不同種植模式下鳳丹根際微生物群落結構和多樣性分析
2022-05-16 23:40:31劉智張虹羅會婷王仲偉湯詩杰
江蘇農業學報 2022年2期
劉智 張虹 羅會婷 王仲偉 湯詩杰

摘要:為探究不同種植模式下鳳丹根際微生物群落結構和多樣性,采用高通量測序技術對采自櫸樹(Zelkova serrata)下種植(ZS)、娜塔櫟(Quercus nattallii)下種植(QS)、露天種植(OS)的鳳丹根際以及空白土壤(CK)樣品土壤細菌和真菌進行測序。結果顯示,與空白土壤相比,種植鳳丹提高了根際微生物群落的豐富度和多樣性,不同種植模式下鳳丹根際微生物群落結構有所差異。3種種植模式下細菌的優勢菌門均為變形菌門(Proteobacteria)、酸桿菌門(Acidobacteria)、放線菌門(Actinobacteria)以及疣微菌門(Verrucomicrobia),但不同種植模式下每個門的相對豐度有所差異。所有樣品中總共檢測出12個真菌門,3種種植模式下真菌的優勢菌門均為子囊菌門(Ascomycota)和擔子菌門(Basidiomycota),不同種植模式下各真菌門的相對豐度存在差異;屬水平上真菌群落結構差異較大,ZS組、QS組和OS組的優勢菌屬分別為Exophiala、Thelephoraceae_unclassified、Ascomycota_unclassified。測定3種種植模式下鳳丹的生長指標,發現林下種植的鳳丹株高、葉長、葉寬均高于露天種植。林下種植模式的細菌需氧表型、革蘭氏陰性表型、生物膜形成表型以及致病潛力表型高于單一種植模式。本研究從根際微生物角度分析了不同種植模式對鳳丹的影響,為鳳丹林下種植提供參考,為后續促生菌篩選、菌肥研制提供理論支撐。
關鍵詞:鳳丹;根際微生物;高通量測序;微生物多樣性;微生物群落結構
中圖分類號:S154.3文獻標識碼:A文章編號:1000-4440(2022)02-0502-10
Analysis on community structure and diversity in rhizosphere microorganisms of Paeonia ostii under different planting patterns
LIU Zhi ZHANG Hong LUO Hui-ting WANG Zhong-wei TANG Shi-jie
Abstract:In order to explore the rhizosphere microbial community structure and diversity of Paeonia ostii under different planting patterns, high-throughput sequencing technology was used to sequence the rhizosphere soil samples of Paeonia ostii collected from three different planting patterns, namely Zelkova serrata (ZS), Quercus nattallii (QS) and open-air planting (OS) and blank soil(CK). The results showed that compared with the blank soil, planting Paeonia ostii increased the richness and diversity of rhizosphere microbial community, and the rhizosphere microbial community structure of Paeonia ostii was different under different planting patterns. The dominant phyla of bacteria under three planting patterns were Proteobacteria, Acidobacteria, Actinobacteria and Verrucomicrobia, but the relative abundance of each phylum under different planting patterns was different. A total of 12 fungal phyla were detected in all samples. The dominant phyla of fungi under three planting modes were Ascomycota and Basidiomycota, and the relative abundance of each fungal phylum was different under different planting patterns. The fungal community structure was quite different at genus level, and the dominant genera in ZS group, QS group and OS group were Exophiala, Thelephoraceae_unclassified and Ascomycota_unclassified, respectively. The growth indices of Paeonia ostii under three planting patterns were measured, plant height, leaf length and leaf width of Paeonia ostii planted under forest were higher than those planted in open field. The aerobic phenotype, Gram-negative phenotype, forms biofilms phenotype and potentially pathogenic phenotype in cultivation pattrerns under forest were higher than those in single planting mode. In this study, the effects of different planting patterns on Paeonia ostii were analyzed from the perspective of rhizosphere microorganisms, so as to provide reference for understory planting of Paeonia ostii, and provide theoretical support for subsequent screening of probiotics and development of bacterial fertilizer.
