)
黍子是一年生禾本科作物,具有生育周期短、抗旱耐瘠、耐鹽堿和抗風(fēng)沙等特點(diǎn),是我國干旱半干旱地區(qū)主要的糧食作物之一,同時(shí)在鹽堿地開發(fā)利用、沙漠治理以及晚播救災(zāi)等方面也發(fā)揮著重要作用[1-3]。此外,由于黍子中含豐富的粗蛋白、膳食纖維、常量元素和微量元素,作為一種健康食品已被消費(fèi)者廣泛關(guān)注[4]。但目前我國黍子產(chǎn)業(yè)的發(fā)展相對于其他作物較緩慢,主要體現(xiàn)在生產(chǎn)技術(shù)水平較低,仍沿襲傳統(tǒng)種植模式,耕作管理粗放,機(jī)械化程度低,另一方面種質(zhì)資源的改良優(yōu)化投入較少,種植品種仍以傳統(tǒng)農(nóng)家種為主,品種混雜、退化嚴(yán)重且產(chǎn)量低[5-7],因此通過選育新品種來實(shí)現(xiàn)黍子優(yōu)質(zhì)高產(chǎn)和適應(yīng)未來機(jī)械化種植勢在必行。
由于誘變育種可以在短時(shí)間內(nèi)獲得一些常規(guī)育種難以獲得的新突變和新種質(zhì),并極大地拓寬遺傳變異范圍,豐富突變體資源庫,因此誘變育種已成為常規(guī)選擇育種和雜交育種的重要補(bǔ)充,得到廣泛應(yīng)用[8-9]。劉美桃等[10]利用EMS誘變獲得了谷子抗除草劑‘拿捕凈’株系;陳天子等[11]通過EMS誘變獲得了水稻非轉(zhuǎn)基因抗除草劑種質(zhì);Kuraparthy等[12]用EMS誘變二倍體小麥并成功克隆到控制分蘗的TIN3基因;Brunelle等[13]利用EMS處理玉米植株后發(fā)現(xiàn)玉米EMB基因座。以上研究為育種提供了寶貴的種子資源。EMS誘變具有突變率高、變異譜廣、染色體畸變率較小、誘變成本低和操作簡單便利等優(yōu)點(diǎn),已成為挖掘水稻和小麥等多種作物基因資源的有力工具,受到國內(nèi)外學(xué)者青睞[14]。