雞爪槭金陵丹楓和金陵黃楓葉片呈色分析
2022-05-16 23:40:31朱璐聞婧馬秋月顏坤元杜一鳴李淑順李倩中
江蘇農業學報 2022年2期
朱璐 聞婧 馬秋月 顏坤元 杜一鳴 李淑順 李倩中


摘要:以江蘇省農業科學院自主選育的雞爪槭新品種金陵丹楓和金陵黃楓為研究對象,對其4月至7月觀賞期葉片進行葉色對比觀察,發現金陵丹楓新葉為亮紅色,而后紅色逐漸變淡,而金陵黃楓新葉為黃色,邊緣呈淺珊瑚色,而后逐漸變為金黃色。比對金陵丹楓和金陵黃楓葉片中的葉綠素、類胡蘿卜素、花青素和黃酮類物質含量,發現金陵丹楓亮紅色葉片中各種色素含量均高于同時期的金陵黃楓金黃色葉片,且花青素和黃酮生物合成途徑中一系列關鍵基因(ApPAL、ApCHS、ApCHI、ApF3H、ApF3′H、ApDFR和ApANS)的相對表達量在金陵丹楓葉片中上調。研究結果提示,這些色素含量的不同可能是導致金陵丹楓和金陵黃楓葉片呈色不同的主要原因。尤其是3種花青素(飛燕草素、矢車菊素和矢車菊半乳糖苷)在金陵丹楓亮紅色葉片中的含量遠高于金陵黃楓金黃色葉片,表明花青素在金陵丹楓葉片呈色中起到關鍵作用。
關鍵詞:雞爪槭;葉片;色素
中圖分類號:S687文獻標識碼:A文章編號:1000-4440(2022)02-0521-07
Analysis on leaf colour generation of Acer palmatum Jinling Danfeng and Jinling Huangfeng
ZHU Lu WEN Jing,MA Qiu-yue,YAN Kun-yuan,DU Yi-ming,LI Shu-shun,LI Qian-zhong
Abstract:New varieties of Acer palmatum Thunb. named Jinling Danfeng and Jinling Huangfeng breeded by Jiangsu Academy of Agricultural Sciences independently were used as experimental materials and colours of the leaves during ornamental period from April to July were observed and compared. It was found that the new leaves of Jinling Danfeng were bright red and then the colour gradually faded, while the new leaves of Jinling Huangfeng were yellow with light coral on the edges, which gradually turned into golden yellow. After comparing the contents of chlorophyll, carotenoids, anthocyanins and flavonoids in the leaves of Jinling Danfeng and Jinling Huangfeng, it was found that contents of various pigments in the bright red leaves of Jinling Danfeng were higher than that in golden yellow leaves of Jinling Huangfeng at the same period, and the relative transcript levels of a series of key genes (ApPAL, ApCHS, ApCHI, ApF3H, ApF3′H, ApDFR and ApANS) in anthocyanin and flavonoid biosynthetic pathways in the leaves of Jinling Danfeng were up-regulated. The results indicated that, differences in pigments content may be the main reason for the different colours generation in Jinling Danfeng and Jinling Huangfeng leaves. In particular, the contents of three anthocyanins (delphinidin, cyanidin and cyanidin 3-galactoside) in the bright red leaves of Jinling Danfeng were much higher than those in the golden yellow leaves of Jinling Huangfeng, indicating that the anthocyanins played a key role in the colour generation in the leaves of Jinling Danfeng.