Key words:Paeonia ostii;rhizosphere microorganisms;high-throughput sequencing;microbial diversity;microbial community structure
鳳丹(Paeonia ostii)屬芍藥科芍藥屬植物,結實能力強,鳳丹籽產量和出油率高。鳳丹籽油富含多種不飽和脂肪酸,其干燥根皮被稱為丹皮,可入藥,花兼具觀賞、食用和藥用價值,在全國范圍內得到了廣泛的種植[1-4]。國家發展和改革委員會等部門于2020年印發的《關于科學利用林地資源促進木本糧油和林下經濟高質量發展的意見》指出,到2025年,要新增或改造木本糧油經濟林3.33×106 hm2,木本食用油年產量達2.5×106 t,林下經濟年產值達1×1012 元。作為多年生灌木,鳳丹抗逆性強、適應性好,可林下種植,與核桃、香椿、油茶等作物已經形成了較為成熟的套種模式。林下種植充分利用了土地資源,同時發揮了林地的林蔭優勢,改善了鳳丹的生長環境,對提高產量、促進增收有著積極意義[5]。
根際是植物根系與土壤微生物相互作用的場所,具體指植物的根表及受根系直接影響的土壤區域,是地球上最復雜的生態系統之一[6-7]。植物的根系分泌物為根際微生物的生長提供營養物質,大約有10%的凈光合固定碳通過分泌物的形式釋放到根際,從而形成不同的根際微生物群落[8]。不同的根際微生物可以通過不同的方式,比如調控植物激素水平、影響植物信號的產生與傳導、改善營養獲取方式以及提高植物抗逆性等來影響植物的生長發育[9-11]。一些根際微生物可以通過形成生物膜來幫助植物抵抗高溫和高鹽脅迫[12],枯草芽孢桿菌(Bacillus subtilis)可以分泌激素類和抗菌素類物質來抵抗病原菌[13]。根際微生物群落結構受到植物類型的影響,楊瀟湘等[14]比較油菜和大豆根際土壤微生物群落結構發現,兩者在門水平上的優勢菌群相同,但是后者富含具有生防作用的微生物,比如黃桿菌屬(Flavobacterium)、鞘脂單胞菌屬(Sphingomonas)、芽孢桿菌屬(Bacillus)等;大豆根際具有更多的病原細菌和真菌,比如腸桿菌(Enterobacter)和炭疽菌(Colletotrichum spp.)等。除此之外,土壤環境、栽培模式、水肥條件、田間管理模式等也會影響植物根際微生物結構。例如劉振香等[15]研究發現,夏玉米田中施用有機肥可以提高土壤微生物多樣性和豐富度;姜莉莉等[16]研究發現,蘋果園內種植黑麥草可提高土壤中鏈格孢菌(Alternaria)和曲霉菌(Aspergillus)的相對豐度,種植白三葉可提高土壤赤霉菌(Gibberella)的相對豐度,而自然生草則會提高青枯菌(Ralstonia)等致病菌的相對豐度。
相比于傳統土壤微生物研究方法,高通量測序技術所需樣本量少、能用環境樣本中直接獲取的總DNA進行文庫的構建,操作簡單、準確度高,成為研究土壤微生物的熱門手段[17-19]。前人已經研究了不同地區[20]、不同種植年限[21-22]鳳丹根際微生物多樣性,但對于不同種植模式下鳳丹根際微生物多樣性的研究還未見報道。本研究基于高通量測序技術,研究不同種植模式下鳳丹根際微生物群落結構和多樣性,以期為鳳丹根際微生態環境研究提供理論基礎,為優化油用牡丹栽培技術提供思路。
1材料與方法
1.1試驗材料
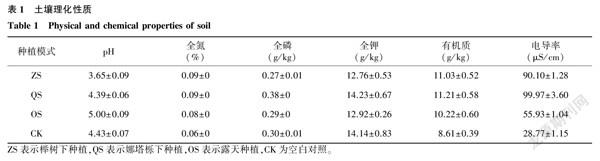
試驗地位于江蘇省中國科學院植物研究所試驗田(南京市六合區竹鎮鎮,32°33′46″N,118°39′28″E),土壤類型均為黃棕壤,屬亞熱帶季風氣候。試驗田內設有櫸樹下種植(ZS)、娜塔櫟下種植(QS)以及露天種植(OS)3種模式,櫸樹和娜塔櫟的株距×行距為4 m×6 m。3種模式下種植的鳳丹均為3年生苗,株距×行距為40 cm×60 cm,種植密度約為1 hm2 42 000株,水肥管理均參照國家林業局《油用牡丹栽培技術規程》[23]進行。于2021年8月采樣,每個試驗區均采用五點采樣法,挖出整株鳳丹植株,采用抖根法采集根際。按照相同方法,在空地上采集地表10~15 cm深度的土壤作為對照組(CK),每組樣品均設6個生物學重復。收集的樣品裝入無菌自封袋,冰盒保存,帶回實驗室備用。土壤理化性質見表1。