目前國內(nèi)外關(guān)于EMS誘變黍子生理的研究鮮見報(bào)道。本研究通過EMS誘變野生型黍子構(gòu)建突變體庫,并從中篩選到穩(wěn)定遺傳的矮稈突變體‘87’,通過對矮稈突變體‘87’的表型及赤霉素敏感性分析,旨在探究黍子突變體矮化生理機(jī)制,以期為改良黍子株型和培育抗倒高產(chǎn)新品種提供參考。
1 材料與方法
1.1 試驗(yàn)材料
野生型黍子材料選取‘石湖千斤糜’,國編號‘5711’,在張家口市種植編號為‘260’(野生型‘260’)及其經(jīng)0.4%(w/v) EMS誘變20 h獲得的穩(wěn)定遺傳的突變體,暫命名為矮稈突變體‘87’。試驗(yàn)材料均來自河北省張家口市農(nóng)業(yè)科學(xué)院黍子研究所。
1.2 試驗(yàn)方法
農(nóng)藝性狀調(diào)查:試驗(yàn)于2017年6月在張家口市農(nóng)業(yè)科學(xué)院沙嶺子試驗(yàn)站(40°41′ N,114°54′ E,海拔587 m)進(jìn)行。每個(gè)材料種植2小區(qū),小區(qū)長20 m,寬6 m,株距15 cm,行距30 cm,肥水管理措施同一般生產(chǎn)大田。出苗后在每個(gè)試驗(yàn)小區(qū)隨機(jī)選取野生型‘260’和矮稈突變體‘87’植株各10株,并掛牌標(biāo)記。成熟期測定掛牌植株的株高、節(jié)間長、基部第一節(jié)莖粗以及穗部等性狀。
主莖剖面細(xì)胞形態(tài)學(xué)觀察:選取抽穗期矮稈突變體‘87’和野生型‘260’第5節(jié)間的中間部位作為橫、縱向切片材料,委托武漢塞維爾生物技術(shù)有限公司進(jìn)行組織軟化、包埋、染色、切片制作與細(xì)胞觀察。
