‘華仲6號’杜仲嫩枝扦插生根的解剖及生理變化*
2022-05-11 11:54:08呂庚鑫孟益德劉攀峰杜慶鑫杜紅巖杜蘭英
林業科學 2022年2期
呂庚鑫 孟益德 慶 軍 何 鳳 劉攀峰 杜慶鑫 杜紅巖 杜蘭英 王 璐
(1.中國林業科學研究院經濟林研究開發中心 鄭州 450003; 2.南京林業大學 南京 210037; 3.國家林業和草原局泡桐研究開發中心 經濟林種質創新與利用國家林業和草原局重點實驗室 鄭州 450003)
杜仲(Eucommiaulmoides)是我國特有的國家戰略儲備資源,在我國29個省(區、市)均有分布(劉攀峰等, 2020)。杜仲高值化綜合利用涉及到杜仲樹皮、雄花、果實、葉片、木材等部位(杜慶鑫等, 2017),在航空航天、化工、醫療等多個領域具有巨大的開發潛能及應用前景(杜紅巖等, 2020)。《全國杜仲產業發展規劃(2016—2030)》明確規劃,到2030年全國新造杜仲林面積333.35萬hm2,發展潛力巨大。現有杜仲資源中約90%為實生林,良種資源規模小且普及率低,苗木供不應求,嚴重限制杜仲產業發展。
杜仲為雌雄異株,種子繁殖不能保持親本的優良性狀,因此杜仲良種繁育需采用無性繁殖的技術。杜仲良種繁育目前主要以嫁接為主(杜紅巖等, 2020),但其成本較高、受季節限制。杜仲組培苗愈傷組織生根困難,褐化、玻璃化等現象嚴重,組培體系的建立未取得技術突破(李響等, 2019; 劉閔豪等, 2020)。楸樹(Catalpabungei)、板栗(Castaneamollissima)、桑樹(Morusalba)等樹種都已通過扦插技術實現了良種的快速繁育(王新建等, 2009; 杜常健等, 2019; 杜偉, 2016)。扦插不僅能保持母本優良性狀,而且操作簡單、成本低、效率高,對于杜仲良種繁育具有重要意義(康向陽, 2017)。
許多研究證實: 扦插成功的關鍵在于不定根的形成和數量,而生長素被認為是誘導根原基發生的先決條件(Duetal., 2016; Druegeetal., 2016),氧化酶和營養物質對不同樹種扦插生根也存在促進或抑制作用(歐陽芳群等, 2015)。近年來杜仲良種嫩枝扦插技術取得了較大突破,顯著提高了杜仲良種嫩枝扦插的生根率(王高鵬等, 2018),但對于杜仲良種嫩枝扦插生根的相關機理尚未開展系統研究。鑒于此,本文以‘華仲6號’杜仲半木質化春梢為試材,對插穗基部進行生根解剖結構和形態觀察,測定生根過程中內源激素、營養物質含量及氧化酶活性等相關生理指標,從解剖和生理學探討杜仲良種嫩枝扦插不定根發生機制。
1 材料與方法
1.1 插穗制備與扦插
試驗材料取自中國林業科學研究院經濟林研究開發中心的河南孟州試驗基地(112°33′E,34°50′N),以健壯、長勢良好、無病蟲害的國審良種‘華仲6號’杜仲3年生嫁接苗為母樹,取上部當年生半木質化枝條制備插穗。
扦插池長8 m、寬1.5 m,下部鋪15 cm厚的粗河砂、上部鋪15 cm厚的基質。以草炭土∶珍珠巖(體積比3∶1)作扦插基質,扦插前用40%多菌靈可濕性粉劑500倍消毒。2020年4月28日早晨采集并制備插穗,插穗長度控制在10~15 cm,保留2~3個飽滿芽和葉片。經杜仲專用生根劑快速浸蘸10 s的插穗作為處理組,清水處理作為對照組,每處理各扦插一池。扦插后每周用多菌靈消毒1次,依據天氣情況合理調整噴霧間隔及時間,控制棚內溫度20~29 ℃,空氣相對濕度90%以上。
1.2 樣品采集
外部形態觀察的取樣從扦插當天開始,每2天取樣1次,直到有不定根生成。每次隨機取20根,挑選生長狀態一致的3~ 4根。取插穗基部1~3 cm長的莖段,放入裝有70% FAA固定液中,實驗室避光低溫保存。