1.2鳳丹生長指標測定
生長指標于2021年6月底測定,每個小區隨機選取10株鳳丹植株,使用卷尺(精度0.1 cm)測量株高,每1株上選取3片健康成熟頂生小葉,使用直尺(精度0.1 cm)測量葉長(葉片基部至頂端長度)和葉寬(葉片最寬處寬度),重復3次,求平均值。
1.3根際微生物基因組DNA的提取、擴增以及高通量測序
使用天根DP712試劑盒(天根生化科技有限公司產品)提取根際微生物基因組DNA,采用1%瓊脂糖凝膠電泳檢測其完整性,采用分光光度計檢測其濃度和純度。使用引物F:5′-CCTACGGGNGGCWGCAG-3′,R:5′-GACTACHVGGGTATCTAATCC-3′擴增細菌的16 S rDNA的V3+V4區;使用引物F:5′-GTGARTCATCGAATCTTTG-3′,R:5′-TCCTCCGCTTATTGATATGC-3′擴增真菌的ITS2區域。PCR反應體系為:Phusion Hot Start Flex 2×Master Mix 12.5 μl,1 μmol/L的正反向引物各2.5 μl,DNA模板50 ng,加ddH2O至25.0 μl。PCR反應條件為:98 ℃ 30 s;98 ℃ 10 s,54 ℃ 30 s,72 ℃ 45 s,35個循環;72 ℃ 10 min。
PCR產物經純化和定量之后構建DNA文庫,評估合格的測序文庫(由聯川生物科技有限公司提供)在Illumina NovaSeq平臺上進行測序。
1.4數據處理
使用軟件Cutadapt(v1.9)去除barcode標簽和引物序列,使用Fqtrim(v0.94)和Vsearch(v2.3.4)分別過濾低質量序列和嵌合體,利用DADA2進行解調,得到特征(feature)序列和特征豐度表格。
對特征豐度進行歸一化后,使用QIIME2計算樣品的α多樣性指數(Chao1指數、Observed species指數、Goods coverage指數、Shannon指數、Simpson指數)和β多樣性指數。使用SILVA數據庫(https://www.arb-silva.de/documentation/release138/)注釋特征序列,獲取分類學信息,使用Excel 2016和SPSS 26處理數據。將樣本信息上傳至BugBase(https://bugbase.cs.umn.edu/),預測細菌表型。使用R(VennDiagram)繪制特征分布Venn圖,使用R(vegen 2.5.4)繪制PCA圖,使用Origin Pro 2021繪制箱線圖和群落結構圖。
2結果與分析
2.1測序結果質量評估
原始數據處理后,統計高質量數據。16 S rDNA測序序列個數為1 717 754,有99.88%的序列長度在400~500 bp,有效數據與原始數據之比(Valid)均值為85.31%,其中最大值為94.01%,最小值為73.87%;所有樣本的Q30均在80.00%以上,平均值為91.76%。
ITS2測序序列個數為1 969 418,有70.10%的序列長度在200~300 bp,29.18%的序列長度在300~400 bp,Vaild最小值為94.26%,Q30均在95%以上。說明,此次測序質量良好。
2.2不同種植模式下鳳丹根際微生物豐富度和多樣性
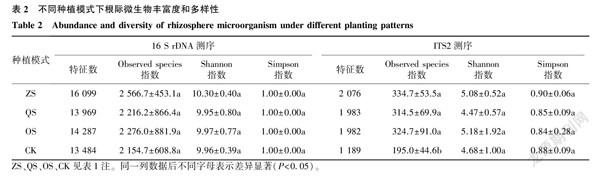
與傳統操作分類單元(OTU)相比,DADA2算法可以對擴增子數據進行去噪、修正、去冗余,得到特征數據,數據精確度和物種分辨率大大提升。由表2可見,在16 S rDNA和ITS2測序中,ZS組的特征數最多,QS組和OS組次之,CK的特征數最少。