內(nèi)源GA含量測定:在抽穗期和成熟期分別采集野生型‘260’和矮稈突變體‘87’的劍葉,采用酶聯(lián)免疫吸附法(ELISA) 測定抽穗期GA3和GA4含量,成熟期的GA1+GA3含量。
外源GA敏感性分析:在種植小區(qū)內(nèi)挑選10株長勢一致的矮稈突變體‘87’植株,選取其中5株從7月19日開始每隔7 d噴施1次50 mg/L的外源GA3(外源GA3為美國Sigma公司產(chǎn)品),另外5株每次噴施等量清水作為對照,共噴施3次,噴施程度為處理液充分而又均勻分布于整個(gè)植株表面,此外每次噴施前測量植株高度。
數(shù)據(jù)處理與分析:試驗(yàn)數(shù)據(jù)采用Microsoft Excel 2013進(jìn)行數(shù)據(jù)整理、作圖,采用IBM SPSS Statistics 24軟件進(jìn)行統(tǒng)計(jì)分析。
2 結(jié)果與分析
2.1 矮稈突變體‘87’的表型差異分析
由圖1可知,矮稈突變體‘87’拔節(jié)期的株高、莖節(jié)數(shù)和分蘗數(shù)遠(yuǎn)低于野生型‘260’。由表1可知,成熟期矮稈突變體‘87’的株高僅為野生型‘260’的46.7%,分蘗數(shù)增加1.7個(gè),差異均達(dá)顯著水平;莖節(jié)數(shù)和野生型‘260’差異不顯著;矮稈突變體‘87’地上第一節(jié)莖粗比野生型‘260’增加14.1%。

表1 野生型‘260’和矮稈突變體‘87’成熟期株型結(jié)構(gòu)Table 1 The plant structure of wild-type ‘260’ and dwarf mutant ‘87’ at mature stage

圖1 拔節(jié)期矮稈突變體‘87’和野生型‘260’株型比較Fig.1 Comparison of plant type between dwarf mutant ‘87’ and wild-type ‘260’ at jointing stage