生理指標測定于扦插后的第0、6、12、18、22、26、32、39、46、53天取樣,每次取樣均在9:00—10:00,共采樣10次。隨機選取20根生長狀態一致的插穗,取基部生根部位1~3 cm的韌皮部于50 mL試管中,立即放入液氮中冷凍,-80 ℃冰箱中保存待用。
1.3 插穗生根形態觀察
取樣時觀察并記錄插穗基部的外部形態變化,扦插60天后統計各處理生根率、生根數量、平均根長、最長根長等指標。長度精確到0.01 cm。生根率=生根插穗數/插穗總數×100%。
1.4 組織結構觀察
將軟化好的莖段流水沖洗 30 min,剪取基部1.0 cm左右材料,在真空組織脫水機(型號Excelsior AS)中脫水浸蠟,包埋機(型號JB-P5)中包埋,將修整好的蠟塊置于石蠟切片機(型號RM2016)切片,切片厚5 μm,組織攤片機(型號KD-P)40 ℃溫水展片,60 ℃烘箱(型號GFL-230)內烘干。依次在二甲苯、不同濃度的乙醇中脫蠟復水,用番紅-固綠染色,掃描電子顯微鏡(型號SU8100)下觀察采圖。
1.5 生理指標測定
參考莫文娟等(2020)用高效液相-串聯質譜法(High Performance Liquid Chromatography-tandem Mass Spectrometry,HPLC-MS/MS)測定莖段基部韌皮部中內源激素(IAA、 GA3、 ABA、 ZR)含量。測定參數為: BEH C18色譜柱(2.1 mm×100 mm,1.8 μm),流動相A為0.1%甲酸水溶液,B為乙腈,進行線性梯度洗脫,流速0.25 mL·min-1,進樣量5 μL,柱溫40 ℃。質譜條件為: 電噴霧電離源(ESI),正負離子電離模式; 離子源溫度500 ℃,正負離子源電壓分別為5 500 V和4 500 V,氣簾氣30 psi,霧化氣和輔助氣均為50 psi,采用多反應監測模式(MRM)進行掃描。吲哚乙酸氧化酶(IAAO)、多酚氧化酶(PPO)、過氧化物酶(POD)活性用比色法測定,可溶性糖、可溶性蛋白含量分別采用蒽酮法、考馬斯亮藍法測定(李合生等, 2000); 全氮含量采用凱氏定氮法(汪欣等, 2020)測定。每個指標均3次重復。
1.6 數據處理
采用Microsoft Excel 2016進行數據統計分析,利用SPSS20.0 for Windows(SPSS,USA)進行方差分析(ANOVA,P<0.05),使用Origin 2018軟件制作圖表。
2 結果與分析
2.1 杜仲良種嫩枝插穗生根的外部形態觀察
杜仲嫩枝插穗不定根形成過程如圖1所示。選取‘華仲6號’杜仲半木質化枝條作為插穗(圖1A)分別浸蘸杜仲專用生根劑和清水(對照)后扦插。第12天,插穗基部傷口基本愈合,出現褐色瘤狀突起,體積膨大(圖1B); 第22天,大部分插穗基部距切口1.0~3.0 cm處皮層突起并裂開,突起頂部呈半透明狀,其內部已誘導形成根原基(圖1C); 第26天,可在大部分插穗基部觀察到幼小白色不定根突破皮層,形成明顯的不定根結構(圖1 D); 扦插約40天,大多數插穗不定根已伸長至3.0~5.0 cm,并開始形成二級側根,插穗地上部位頂芽舒展,嫩葉數量增多(圖1E); 第53天,插穗新葉生長迅速,葉面積增加,不定根伸長速度加快,此時,插穗已經成為具有根、莖、葉的完整植株(圖1F)。由表1可知,處理組與對照組的插穗生根效果差異顯著(P<0.05)。處理組生根率為85.3%,比對照提高了60%; 平均每根插穗生根數達25.6根,比對照多20根; 平均根長8.41 cm,約為對照2倍。
2.2 杜仲良種嫩枝扦插不定根發育的解剖觀察