Key words:Acer palmatum;leaf;pigment
雞爪槭(Acer palmatum Thunb.)為槭樹科槭屬落葉小喬木,因其葉片豐富的季相變化和優美的形態,成為中國極為重要的觀賞彩葉樹種,并被廣泛應用于園林景觀和城市綠化中[1-2]。隨著季節更替,不斷變化的葉色是雞爪槭重要的觀賞性狀之一。影響植物葉色呈現的因素非常復雜,其中色素(主要包括葉綠素、類胡蘿卜素和花青素等)的組成、含量及配比是最為直接的影響因素[3-5]。
葉綠素(Chlorophyll)不僅是植物進行光合作用的重要物質基礎,也是葉片呈現綠色至關重要的原因。葉綠素代謝途徑主要包含葉綠素合成、循環和降解[6-7]。其中任何一步受到阻礙都有可能導致葉片失綠,造成黃葉甚至是白化葉片的出現。如水稻嘌呤合成途徑中的關鍵酶VAL1,通過參與調控葉片中葉綠體發育和葉綠素代謝過程影響葉綠素的合成,該基因突變后導致水稻葉色白化[8]。水稻RA1基因編碼甘氨酰-tRNA合成酶(GlyRS),該基因通過參與水稻早期葉綠體的發育影響葉綠素的合成,其突變體ra1在幼苗期呈現白化葉片[9]。類胡蘿卜素(Carotenoids)含量的變化是影響葉色呈現的另一重要因素。目前植物中類胡蘿卜素生物合成途徑已較為清晰,八氫番茄紅素合成酶(PSY)是類胡蘿卜素生物合成途徑的第一個限速酶[10-11]。在銀杏黃葉突變體中,類胡蘿卜素生物合成途徑中的關鍵基因PSY、Z-ISO、ZDS和LCYE等的表達量均上調[12]。黃酮類化合物(Flavonoids)可以產生豐富的色彩,是彩葉植物呈色的主要影響因素。花青素(Anthocyanin)是植物類黃酮化合物中重要的一類,主要分為6大類:天竺葵素(Pelargonidin)、矢車菊素(Cyanidin)、芍藥花素(Peonidin)、飛燕草素(Delphinidin)、矮牽牛素(Petunidin)和錦葵素(Malyidin)。呈現不同色彩的花色苷大多由這6類花青素衍生而來[13-14]。目前花青素生物合成途徑作為苯丙烷類代謝途徑的一個重要分支已被完整解析。PAL、C4H、CHS、CHI、F3H、F3′H、DFR和ANS基因均為花青素合成途徑中的關鍵基因,研究者已在多種植物,如海棠、康乃馨、葡萄、小蒼蘭和煙草中發現這些關鍵基因表達差異影響了花青素的含量[15-19]。
本研究以江蘇省農業科學院自主選育的雞爪槭新品種金陵丹楓和金陵黃楓為試驗材料,通過分析這2個品種4月至7月葉片中葉綠素、類胡蘿卜素、花青素和黃酮的含量及花青素和黃酮生物合成途徑中一些關鍵基因的表達量,初步解析雞爪槭金陵丹楓和金陵黃楓葉片呈色機制,以期為品種改良提供理論依據。
1材料與方法
1.1試驗材料
供試雞爪槭品種金陵丹楓和金陵黃楓種植于江蘇省農業科學院槭樹良種基地。于2020年4月8日開始取樣,每隔15 d取第3張和第4張完全展開葉用于拍照、色素提取和基因表達量分析。取樣重復3次,取樣后將樣品立即放入液氮中,于-80 ℃冰箱保存備用。
1.2葉綠素和類胡蘿卜素提取及測量