由圖2可知,矮稈突變體‘87’和野生型‘260’節(jié)間長度變化趨勢基本一致,均表現(xiàn)為隨莖節(jié)數(shù)的增加節(jié)間長度先上升后下降,地上第4莖節(jié)節(jié)間長度最大。矮稈突變體‘87’基部第1莖節(jié)莖長僅為野生型的32.3%,第2~5節(jié)長為野生型的40.0%左右,第6~9節(jié)長為野生型的45.7%~49.2%。以莖節(jié)長度比值作線性趨勢線后可知矮稈突變體‘87’基部縮短較多,莖節(jié)上部縮短較少。
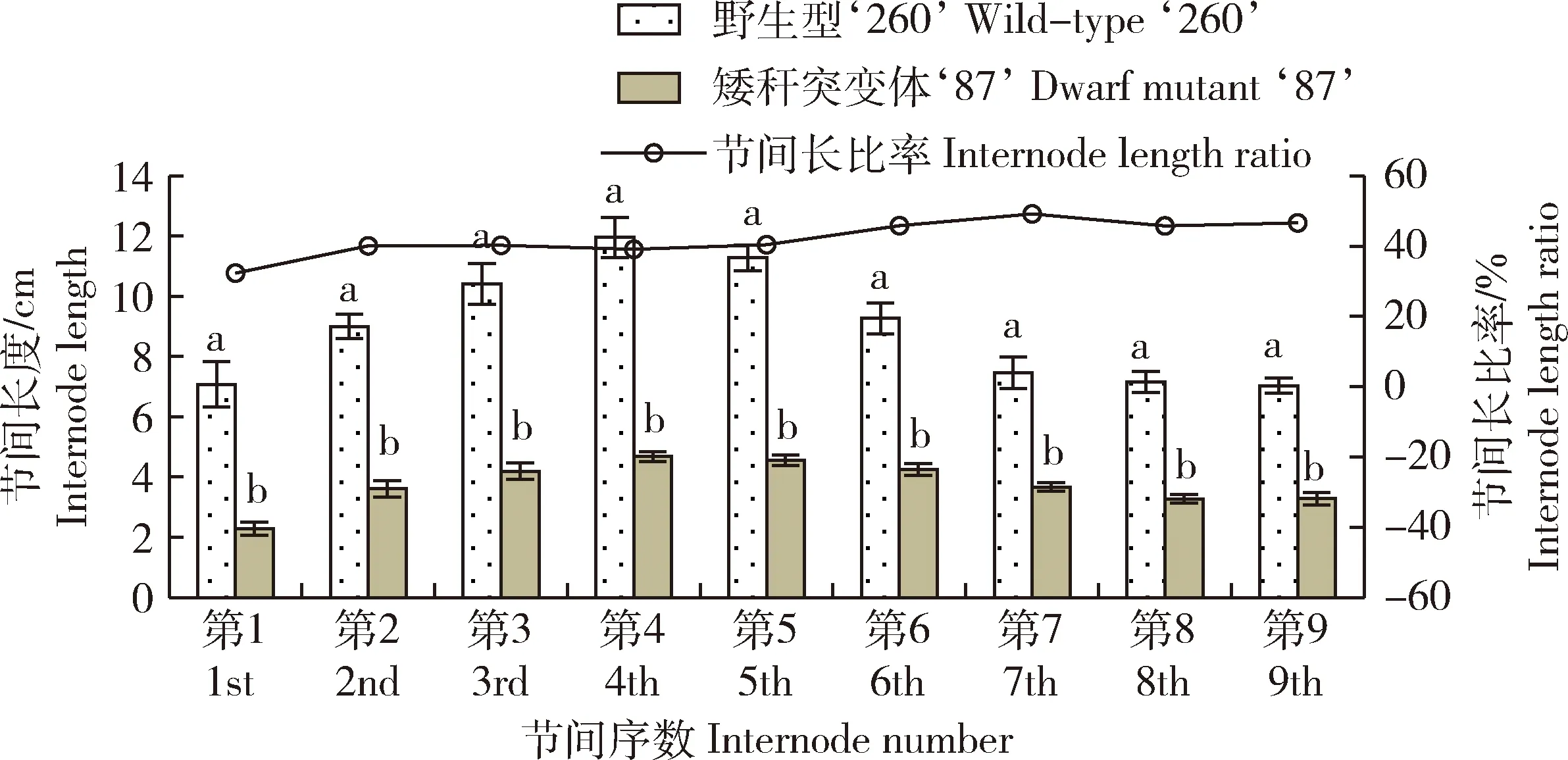
不同小寫字母表示在0.05水平上差異顯著。下同。Different lowercase letters indicate significant differences at the 0.05 level. The same below.圖2 成熟期矮稈野生型‘260’和突變體‘87’節(jié)間長度及節(jié)間比率Fig.2 Internode length and ratio of wild-type ‘260’and dwarf mutant ‘87’ at mature stage
比較野生型‘260’和矮稈突變體‘87’成熟期穗部性狀可知,矮稈突變體‘87’的穗柄長度、穗長和千粒重相比于野生型‘260’均有下降,降幅分別為54.5%、31.8%、1.7%,見表2。
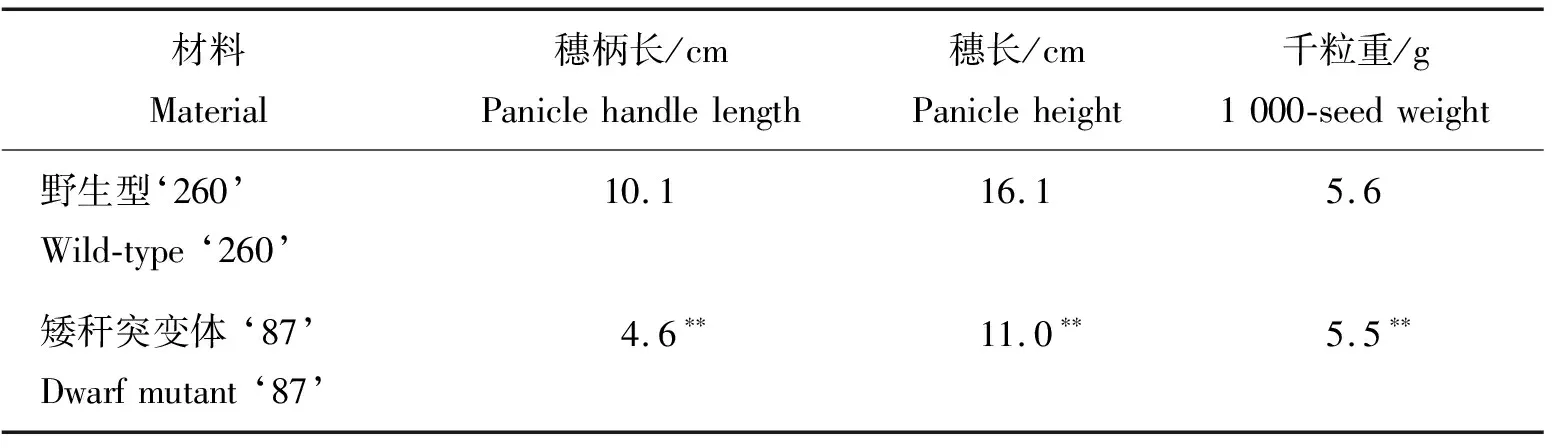
表2 野生型‘260’和矮稈突變體‘87’成熟期穗部性狀Table 2 Panicle characters of wild-type ‘260’ and dwarf mutant ‘87’ at mature stage
2.2 主莖剖面細(xì)胞形態(tài)學(xué)觀察分析