α多樣性指數可以用來反映樣本的物種豐富度和多樣性。在16 S rDNA測序中,4組的Observed species指數之間無顯著差異;4組的Simpson指數均為1.00。在ITS2測序中,ZS、QS、OS組的Observed species指數均高于CK; 各組間Simpson指數和Shannon指數差異不顯著。
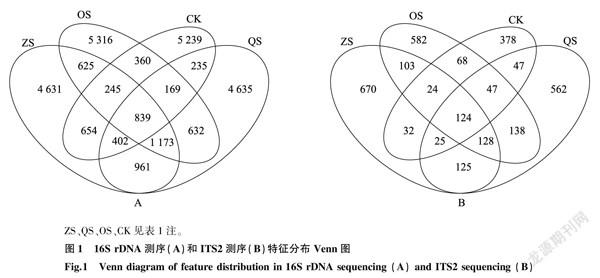
從圖1可以直觀地看出,每組樣品共有和特有的特征數目。在16 S rDNA測序中,4組樣品共有的特征數為839,ZS組、QS組、OS組、CK特有的特征數分別為:4 631、4 635、5 316和5 239;ITS2測序中,4組樣品共有的特征數為124,ZS組、QS組、OS組、CK特有的特征數分別為670、562、582和378。
從以上結果可以看出,相比于空白土壤,3種種植模式均提高了鳳丹根際微生物的豐富度和多樣性;3種不同的種植模式中,鳳丹根際細菌和真菌的豐富度和多樣性也存在差異。
2.3不同種植模式下鳳丹根際細菌群落結構
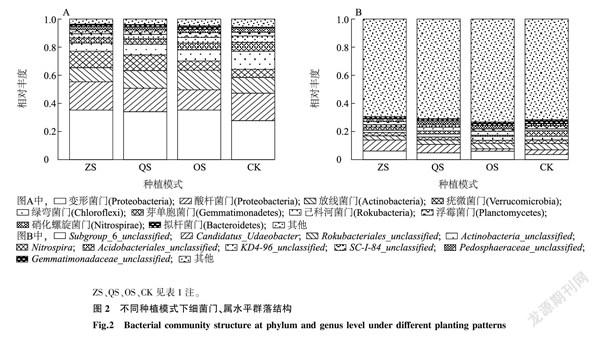
取鳳丹根際細菌門水平相對豐度前10位(TOP10)繪制柱狀堆疊圖(圖2A)。在門水平上細菌群落結構中,變形菌門(Proteobacteria)在4組中豐度最高,ZS組、QS組、OS組均高于CK;其次是酸桿菌門(Proteobacteria),在ZS組和CK中的豐度相當,均高于QS組和OS組;OS組中放線菌門(Actinobacteria)的豐度大于其他3組;疣微菌門(Verrucomicrobia)在ZS組和QS組中的豐度高于OS組和CK;擬桿菌門(Bacteroidetes)在OS組中的豐度要高于其他3組;綠彎菌門(Chloroflexi)、己科河菌門(Rokubacteria)以及硝化螺旋菌門(Nitrospirae)在CK中的豐度分別為12.9%、4.6%和3.0%,均高于在其他3組中的豐度。
圖2B是屬水平上鳳丹根際細菌群落結構,屬水平上優勢菌多為現階段未分類的細菌,在ZS組和QS組中,Candidatus_Udaeobacter的相對豐度均為最高,分別為7.80%和6.07%,其次為Subgroup_6_unclassified,相對豐度分別為6.1%和4.8%, 排在第三位的是Rokubacteriales_unclassified,相對豐度分別為3.0%和3.1%。在OS組中Subgroup_6_unclassified的相對豐度最高,之后按照相對豐度高低依次是Rokubacteriales_unclassified、KD4-96_unclassified、Actinobacteria_unclassified。在CK中,按照豐度高低依次是:Rokubacteriales_unclassified、Acidobacteriales_unclassified、Subgroup_6_unclassified、Candidatus_Udaeobacter。
由此可見,不同的種植模式對鳳丹根際細菌群落結構是有一定影響的,特別是酸桿菌門、放線菌門和疣微菌門。在屬水平上,Subgroup_6_unclassified、Candidatus_Udaeobacter和Rokubacteriales_unclassified在不同的種植模式下差異較大。
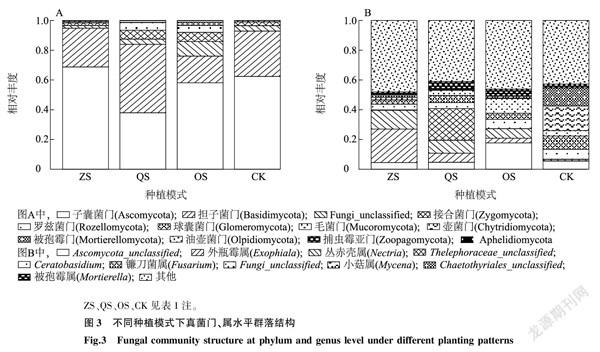
2.4不同種植模式下鳳丹根際真菌群落結構
由圖3A可以看出,在ZS組、OS組和CK中子囊菌門(Ascomycota)相對豐度最高,擔子菌門(Basidiomycota)位居第二,而在QS組中擔子菌門豐度最高。OS組中Fungi_unclassified、球囊菌門(Glomeromycota)以及接合菌門(Zygomycota)的相對豐度要高于其他3組。毛霉門(Mucoromycota)和被孢霉門(Mortierellomycota)沒有出現在CK中。