取金陵丹楓和金陵黃楓7個不同時期的新鮮葉片,分別稱量0.1 g,剪碎放入10 ml丙酮-0.1 mol/L NH4OH提取液(丙酮、NH4OH體積比為9∶1)中浸提24 h。使用全波長酶標儀(MD SpectraMax 190)分別在663 nm、645 nm和440 nm下測量提取液的吸光度,以空白提取液作為對照。根據以下公式分別計算葉綠素a、葉綠素b和類胡蘿卜素的含量。計算公式如下:Ca=12.72A663-2.59A645,Cb=22.88A645-4.67A663,Cc=4.695A440-0.268(Ca+Cb) ,式中,Ca為葉綠素a含量,Cb為葉綠素b含量,Cc為類胡蘿卜素含量,A為吸光度。
1.3花青素和黃酮提取及測量
取金陵丹楓和金陵黃楓7個不同時期葉片,分別稱量0.2 g,用液氮研磨后將粉末轉移至10 ml提取液[甲醇∶蒸餾水∶甲酸∶三氟乙酸=70∶27∶2∶1(體積比)]中,超聲波30 min,4 ℃黑暗浸提24 h,其間每6 h振蕩1次。使用高效液相色譜儀(Agilent 1290)分離黃酮類化合物,檢測參數為C18色譜柱(Waters, 1.8 μm, 2.1 mm×75.0 mm),進樣體積1 μl。使用A(0.1%甲酸)和B(色譜甲醇)作為流動相,梯度洗脫條件為0~7 min,90%~10% A,10%~90% B;7~10 min,10%~90% A,90%~10% B,流速0.3 ml/min。再使用三重四級桿液質聯用系統(SCIEX QTRAP 6500+ LC-MS/MS)進行檢測及定量,根據標準品建立的標準曲線計算葉片中花青素和黃酮含量。
1.4花青素和黃酮合成途徑關鍵基因表達分析
取金陵丹楓和金陵黃楓7個不同時期葉片,利用Trizol(TaKaRa)提取法提取葉片總RNA,并使用RNase-free DNase I(TaKaRa)去除基因組DNA。利用反轉錄酶M-MLV(TaKaRa)將RNA反轉錄獲得cNDA備用。利用熒光定量PCR(Real-time PCR)技術對花青素和黃酮合成途徑中的關鍵基因進行表達分析,使用ApActin基因(MN026864)[20]作為內參基因。反應體系為:10 μl SYBR Premix Ex TaqTM(TaKaRa),各0.4 μl上游和下游引物(10 μmol/L),5 μl cDNA模板(1 ng/μl),0.4 μl Reference Dye Ⅱ(ROX),用ddH2O補至20 μl。使用Real-Time熒光定量PCR儀(Applied Biosystems 7500, Carlsbad, USA)進行相對實時熒光定量PCR。PCR反應程序:95 ℃ 30 s;95 ℃ 5 s,60 ℃ 35 s,40個循環;95 ℃ 15 s,60 ℃ 60 s,95 ℃,15 s,60 ℃ 15 s[20]。采用2-△△Ct法計算各個基因的相對表達量,每個反應設3個生物學重復。利用Primer 5.0軟件設計花青素和黃酮合成途徑中關鍵基因的特異性引物,引物序列見表1。
2結果與分析
2.1金陵丹楓和金陵黃楓4月至7月葉片表型分析