由圖3可知, 野生型‘260’和矮稈突變體‘87’縱切細(xì)胞排列均較為規(guī)則,但矮稈突變體‘87’地上第五節(jié)間中5個(gè)連續(xù)縱向細(xì)胞長度為341.0 nm,明顯小于野生型‘260’的長度,這表明矮稈突變體‘87’主莖的節(jié)間較短是由于細(xì)胞縱向長度減小所致。矮稈突變體‘87’地上第五節(jié)間中5個(gè)連續(xù)橫向細(xì)胞長度為320.0 nm,野生型‘260’的5個(gè)連續(xù)縱向細(xì)胞長度為316.5 nm(圖3(c)和(d)),說明矮稈突變體‘87’和野生型‘260’的莖稈橫切面的細(xì)胞長度基本一致,因此推測矮稈突變體‘87’莖粗的增加可能是由于單位面積細(xì)胞數(shù)目增加導(dǎo)致。
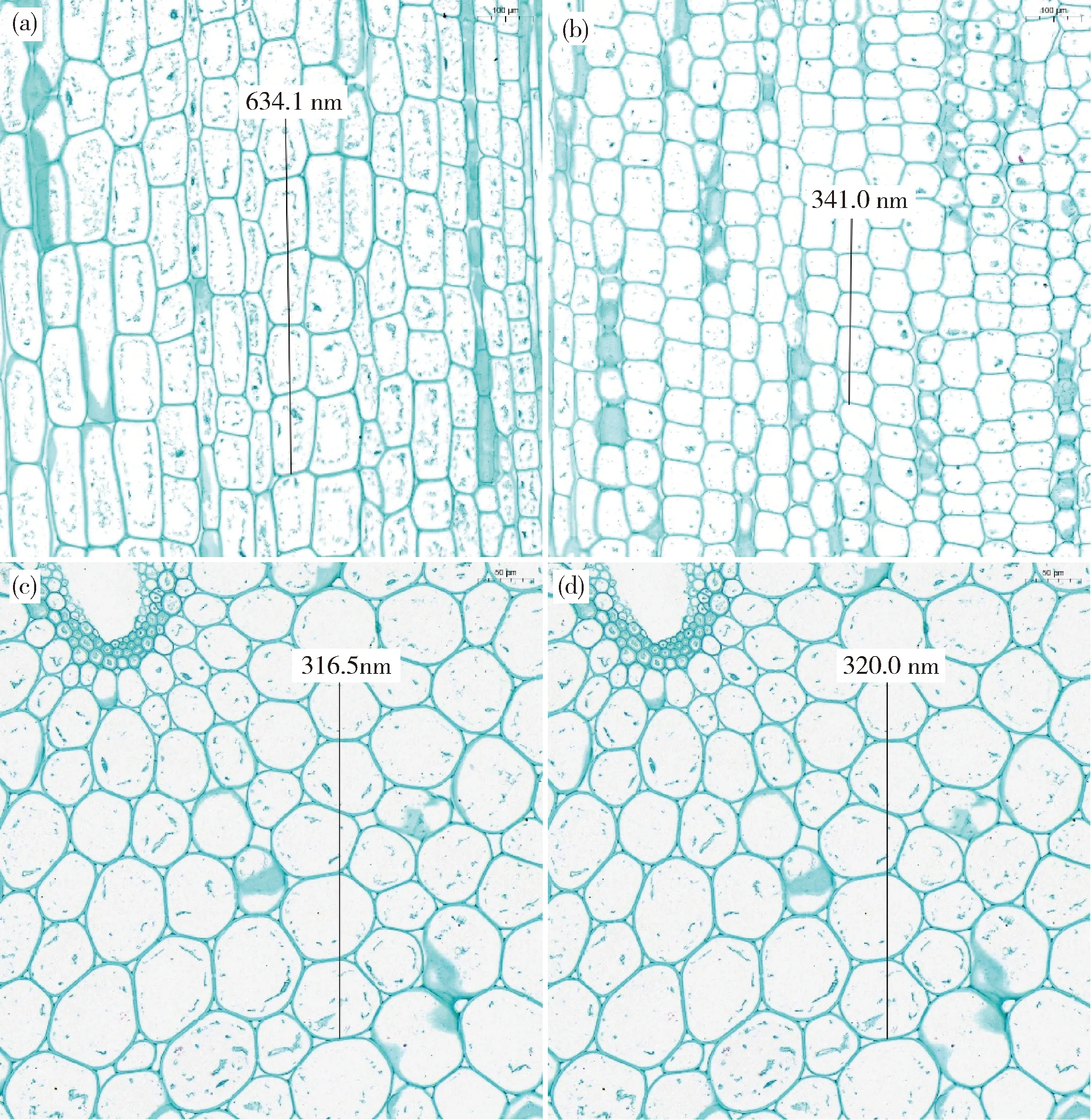
(a)和(c)分別為野生型‘260’地上第五莖間縱和橫剖面結(jié)構(gòu);(b)和(d)分別為矮稈突變體‘87’地上第五莖間縱和橫剖面結(jié)構(gòu)(a) and (c) are the longitudinal and transverse cross-sectional structure of the fifth aboveground stem of the wild-type ‘260’,respectively; (b) and (d) are the longitudinal and transverse cross-sectional structure of the fifth aboveground stem of the dwarf mutant ‘87’,respectively.圖3 成熟期野生型‘260’和矮稈突變體‘87’地上第五莖間縱、橫剖面結(jié)構(gòu)比較Fig.3 Comparison on the longitudinal and transverse cross-sectional structure of the fifth stem aboveground between wild-type ‘260’ and dwarf mutant ‘87’ at mature stage