由圖3B可知,ZS組中相對豐度TOP3依次是外瓶霉屬(Exophiala)、叢赤殼屬(Nectria)、Ascomycota_unclassified,其中外瓶霉屬的相對豐度達到了22.5%;QS組中Thelephoraceae_unclassified的相對豐度最高,為21.3%,其次為叢赤殼屬;OS組中,Ascomycota_unclassified的豐度最高,其次為Fungi_unclassified;而在CK中,相對豐度TOP3為小菇屬(Mycena)(16.6%)、Chaetothyriales_unclassified(12.2%)、鐮刀菌屬(Fusarium)(9.0%)。
不同種植模式下,鳳丹根際真菌群落結構有著較大的差異。門水平上子囊菌門、擔子菌門、Fungi_unclassified、接合菌門豐度差異較大。而在屬水平上差異更明顯,每組的優勢屬均不同。
2.5不同種植模式下鳳丹根際微生物β多樣性分析
對不同種植模式下鳳丹根際細菌和真菌群落結構進行主成分分析,結果見圖4,根據樣品代表的點在PCA圖中的距離,可以判斷樣品之間的相似度。同時PCA圖給出了基于Anosim相似性分析計算出的R值,R值越接近于1,說明組間差異越大于組內差異。
從圖4A可以看出,在不同種植模式下鳳丹根際細菌群落PCA圖中,前2個主成分的累計貢獻率達到了48.58%。ZS組和QS組樣品對應的點在PCA圖中較為集中,且兩組距離較近,說明ZS組和QS組的細菌群落組成較為相似。OS組中的大部分點與ZS組、QS組分離,CK與ZS組、QS組明顯分離。結合R=0.729 5可知,不同組別之間差異較大,說明相比于CK,種植鳳丹改變了根際細菌的群落結構,2種林下種植模式的細菌群落組成較為相似,且與露天種植模式存在差異。
圖4B是不同種植模式下鳳丹根際真菌群落PCA圖,ZS組、QS組和OS組樣品對應的點在PCA圖中距離較近,說明3組的真菌群落結構較為相似,CK與其他3組有一定的分離,結合R=0.652 9可知,相比于CK,種植鳳丹改變了根際真菌群落結構。
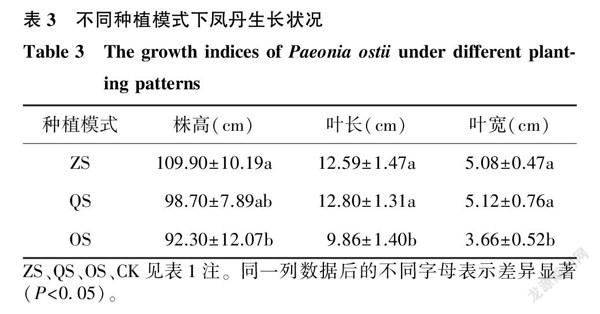
2.6不同種植模式下鳳丹的生長狀況
不同種植模式下鳳丹的生長狀況如表3所示,ZS組的株高高于QS組和OS組,其中和OS組差異顯著;ZS組和QS組的葉長均顯著高于OS組;ZS組和QS組的葉寬均顯著高于OS組。
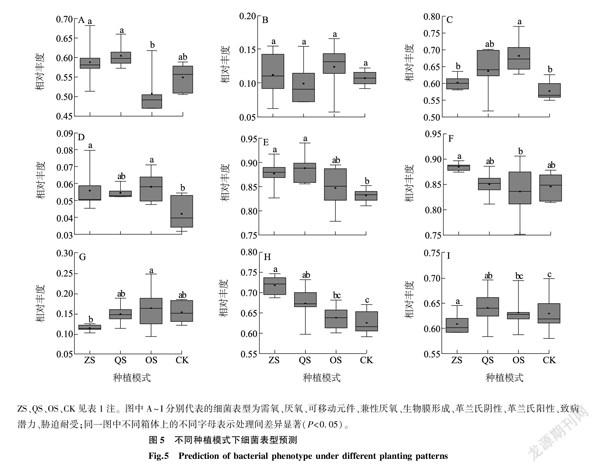
2.7不同種植模式下鳳丹根際細菌表型預測
細菌表型預測結果(圖5)表明,不同種植模式下鳳丹根際細菌表型之間存在較大差異。ZS組和QS組的需氧表型顯著高于OS組;可移動元件表型在OS組中最高,顯著高于ZS組和CK,但與QS組沒有顯著差異;兼性厭氧表型表現為ZS組、OS組高于CK, QS組和CK沒有顯著差異;ZS組、QS組的生物膜形成表型要高于CK, OS組與CK無顯著差異;革蘭氏陰性表型在ZS組、QS組和CK中無顯著差異,但ZS組顯著高于OS組;革蘭氏陽性表型則表現為OS組顯著高于ZS組;ZS組和QS組的致病潛力顯著高于CK;ZS組的脅迫耐受低于OS組和CK。厭氧表型在4組中沒有顯著差異。
3討論
研究結果表明,選取合理的種植模式能增強作物根際的交互作用,優化根際微生物群落結構,對提高土壤肥力、促進作物生長發育有著積極作用[24]。輪作、套作以及連作模式下,丹參根際微生物群落結構存在一定差異,輪作和套作對改善土壤質量以及緩解丹參連作障礙有著一定的積極作用[25]。香蕉間作韭菜、假花生和覆蓋稻草處理,顯著提高了香蕉根際放線菌的相對豐度,但是尖孢鐮刀菌(F. oxysporum)的豐度有所下降,降低了香蕉枯萎病的發生幾率[26]。探究不同種植模式下草莓根際細菌和真菌群落結構,發現限根種植提高了草莓根際放線菌門、變形菌門、擬桿菌門以及子囊菌門的豐度,降低了綠彎菌門、酸桿菌門和接合菌門的豐度[27]。研究發現,鳳丹與香椿、女貞間作提高了單株地上部分生物量和產量[28],鳳丹-香椿套種模式在定植4年后產量高出單一種植模式51.97%,極大提高了籽粒產量[29]。