于2020年4月8日開始每隔15 d對金陵丹楓和金陵黃楓進行采樣觀察,發現金陵丹楓在4月8日至23日葉片呈亮紅色,而后亮紅色逐漸變淺,直至7月7日逐漸成為淡橙黃色。而金陵黃楓4月8日至23日新葉呈黃色,葉緣呈淺珊瑚色,而后葉片逐漸成為金黃色,6月22日至7月7日葉片開始轉綠(圖1)。
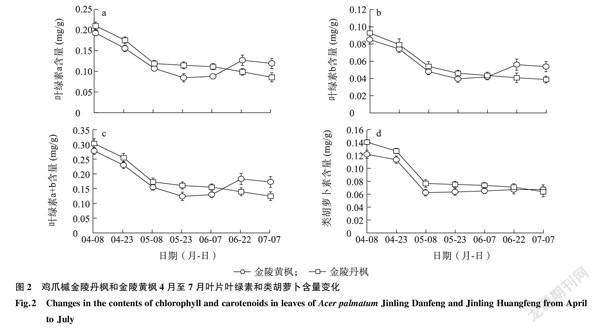
2.2金陵丹楓和金陵黃楓葉片葉綠素和類胡蘿卜素含量分析
分析金陵丹楓和金陵黃楓7個時期葉片中的葉綠素和類胡蘿卜素含量,發現金陵丹楓在4月8日至6月7日葉綠素a、葉綠素b和總葉綠素含量均高于金陵黃楓,但隨著金陵黃楓葉片自6月22日開始轉綠,其葉綠素含量開始高于金陵丹楓葉片(圖2a~圖2c)。金陵丹楓葉片中的葉綠素a、葉綠素b、總葉綠素和類胡蘿卜含量則一直呈現下降趨勢(圖2a~圖2d)。金陵黃楓金黃色成熟葉片(5月8日至7月7日)中的類胡蘿卜素含量也低于具有淺珊瑚色葉緣的金陵丹楓葉片(4月8日至4月23日)(圖2d)。
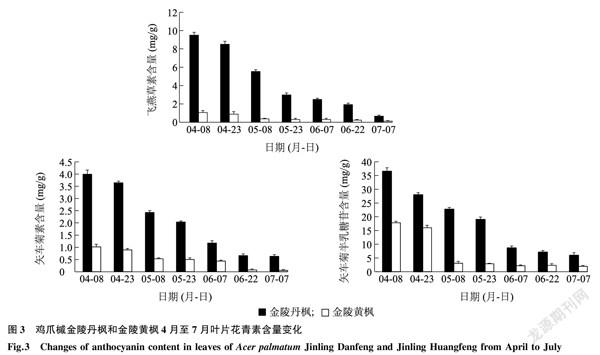
2.3金陵丹楓和金陵黃楓葉片黃酮類物質含量分析
采用超高效液質聯用法對金陵丹楓和金陵黃楓7個時期葉片中的飛燕草素、矢車菊素和矢車菊半乳糖苷含量進行檢測,發現金陵丹楓7個時期葉片中飛燕草素、矢車菊素和矢車菊半乳糖苷含量均遠高于金陵黃楓。其中飛燕草素在5月23日的金陵丹楓葉片中含量是金陵黃楓的9.9倍,矢車菊素在7月7日的金陵丹楓葉片中含量是金陵黃楓的11.5倍,矢車菊半乳糖苷在5月8日的金陵丹楓葉片中含量是金陵黃楓的7.3倍。隨著金陵丹楓葉片紅色逐漸褪去,金陵黃楓葉片完全變成金黃色,這3種花青素的含量逐漸降低(圖3)。在金陵黃楓葉片轉成綠時,這3種花青素含量均降至最低。
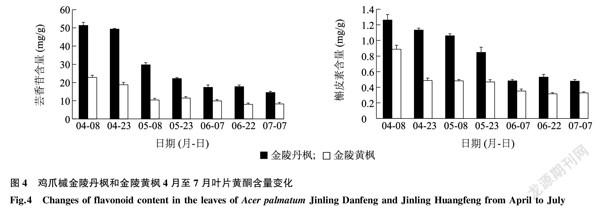
采用超高效液質聯用法對金陵丹楓和金陵黃楓7個時期葉片中的蕓香苷和槲皮素含量進行檢測。結果顯示,金陵丹楓7個時期葉片中的蕓香苷和槲皮素含量均高于金陵黃楓。其中蕓香苷在5月8日的金陵丹楓葉片中含量是金陵黃楓的2.9倍,而槲皮素在4月23日的金陵丹楓葉片中含量是金陵黃楓的2.3倍。蕓香苷和槲皮素在金陵丹楓和金陵黃楓葉片中的含量均在4月至7月間呈逐漸降低的趨勢(圖4)。
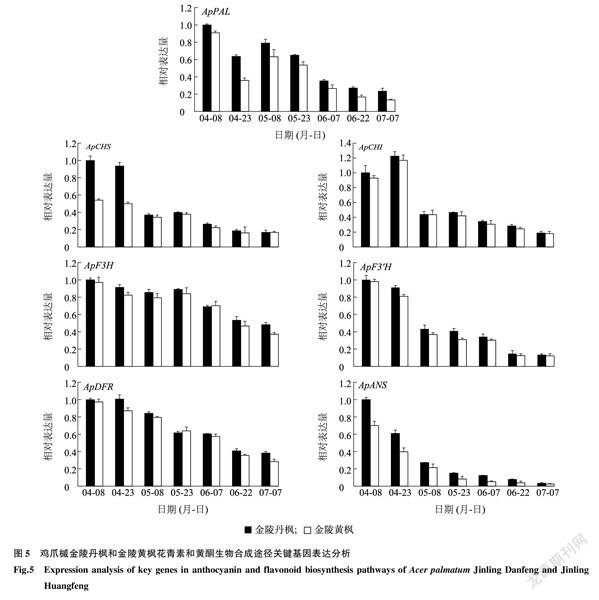
2.4花青素和黃酮生物合成途徑關鍵基因表達分析
利用熒光定量PCR技術對金陵丹楓和金陵黃楓7個時期葉片中花青素和黃酮生物合成途徑關鍵基因進行表達分析,發現花青素和黃酮生物合成途徑關鍵基因ApPAL、ApCHS、ApCHI、ApF3H、ApF3′H、ApDFR和ApANS在金陵丹楓中的相對表達量均高于金陵黃楓。4月至7月間,金陵丹楓葉片亮紅色逐漸變淡,金陵黃楓葉片完全變成金黃色,這些關鍵基因的相對表達量也大致呈現逐漸降低的趨勢(圖5)。
3討論
葉色是雞爪槭重要的觀賞性狀之一。葉片呈色表達是內外環境因子(如光照、溫度、植物激素和糖類等)和植物自身遺傳因子共同作用的結果[21-22]。而色素的種類、含量及其分布均是直接影響葉色呈現的重要因素。葉綠素、類胡蘿卜素和黃酮類化合物是影響植物呈色的重要色素,它們共同作用,使雞爪槭葉片呈現多種顏色模式[22-23]。金陵丹楓和金陵黃楓均是江蘇省農業科學院自主選育的雞爪槭新品種。金陵丹楓是從金陵黃楓組培苗中選育出的新品種[24-25],兩者的遺傳背景極為相似。金陵丹楓的亮紅色葉片和金陵黃楓的金黃色葉片為研究雞爪槭春季彩葉呈現提供了試驗材料。