2.3 矮稈突變體‘87’對外源GA3敏感性分析
通過不同時(shí)期對矮稈突變體‘87’噴施外源GA3及清水,并測量植株高度后可以看出,2種處理植株高度均增加,同一測定日期2種處理的株高差異不顯著(圖4)。相較于07-19株高,07-26和08-02噴施外源GA后株高增長率分別為16.0%和47.4%;噴施清水后株高增長率分別為24.1%和48.6%,2種處理株高的增長率相近,可見矮稈突變體‘87’對GA3反應(yīng)不敏感。
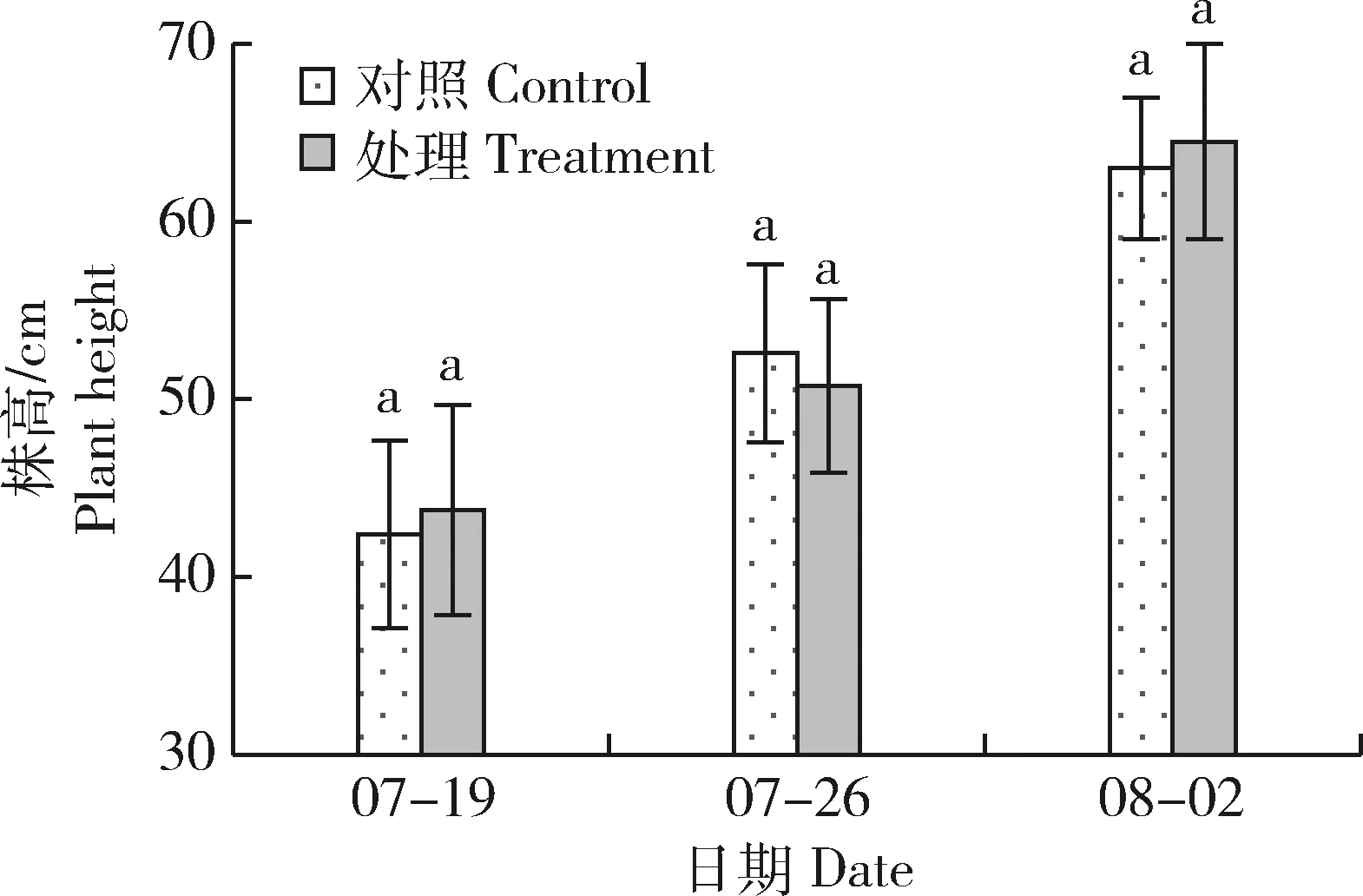
圖4 外源GA3對矮稈突變體‘87’株高的影響Fig.4 The effect of exogenous GA3 on plant height of dwarf mutant ‘87’
2.4 內(nèi)源GA含量分析
由圖5可知,抽穗期矮稈突變體‘87’和野生型‘260’內(nèi)源GA4含量分別高于GA3含量8.1%和5.6%,且矮稈突變體‘87’內(nèi)源GA3含量和GA4含量均高于野生型‘260’,但均未達(dá)顯著水平;成熟期,矮稈突變體‘87’內(nèi)源GA1+GA3含量比野生型‘260’增加14.4%。綜上,矮稈突變體‘87’的矮化性狀可能與內(nèi)源GA含量增高有關(guān)。
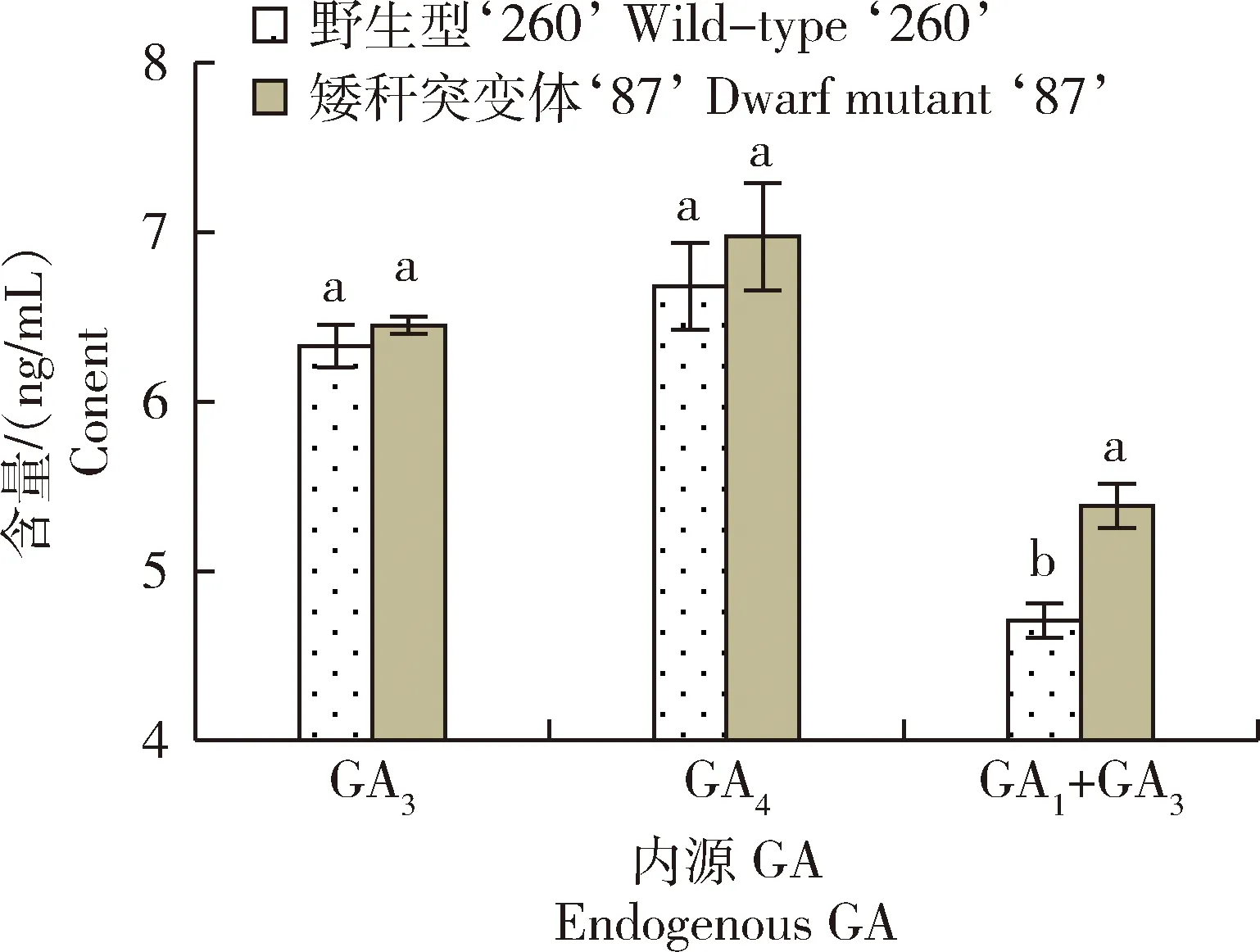