與CK相比,綠彎菌門、己科河菌門以及硝化螺旋菌門在其他處理組中的相對豐度有所降低。綠彎菌多為細胞絲狀,營養方式多樣,參與CO2的同化過程[30];硝化螺旋菌門可將銨態氮氧化為硝態氮,參與土壤氮循環[31]。3種種植模式下細菌的優勢菌門均為放線菌門,這與張永敢[32]等的研究結果一致,但是相比于露天種植,林下種植鳳丹根際細菌酸桿菌門和疣微菌門的相對豐度得到了提高,芽單胞菌門、放線菌門以及擬桿菌門有所下降。相關研究結果表明,擬桿菌門在輪作休耕模式中與土壤全鉀含量呈負相關[33];而在黃花蒿連作中,隨著種植年限的增加,根際芽單胞菌門和放線菌門的豐度有所降低,說明黃花蒿能選擇性抑制它們的生長繁殖[34]。2種林下種植模式鳳丹根際細菌群落結構和多樣性也存在一定差異,相比在娜塔櫟下種植鳳丹,在櫸樹下種植鳳丹提高了根際酸桿菌門的豐度,酸桿菌門細菌能夠在缺氧酸性環境下生存,抗逆性強,對林地中樹葉等植物殘體具有降解功能[35],本研究中ZS組的pH低至3.65,也佐證了這一結論。
在所有樣品中,鑒定出的真菌分別屬于12個門,3種種植模式下真菌優勢菌門均為子囊菌門和擔子菌門,且在ZS組中子囊菌門的豐度最高。子囊菌門真菌多為腐生真菌,能分解土壤中的化合物,對土壤養分循環有著重要的作用,一定程度上可以促進作物的生長發育[36-38];擔子菌門的一些真菌可以與作物共生形成菌根,增強作物抗性[39]。屬水平上,ZS組叢赤殼屬豐度高于其他3組,被孢霉屬在QS組中豐度最高。王麗娟等[40]從鹿蹄草中分離出1株促生菌,鑒定為叢赤殼屬真菌,該菌株可以顯著提高鹿蹄草的根系活力,提高葉片中葉綠素a的含量、可溶性糖含量以及超氧化物歧化酶活性,從而促進鹿蹄草的生長發育。被孢霉屬真菌主要腐生于土壤,部分種類可以和植物共生,對改善土壤結構有一定作用,能促進植物的生長[41]。鐮刀菌屬(Fusarium)和Ceratobasidium作為常見的植物病原菌,對種子的萌發及植物根莖的生長不利,可能導致作物發生枯萎病、根腐病等[42-43],而本研究發現在ZS組中這2種真菌的豐度要低于其他3組,這說明櫸樹下種植鳳丹對抵抗病原菌有一定的作用。
BugBase是基于測序數據進行的預測分析,并不能真實反映細菌群落的全部表型,但是分析不同種植模式下鳳丹根際的細菌表型,對于鳳丹的種植仍有一定的理論參考價值。細菌表型預測結果顯示,兼性厭氧表型在CK中最低,這可能是因為隨著鳳丹種植年限增加,連作土壤總孔度減少,從而影響通氣性所導致的[44]。林下種植模式下需氧表型、革蘭氏陰性表型、生物膜形成表型以及致病潛力表型在一定程度上高于單一種植模式,這可能跟變形菌門的豐度升高有關。變形菌門細菌在自然界中廣泛存在,革蘭氏陰性細菌,大多數都具有脂多糖組成的外膜,其中β-變形菌多為好氧及兼性細菌,包含很多病原菌,這與楊睿[45]研究貴州喀斯特山區花椒林根際細菌表型的結果相符,隨著變形菌門豐度的升高,具有上述表型的細菌豐度也隨著升高。研究結果表明,放線菌門有抗脅迫的能力[46],本研究中QS組和OS組中放線菌門的相對豐度高于其他2組,脅迫耐受表型也表現出相同的變化。細菌的表型受多種因素影響,在鳳丹的種植中,不同模式下土壤理化性質存在差異,環境異質性大,微生物通過表型的適應性調整,以應對環境的改變[47]。
對果實生長期鳳丹的部分生長指標進行測定,發現套種在櫸樹下的鳳丹植株生長狀態最佳,其株高顯著高于其他2組,這可能和子囊菌門、酸桿菌門中一些有益菌的豐度增加有關。
綜上,林下種植鳳丹提高了根際微生物多樣性,不同種植模式下鳳丹根際微生物群落結構存在較大差異,櫸樹下套種鳳丹提高了酸桿菌門、子囊菌門、叢赤殼屬等有益菌的豐度,減少了鐮刀菌屬和Ceratobasidium等致病菌的豐度。測量生長指標發現櫸樹下種植的鳳丹長勢最好,由此推斷,櫸樹下種植鳳丹有助于植株的生長發育。我們今后還將從土壤代謝組學、籽粒產量以及鳳丹藥用成分等角度繼續探索不同種植模式對鳳丹的影響,以期為鳳丹高效栽培技術以及林農復合經營提供新思路。
參考文獻:
[1]張姍姍,趙凡,魏小豹,等. 鳳丹和紫斑牡丹6個產地種子脂肪酸組分的比較[J]. 中國糧油學報, 2021, 36(3): 84-90.