對插穗生根過程進行解剖觀察,扦插前杜仲莖的橫切面初生構造由外向內依次為: 表皮、皮層、韌皮部、維管形成層、木質部及髓(圖2A、E),未發現潛伏根原基,因此杜仲扦插生根屬于誘導生根類型。處理組扦插12天左右,大部分插穗基部形成層部位出現排列緊密,細胞質濃、細胞核大的薄壁細胞(圖2F),扦插18天進一步發育為團狀分生組織,根原基原始細胞分化形成根原基結構(圖2B); 扦插22~26天,根原基細胞繼續增殖、生長(圖2 G),形成圓錐狀(圖2C),發達的根冠為其內部分生組織提供保護,插穗基部切口以上3 cm范圍內皮層有密集的突起; 扦插1個月左右,大部分插穗不定根突破皮層、表皮伸出體外,橫切面為“個”字形(圖2 D),此時不定根已分化形成維管系統,并與插穗莖內維管組織相連(圖2 H)。
通過對杜仲生根解剖及形態變化觀察,可確定杜仲不定根主要由維管形成層外薄壁細胞誘導分化形成的根原基發育而來,扦插生根過程劃分為4個階段: 不定根誘導期(扦插0~12天)、啟動期(扦插12~22天)、表達期(扦插22~32天)和伸長期(扦插32~53天)。

圖1 杜仲嫩枝扦插生根的外部形態變化

表1 外源激素處理對杜仲嫩枝扦插根系形態指標及生根時間的影響①

圖2 杜仲嫩枝扦插不定根發生過程的解剖觀察
2.3 杜仲扦插生根進程中內源激素含量變化
處理組生長素(IAA)含量呈“上升—下降”的單峰變化趨勢,對照組峰值出現時間滯后(圖3A)。處理組插穗自扦插日起IAA含量快速升高,22天形成峰值(30.12 ng·g-1FW)后開始降低; 而對照組直到39天形成峰值,比處理組推遲了17天。對照組IAA峰值出現較晚,可見,IAA是誘導杜仲插穗不定根形成的關鍵內源激素。
處理及對照組赤霉素(GA3)含量呈“上升—下降—上升”變化趨勢(圖3B)。GA3分別在不定根誘導期和伸長期顯著升高,處理組GA3含量始終高于對照,有助于插穗抗性的提高和不定根的伸長。生根期間GA3濃度降低更利于根原基的形成和發育。GA3可促進水解酶的合成,有利于營養物質的供給,為不定根的啟動及表達做好物質準備。因此處理組GA3含量較高。
處理組插穗的脫落酸(ABA)含量整體呈“上升—下降”變化趨勢(圖3C)。處理組ABA含量在不定根誘導期升高至峰值(107.0 ng·g-1FW),對照組則降低至谷值(44.15 ng·g-1FW); 啟動期、表達期及伸長期,處理組ABA含量均持續降低; 對照組ABA含量在啟動期(第22天)上升至峰值(73.06 ng·g-1FW),表達期及伸長期持續下降。ABA主要抑制杜仲插穗不定根的發育,在扦插初期含量升高可提高插穗的抗性。
處理組插穗玉米素核苷(ZR)含量整體呈“下降—上升—下降”變化趨勢,且對照組始終低于處理組(圖3D)。插穗剛脫離母體,ZR的供應通路被切斷,而ZR的消耗利用正常進行,因此在扦插前期ZR含量逐漸下降; 處理組插穗ZR含量在表達期出現峰值,顯著高于對照,這可能與前期外源激素處理有關; 不定根伸長期,處理組ZR含量平緩走低,而對照組ZR含量在第39天降至谷值(1.66 ng·g-1FW)后回升,可能是由于對照組插穗整體生根進程較晚,此時正處于不定根開始形成階段。不定根形成前,ZR含量下降,而表達期ZR含量上升。說明根原基的誘導及分化需要低水平的ZR含量,而不定根的表達需要大量ZR參與。
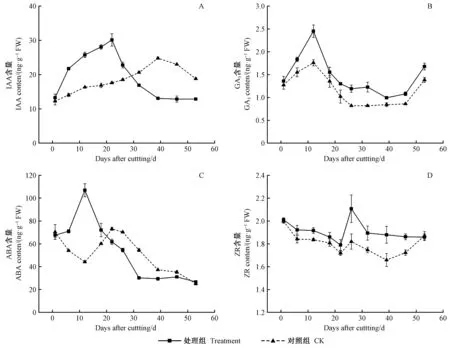
圖3 杜仲嫩枝插穗生根進程中內源激素變化

圖4 杜仲嫩枝插穗生根過程中氧化酶活性的變化
2.4 杜仲生根過程中氧化酶活性的變化