內(nèi)源GA3和內(nèi)源GA4測定時(shí)期為抽穗期,內(nèi)源GA1+GA3測定時(shí)期為成熟期。The measurement period of endogenous GA3 and endogenous GA4 is the heading period, and the measurement period of endogenous GA1+GA3 is the mature period.圖5 野生型‘260’和矮稈突變體‘87’內(nèi)源GA含量測定Fig.5 Determination of endogenous GA content in wild-type ‘260’ and dwarf mutant ‘87’
3 討 論
黍子是非常容易發(fā)生倒伏的禾本科作物,加之近年黍子產(chǎn)區(qū)降雨增多、溫度升高等極端天氣的出現(xiàn),倒伏已成為黍子生產(chǎn)中存在的普遍問題,嚴(yán)重影響了產(chǎn)量[15]。大量研究表明,株高、節(jié)間數(shù)和節(jié)間長度等因素與倒伏有極顯著相關(guān)關(guān)系[16-17],這是因?yàn)橹旮吲c地上部鮮重呈顯著正相關(guān),地上部鮮重越大,莖稈基部的彎曲力矩越大,越容易發(fā)生倒伏[18]。但也有一些研究者認(rèn)為株高并不是影響倒伏的主要因素,在一定株高范圍內(nèi),株高與倒伏并沒有必然的聯(lián)系,而莖稈基部粗度及充實(shí)度的貢獻(xiàn)更大[19-20]。本研究利用EMS誘變獲得的矮稈突變體‘87’節(jié)間數(shù)與野生型‘260’基本一致,而地上各節(jié)間長度和株高均顯著低于野生型‘260’,且地上第五節(jié)間的縱向細(xì)胞長度顯著低于野生型‘260’,可見矮稈突變體‘87’的株高降低是由于細(xì)胞縱向長度減小使節(jié)間長度下降所致,這一結(jié)果與Yang等[21]在矮稈甜瓜上的研究結(jié)論一致;相較于野生型‘260’,矮稈突變體‘87’基部莖粗增加,且莖稈橫切片單位面積內(nèi)細(xì)胞數(shù)目增加,從而使矮稈突變體‘87’的莖稈力學(xué)性能強(qiáng)于野生型‘260’[22],正是矮稈突變體‘87’株高和莖稈力學(xué)性能上的改善,使其抗倒伏性優(yōu)于野生型‘260’。