葉綠素是植物進行光合作用的重要物質,也是植物呈現綠色的主要因素[26]。彩色葉片中的葉綠素含量往往低于綠色葉片。如雞爪槭黃葉突變體金陵黃楓中的葉綠素含量遠低于普通雞爪槭[1,23];銀杏和美洲黑楊的黃葉突變體中的葉綠素含量也比正常植株明顯降低,且葉綠素合成途徑相關基因表達量下調,葉綠素降解途徑相關基因表達量上調[12,22]。本研究發現,金陵黃楓的突變株金陵丹楓亮紅色葉片的葉綠素含量高于金陵黃楓的金黃色葉片,提示金陵丹楓的高葉綠素含量可能也在其紅色呈現中起著一定作用。
類胡蘿卜素不僅是一類光合和保護色素,其含量的提高也往往導致葉片呈現黃色[27]。比如,櫟樹和銀杏黃葉突變體中類胡蘿卜素含量升高,類胡蘿卜素合成途徑相關基因表達量上調[12,28];大花蕙蘭花葉突變體中類胡蘿卜素含量顯著降低,轉錄組分析發現多個類胡蘿卜素生物合成相關基因表達量下調[29]。本研究結果卻顯示,6月22日前的金陵丹楓紅色葉片中的類胡蘿卜素含量也高于金陵黃楓,這提示類胡蘿卜素確實在金陵丹楓葉片紅色呈現中起著一定作用。
花青素以及其他黃酮類物質含量的變化是影響植物器官呈色的決定性因素之一[30-32]。紫薯較紅薯塊根中積累了大量的花青素,其花青素代謝途徑中的一系列基因被上調表達[33]。文心蘭花瓣中同時具有紅色和黃色2種顏色,其中紅色花瓣中含有錦葵色素半乳糖、芍藥素葡萄糖苷、飛燕草素葡萄糖苷和矢車菊素葡萄糖苷這4種花色苷,而黃色花瓣中則未檢測到這4種花色苷,進一步研究發現花青素代謝途徑中的2個關鍵基因CHI和DRF在黃色花瓣中表達量下調是導致兩種顏色形成的主要原因[34]。本研究發現,金陵丹楓葉片中的花青素(飛燕草素、矢車菊素和矢車菊半乳糖苷)和蕓香苷、槲皮素等黃酮類物質含量遠高于金陵黃楓,同時其花青素等黃酮類物質生物合成關鍵酶的編碼基因(ApPAL、ApCHS、ApCHI、ApF3H、ApF3′H、ApDFR和ApANS)表達量上調。這提示花青素等黃酮類物質含量的不同是導致金陵丹楓和金陵黃楓葉片呈現不同顏色的重要原因。花青素和黃酮含量除了受上述生物合成途徑基因的調控外,轉錄水平的調控也同樣起作用,已知影響花青素和黃酮的轉錄因子主要有MYB、bHLH、WD40 3類[35-37]。金陵丹楓和金陵黃楓葉片呈色不同是否受到上述轉錄因子的調控還需進一步研究。
參考文獻:
[1]LI S S, LI Q Z, Rong L P, et al. Gene expressing and sRNA sequencing show that gene differentiation associates with a yellow Acer palmatum mutant leaf in different light conditions[J]. Biomed Research International, 2015, 2015:1-10.