[2]謝一青,吳慶錐,余孟楊,等. 鳳丹南移福建的適應性分析[J]. 熱帶作物學報, 2021,42(2): 592-598.
[3]王仲偉,王歡利,張文獻,等. 江蘇油用牡丹的發展思考[J]. 江蘇林業科技, 2016, 43(6): 53-55.
[4]裴姿琛,胡永宏,劉澤,等. 新型木本油料作物——油用牡丹的開發和利用[J]. 林業與生態科學, 2018, 33(4): 358-363.
[5]王勇,何舒,熊冰杰,等. 不同栽培模式對人參根際土壤微生物多樣性的影響研究[J]. 中草藥, 2021, 52(17): 5303-5310.
[6]劉王鎖,李海泉,何毅,等. 根際微生物對植物與土壤交互調控的研究進展[J]. 中國土壤與肥料, 2020(5): 318-327.
[7]KHAN N, ALI S, SHAHID M , et al. Insights into the interactions among roots, rhizosphere, and rhizobacteria for improving plant growth and tolerance to abiotic stresses: a review[J]. Cells,2021,10(6): 1551
[8]SONG Y, LI X, YAO S, et al. Correlations between soil metabolomics and bacterial community structures in the pepper rhizosphere under plastic greenhouse cultivation[J]. Science of the Total Environment, 2020, 728: 138439.
[9]KANG S, KHAN A L, WAQAS M, et al. Plant growth-promoting rhizobacteria reduce adverse effects of salinity and osmotic stress by regulating phytohormones and antioxidants in Cucumis sativus[J]. Journal of Plant Interactions, 2014, 9(1): 673-682.
[10]YUAN J, ZHAO J, WEN T, et al. Root exudates drive the soil-borne legacy of aboveground pathogen infection[J]. Biomed Central, 2018, 6(1): 156.
[11]BAL H, NAYAK L,DAS S, et al. Isolation of ACC deaminase producing PGPR from rice rhizosphere and evaluating their plant growth promoting activity under salt stress[J]. Plant Soil,2013, 366: 93-105.
[12]KIM Y C, LEVEAU J, MCSPADDEN GARDENER B B, et al. The multifactorial basis for plant health promotion by plant-associated bacteria[J]. Applied and Environmental Microbiology, 2011, 77(5): 1548-1555.
[13]KASIM W A, GAAFAR R M, ABOU-ALI R M, et al. Effect of biofilm forming plant growth promoting rhizobacteria on salinity tolerance in barley[J]. Annals of Agricultural Sciences, 2016,61(2):217-227.
[14]楊瀟湘,張蕾,黃小琴,等. 基于高通量測序分析大豆和油菜根際微生物群落結構的差異[J]. 應用生態學報, 2019, 30(7): 2345-2351.
[15]劉振香,劉鵬,賈緒存,等. 不同水肥處理對夏玉米田土壤微生物特性的影響[J]. 應用生態學報, 2015, 26(1): 113-121.
[16]姜莉莉,宮慶濤,武海斌,等. 不同生草處理對蘋果園土壤微生物群落的影響[J]. 應用生態學報, 2019, 30(10): 3482-3490.
[17]艾鑠,張麗杰,肖芃穎,等. 高通量測序技術在環境微生物領域的應用與進展[J]. 重慶理工大學學報(自然科學), 2018, 32(9): 111-121.
[18]王林,李冰,朱健. 高通量測序技術在人工濕地微生物多樣性研究中的研究進展[J]. 中國農學通報, 2016, 32(5): 10-15.
[19]牛世全,龍洋,李海云,等. 應用IlluminaMiSeq高通量測序技術分析河西走廊地區鹽堿土壤微生物多樣性[J]. 微生物學通報, 2017, 44(9): 2067-2078.
[20]尹原森,馬國勝,曹春燕,等. 不同地區鳳丹根際土壤微生物功能多樣性分析[J]. 分子植物育種, 2021, 19(20):6918-6926.
[21]郭麗麗,尹偉倫,郭大龍,等. 油用鳳丹牡丹不同種植時間根際細菌群落多樣性變化[J]. 林業科學, 2017, 53(11): 131-141.
[22]李昱瑩,劉曙光,廉小芳,等. 油用牡丹鳳丹不同種植年限根際真菌群落多樣性變化研究[J]. 基因組學與應用生物學, 2020, 39(4): 1672-1685.
[23]全國花卉標準化技術委員會. 油用牡丹栽培技術規程: LY/T 2958-2018[S]. 北京: 國家林業局,2018.
[24]章家恩,高愛霞,徐華勤,等. 玉米/花生間作對土壤微生物和土壤養分狀況的影響[J]. 應用生態學報, 2009, 20(7): 1597-1602.