處理組吲哚乙酸氧化酶(IAAO)活性呈“降低—升高”的變化趨勢,對照組IAAO活性持續下降(圖4A)。不定根誘導及啟動期,處理組IAAO活性快速下降至谷值(4 654.52 U·g-1min-1),而對照組則下降趨勢較平緩,說明外源激素抑制了IAAO活性,IAAO活性降低的同時對應IAA含量的顯著升高,有利于根原基的誘導分化。在不定根表達及伸長期,處理組IAAO活性快速升高,39天達到峰值(6 795.12 U·g-1min-1); 對照組IAAO活性繼續下降緩慢,最后趨于穩定水平。IAAO活性下降,IAA含量升高,有利于不定根的發生; IAAO活性升高,IAA含量降低,有利于不定根的伸長。
過氧化物酶(POD)活性呈“上升—下降”趨勢,處理與對照變化趨勢一致(圖4B)。生根前,處理組插穗POD活性高于對照,生根后相反。杜仲生根劑處理在生根前顯著提高了插穗POD活性,使POD活性提前達到峰值,生根后抑制POD活性。這說明較高活性的POD有利于根原基的誘導及分化,低活性的POD則有利于不定根的表達及伸長。
多酚氧化酶(PPO)活性整體呈“上升—下降”變化趨勢,處理與對照組插穗中PPO變化趨勢相同(圖4C)。不定根誘導及啟動期PPO活性升高,處理組在第22天達到峰值(30.12 U·g-1min-1),而對照組升高較為緩慢。不定根表達及伸長階段(22~53天)PPO活性逐漸降低。扦插初期處理及對照插穗PPO活性無顯著差異,隨著不定根的發生,2組間達到顯著差異水平(P<0.05)。這說明高活性的PPO與不定根的發生關系密切。
2.5 生根過程中營養物質含量的變化
由圖5A可知,可溶性糖含量均呈“下降—上升—下降”的變化趨勢,對照與處理變化趨勢相同,但整體低于處理組。扦插前期,可溶性糖含量有下降趨勢,這可能與插穗愈合傷口快速消耗自身糖類物質有關。不定根形成期,處理組插穗的可溶性糖含量持續增加,始終高于對照組,該過程中插穗的細胞分化和代謝能力旺盛,相關酶的活性升高,可溶性糖含量積累; 處理組插穗扦插第39天可溶性糖含量升高到峰值(72.60 mg·g-1FW)。在生根后期,生根效果較好的處理組可溶性糖含量下降幅度更大,可能是處理組插穗生根數量較多,幼根有較強的代謝和呼吸作用,對糖類物質消耗較多所致。
由圖5B可知,可溶性蛋白含量整體呈“升高—下降”的變化趨勢,處理組峰值在第26天出現,比對照組提前13天。不定根誘導及啟動期,處理組插穗可溶性蛋白含量高于對照,這可能是由于插穗生理活動旺盛,需要大量蛋白酶類參與。表達及伸長期,處理組插穗可溶性蛋白含量快速下降,不定根的大量表達及伸長消耗了大量的可溶性蛋白。而對照組可溶性蛋白含量峰值出現時間較晚。調查發現處理組插穗多成爆發式根系,這可能與前期可溶性蛋白的積累有關。在生根前,經外源激素處理插穗可溶性蛋白含量升高較快,并提前達到峰值,可見可溶性蛋白的積累有利于插穗生根。
由圖5C可知,處理組插穗在生根期間全氮含量呈“上升—下降—緩慢上升”的變化趨勢,而對照整體變化不大。扦插后,處理組全氮含量快速升高,18天時達到峰值(6.06 mg·g-1FW),可能是外源激素的刺激提高了細胞代謝水平,從而促進氮素向下運輸并在基部大量積累,氮元素含量升高; 表達期,處理組全氮含量顯著降低至谷值(4.02 mg·g-1FW),氮素可合成蛋白質和核酸等物質為根原基細胞發育提供能量和物質,因此含量降低; 不定根伸長期,總氮含量略有上升,可能是插穗形成的新根能夠吸收營養物質,對自身氮素消耗減少。可見杜仲不定根形成過程中氮素化合物最先被消耗。

圖5 杜仲嫩枝插穗生根過程中營養物質含量的變化
3 討論
3.1 杜仲良種嫩枝扦插不定根發育特征
目前對扦插不定根發生形態解剖學研究主要集中在插穗的根原基類型、生根類型和莖的解剖結構3個方面(杜學梅等, 2019)。