[2]KIM J H, LEE B C, SIM G S, et al. The isolation and antioxidative effects of vitexin from Acer palmatum [J]. Archives of Pharmacal Research, 2005,28:195.
[3]徐明遠,何鵬,賴偉,等. 植物葉色變異分子機制研究進展[J].分子植物育種, 2021, 19(10):3448-3455.
[4]TANAKA Y, SASAKI N, OHMIYA A. Biosynthesis of plant pigments: anthocyanins, betalains and carotenoids[J]. The Plant Journal, 2008,54:733-749.
[5]ZHAO D Q, TAO J. Recent advances on the development and regulation of flower color in ornamental plants[J]. Frontiers in Plant Science, 2015,6:261.
[6]KRAUTLER B. Breakdown of chlorophyll in higher plants-phyllobilins as abundant, yet hardly visible signs of ripening, senescence, and cell death[J]. Angewandte Chemim-International Edition, 2016, 55(16):4882-4907.
[7]SCHELBERT S, AUBYR S, BURTA B, et al. Pheophytin pheophorbide hydrolase (pheophytinase) is involved in chlorophyll breakdown during leaf senescence in Arabidopsis[J]. Plant Cell, 2009, 21(3):767-785.
[8]ZHANG T, FENG P, LI Y F, et al. VIRESCENT-ALBINO LEAF 1 regulates leaf colour development and cell division in rice[J]. Journal of Experimental Botany, 2018, 69(20): 4791-4804.
[9]ZHENG H,WANG Z R, TIAN Y L, et al. Rice albino 1, encoding a glycyl-tRNA synthetase, is involved in chloroplast development and establishment of the plastidic ribosome system in rice[J]. Plant Physiology and Biochemistry, 2019, 139:495-503.
[10]CAZZONELLI C I, POGSON B J. Source to sink: regulation of carotenoid biosynthesis in plants[J]. Trends in Plant Science, 2010, 15(5): 266-274.
[11]GADY A L F, VRIEZEN W H, VAN DE WAL M H B J, et al. Induced point mutations in the phytoene synthase 1 gene cause differences in carotenoid content during tomato fruit ripening[J]. Molecular Breeding, 2012, 29(3): 801.
[12]LI W X, YANG S B, LU Z G, et al. Cytological, physiological, and transcriptomic analyses of golden leaf coloration in Ginkgo biloba L[J]. Horticulture Research, 2018, 5:12.
[13]劉愷媛,王茂良,辛海波,等. 植物花青素合成與調控研究進展[J]. 中國農學通報, 2021, 37(14):41-51.
[14]SMERIGLIO A, BARRECA D, BELLOCCO E. et al. Chemistry, pharmacology and health benefits of anthocyanins[J]. Phytotherapy Research, 2016, 30(8):1265-1286.
[15]ITOH Y, HIGETA D, SUZUKI A, et al. Excision of transposable elements from the chalcone isomerase and dihydroflavonol 4-reductase genes may contribute to the variegation of the yellow-flowered carnation (Dianthus caryophyllus)[J]. Plant and Cell Physiology, 2002, 43(5):578-585.
[16]NISHIHARA M, NAKATSUKA T, YAMAMURA S. Flavonoid components and flower color change in transgenic tobacco plants by suppression of chalcone isomerase gene[J]. FEBS Letters, 2005, 579(27): 579.
[17]SUN W, MENG X Y, LIANG L J, et al. Molecular and biochemical analysis of chalcone synthase from freesia hybrid in flavonoid biosynthetic pathway[J]. Plos One, 2015, 10(3):e0119054.
[18]TAI D Q, TIAN J, ZHENG J, et al. A malus crabapple chalcone synthase gene, mcchs, regulates red petal color and flavonoid biosynthesis[J]. Plos One, 2014, 9(10): e110570.
[19]WANG H L, WANG W, LI H, et al. Expression and tissue and subcellular localization of anthocyanidin synthase (ANS) in grapevine[J]. Protoplasma, 2011, 248(2): 267.
[20]ZHU L, MA Q Y, LI S S, et al. Reference gene selection for quantitative real-time PCR analyses of Acer palmatum under abiotic stress[J]. Phyton-International Journal of Experimental Botany, 2020, 89(2):385-403.