[25]王悅,楊貝貝,王浩,等. 不同種植模式下丹參根際土壤微生物群落結構變化[J]. 生態學報, 2019, 39(13): 4832-4843.
[26]王麗霞,井濤,殷曉敏,等. 不同種植模式對香蕉根區土壤養分及可培養微生物數量的影響[J]. 中國南方果樹, 2020, 49(4): 80-86.
[27]陳澤斌,陳敏,阮亞男,等. 不同種植模式草莓根際細菌和真菌群落結構的變化[J]. 昆明學院學報, 2019, 41(6): 52-56.
[28]李娟. 油用牡丹與經濟林木立體栽培模式研究[D]. 鄭州: 鄭州大學, 2019.
[29]王國霞,羅青,楊玉珍,等. 立體栽培模式對油用牡丹生長和產量的影響[J]. 貴州農業科學, 2018, 46(12): 100-103.
[30]鮮文東,張瀟橦,李文均. 綠彎菌的研究現狀及展望[J]. 微生物學報, 2020,60(9):1801-1820.
[31]劉芯竹. 覆蓋經營對毛竹林土壤理化性質和細菌群落多樣性影響[D]. 杭州:浙江農林大學, 2021.
[32]張永敢,趙娟,張玉潔,等. 藥用植物鳳丹(Paeonia suffruticosa)根際土壤細菌群落16S rRNA基因的ARDRA分析[J]. 生態學報, 2016, 36(17): 5564-5574.
[33]南麗麗,郭全恩,譚杰輝,等. 輪作休耕模式對土壤細菌群落的影響[J]. 干旱地區農業研究, 2020, 38(6): 128-134.
[34]李倩,袁玲,楊水平,等. 連作對黃花蒿生長及土壤細菌群落結構的影響[J]. 中國中藥雜志, 2016, 41(10): 1803-1810.
[35]楊安娜,陸云峰,張俊紅,等. 杉木人工林土壤養分及酸桿菌群落結構變化[J]. 林業科學, 2019, 55(1): 119-127.
[36]劉婷,肖仲久,李小霞,等. 高通量測序技術分析茅臺釀酒高粱根際真菌群落特征[J]. 內蒙古農業大學學報(自然科學版), 2021,43(1): 48-52.
[37]馬繼瓊,孫一丁,楊奕,等.不同葉稻瘟抗性水稻品種對根際真菌的影響[J]. 江蘇農業科學,2021,49(12):75-80.
[38]劉震,徐玉鵬,王秀領,等.黑龍港苜蓿根際土壤真菌群落結構及多樣性分析[J]. 江蘇農業科學,2021,49(10):197-201.
[39]薛曉敏,王來平,韓雪平,等. 不同樹盤覆蓋對矮砧蘋果園土壤微生物群落結構和多樣性的影響[J]. 生態學報, 2021,41(4): 1528-1536.
[40]王麗娟,王威威,呂雪,等. 鹿蹄草內生真菌的分離鑒定及其對鹿蹄草的生理效應[J]. 東北農業大學學報, 2013, 44(8): 87-93.
[41]MICHAL J J, ANATOLI L, ALEXANDRA F, et al. The beneficial root-colonizing fungus Mortierella hyalina promotes the aerial growth of Arabidopsis and activates calcium-dependent responses which restrict Alternaria brassicae-induced disease development in roots.[J]. Molecular Plant-microbe Interactions, MPMI, 2018, 32(3): 351-363.
[42]張向民. 鐮刀菌屬分類學研究歷史與現狀[J]. 菌物研究, 2005(2): 63-66.
[43]唐濤,王帆帆,郭杰,等. 12種生物源殺菌劑對白術根腐病的防效[J]. 植物保護, 2021, 47(3): 288-293.
[44]耿貴,楊瑞瑞,於麗華,等. 作物連作障礙研究進展[J]. 中國農學通報, 2019, 35(10): 36-42.
[45]楊睿. 貴州喀斯特山區花椒林根際土壤微生物群落結構及功能研究[D]. 貴州:貴州師范大學, 2021.
[46]楊睿,李娟,龍健,等. 貴州喀斯特山區不同種植年限花椒根際土壤細菌群落結構特征研究[J]. 生態環境學報, 2021, 30(1): 81-91.
[47]趙鵬宇,白雪,燕平梅,等. 華北落葉松林土壤細菌群落結構與表型的環境異質性響應[J]. 林業科學, 2021, 57(7): 101-110.
(責任編輯:陳海霞)
收稿日期:2021-12-16
基金項目:江蘇省農業科技自主創新基金項目[CX(19)3066] ;江蘇現代農業產業技術體系建設項目[JATS(2021)507]
作者簡介:劉智(1996-),男,陜西漢中人,碩士研究生,主要從事油用牡丹根際微生物研究。(E-mail)liuzhi2021702@163.com
通訊作者:王仲偉,(E-mail)w0414115@Sina.cn;湯詩杰,(E-mail)tangshijie69@aliyun.com