矮稈突變體植株常常由于赤霉素的生物合成或信號傳導(dǎo)缺陷在表型上表現(xiàn)出矮化、分蘗增多、葉色變深、開花推遲、花發(fā)育異常、產(chǎn)量及產(chǎn)量構(gòu)成下降等變異[23-25],這和本研究中獲得的矮稈突變體‘87’表型變化基本一致,成熟期矮稈突變體‘87’與野生型‘260’相比,株高下降、分蘗數(shù)增加,穗長、千粒重顯著下降。通過對矮稈突變體‘87’施加外源激素及內(nèi)源激素含量測定發(fā)現(xiàn),矮稈突變體‘87’在施加外源GA3后,株高變化與對照清水處理基本一致,且抽穗期內(nèi)源GA3和GA4含量及成熟期內(nèi)源GA1+GA3含量均高于野生型‘260’,其中成熟期內(nèi)源GA差異達(dá)顯著水平。這一結(jié)果與GA3鈍感型植株研究結(jié)果一致,當(dāng)施加外源GA后,植株表型沒有明顯響應(yīng),不能恢復(fù)野生型表型[26],且內(nèi)源活性GA水平高于野生型或與野生型相當(dāng)[27-28]。因此,推斷矮稈突變體‘87’是由于GA合成或信號轉(zhuǎn)導(dǎo)途徑發(fā)生變化導(dǎo)致植株矮化,是一個(gè)GA不敏感性矮化突變體,有待進(jìn)一步研究驗(yàn)證。
4 結(jié) 論
本研究結(jié)果表明,矮稈突變體‘87’由于莖稈縱向細(xì)胞數(shù)目減少,節(jié)間長度下降,使其株高降低;由于莖稈橫向數(shù)目增加,使其莖稈變粗,從而增強(qiáng)抗倒伏性能。抽穗期和成熟期突變體‘87’植株內(nèi)源活性GA水平均高于野生型。因此,矮稈突變體‘87’可以作為黍子矮化育種的種質(zhì)材料。