[21]TANAKA Y, BRUGLIERA F, CHANDLER S. Recent progress of flower colour modification by biotechnology[J]. International Journal of Molecular Sciences, 2009, 10(12):5350-5369.
[22]CHEN Z, LU X Y, XUAN Y, et al. Transcriptome analysis based on a combination of sequencing platforms provides insights into leaf pigmentation in Acer rubrum[J]. BMC Plant Biology, 2019, 19(1):240.
[23]LI S S, LI Q Z, T L, et al. Pigment comparison and expression of chlorophyll metabolism genes in yellow and green Acer palmatum leaves[J]. Canadian Journal of Plant Scienc, 2017, 97(5), 1-21.
[24]李倩中,李淑順,榮立蘋,等. 雞爪槭新品種‘金陵黃楓’[J]. 園藝學報, 2011, 38(8):1627-1628.
[25]聞婧,李淑順,朱璐,等. 雞爪槭新品種‘金陵丹楓’[J]. 園藝學報, 2019,46(S2):2920-2921.
[26]MASUDA T, FUJITA Y. Regulation and evolution of chlorophyll metabolism[J]. Photochemical & Photobiological Sciences Official Journal of the European Photochemistry Association & the European Society for Photobiology, 2008, 7(10):1131-1149.
[27]DEMMING A B, ADAMS W W. The role of xanthophyll cycle carotenoids in the protection of photosynthesis[J]. Trends in Plant Science, 1996, 1(1): 21-26.
[28]DONG X Y, HUANG L B, CHEN Q S, et al. Physiological and anatomical differences and differentially expressed genes reveal yellow leaf coloration in Shumard oak[J]. Plants, 2020, 9(2):169.
[29]JIANG Y, SONG H Y, HE J R, et al. Comparative transcriptome analysis provides global insight into gene expression differences between two orchid cultivars[J]. PLoS One, 2018,13(7):e0200155.
[30]JUN J H, XIAO X, RAO X, et al. Proanthocyanidin subunit composition determined by functionally diverged dioxygenases[J]. Nature Plants, 2018, 4(12):1034-1043.
[31]GUO J, HAN W, WANG M H. Ultraviolet and environmental stresses involved in the induction and regulation of anthocyanin biosynthesis: a review[J]. African Journal of Biotechnology, 2008, 25:1-4.
[32]練冬梅,賴正鋒,姚運法,等.玫瑰茄轉錄組測序及花青素合成相關基因表達分析[J].江蘇農業科學,2020,48(6):41-45.
[33]MANO H, OGASAWARA F, SATO K, et al. Isolation of a regulatory gene of anthocyanin biosynthesis in tuberous roots of purple-fleshed sweet potato[J]. Plant Physiology, 2007, 143(3):1252-1268.
[34]CHIOU C Y, YEH K W. Differential expression of MYB gene (OgMYB1) determines color patterning in floral tissue of Oncidium Gower Ramsey[J]. Plant Molecular Biology, 2008, 66(4):379-388.
[35]DAVIES K M, ALBERT N W, SCHWINN K E. From landing lights to mimicry: the molecular regulation of flower colouration and mechanisms for pigmentation patterning[J]. Functional Plant Biology, 2012, 39:619-638.
[36]RAMSAY N A, GLOVER B J. MYB-b HLH-WD40 protein complex and the evolution of cellular diversity[J]. Trends in Plant Science, 2005, 10:63-70.
[37]ZHU Z X, WANG H L, WANG Y Y, et al. Characterization of the cis elements in the proximal promoter regions of the anthocyanin pathway genes reveals a common regulatory logic that governs pathway regulation[J]. Journal of Experimental Botany, 2015, 66:3775-3789.
(責任編輯:張震林)
收稿日期:2021-08-19
基金項目:國家自然科學基金項目(32001357);江蘇省農業科技自主創新資金項目[CX(18)3073]
作者簡介:朱璐(1987-),女,山東淄博人,博士,副研究員,研究方向為雞爪槭遺傳育種。(E-mail)luzhu@jaas.ac.cn
通訊作者:李倩中,(E-mail)qianzhongli@jaas.ac.cn;李淑順,(E-mail)shushunli@jaas.ac.cn