根據插穗根原基形成方式及時間可分為潛伏根原基和誘導根原基(杜常健等, 2019a)。潛伏根源基在扦插前枝條本身就已存在,受離體刺激且條件合適時可繼續發育為不定根,如歐美雜種山楊(Populustremula×P.tremuloides)(周容濤等, 2013); 誘導根源基是在扦插前枝條中不存在根原基,在扦插后經外源激素或適宜條件誘導方可產生(杜偉, 2016)。在杜仲枝條中未發現潛伏根原始體,不定根是由扦插后維管形成層外的薄壁細胞誘導分化形成的根原基發育而來,屬于誘導根原始體類型。
根據插穗不定根形成部位可以劃分為4種生根類型: 皮部生根型、愈傷組織生根型、側芽或潛伏芽基部分生組織生根型和混合生根型(王濤, 1996)。杜仲插穗不定根絕大多數由基部皮層冒出,極少見到愈傷生根,杜仲以皮部生根為主,這與歐美雜種山楊(周容濤等, 2013)、側柏(Platycladusorientalis)(杜常健等, 2019a)和小葉芳樟(Cinnamomumcamphoravar.linaloolifera)(梁曉靜等, 2020)等結果相似。
根據生根解剖結構特征,通常把不定根的形成過程分為3個階段: 誘導期、啟動期、表達期(Danieletal., 2014; Lietal., 2008; Zhangetal., 2017; 張煥欣等, 2017; 周容濤等, 2013)。誘導期表示不定根起源細胞感知內外信號、提高代謝水平(Ahkamietal., 2009); 啟動期以細胞分裂、分生組織形成、根原基的發育為特征(Ramirez-Carvajaletal., 2009); 而表達期的特征是根原基的生長并且伸出表皮層,形成肉眼可見不定根(Syrossetal., 2004; 張煥欣等, 2017)。有研究認為,插穗內部靠近韌皮部的皮層中存在數層纖維細胞構成的厚壁細胞,對根原基的生長有機械阻礙作用,導致插穗生根困難(杜常健等, 2019b; 梁曉靜等, 2020; 張猛等, 2010)。王濤(1996)研究表明,在沒有環狀厚壁組織或厚壁組織不連續時,不定根發生較容易,反之生根困難。而厚壁組織可通過外源生長素刺激而打破(杜學梅等, 2019)。解剖發現在杜仲插穗皮層與韌皮部間存在連續的厚壁組織,阻礙生根。因此,杜仲皮層在生根前出現連續環狀的厚壁組織和單一的生根方式可能是造成杜仲扦插生根困難和生根時間久的重要原因。
3.2 杜仲扦插生根生理學研究
3.2.1 內源激素與生根的關系 不定根的發生是受多種內源激素共同參與、共同調控的復雜過程(杜學梅等, 2019; 張煥欣等, 2017)。IAA在不定根生長發育過程中起著核心作用,國內外學者已經從多個角度進行了論證(Cibeleetal., 2013; 張曉平等, 2004)。Taramino 等(2007)研究認為,高濃度的IAA可使根系分生組織細胞分裂次數增加,而低濃度的IAA可加快根系伸長區細胞的分化。在麻楝(Chukrasiatabularis)、北美香柏(Thujaoccidentalis)等扦插生根試驗中,均發現IAA含量在不定根形成的關鍵時期大幅度升高,在不定根長出后下降(王青, 2019; 劉明國等, 2010)。Naoki等(2011)發現藍桉(Eucalyptusglobulus)品種體內IAA含量是難生根品種的2倍。IAA能促進插穗生根,具體作用主要體現在: 營養物質調配利用、調節酶活性及分子水平信號傳導3方面(敖紅等, 2002; 吳文浩等, 2016; 王青, 2019; 曹凡等, 2016)。本研究中,杜仲處理組插穗內IAA含量在不定根誘導及啟動期大幅度升高,22天達到峰值,與解剖觀察到的根原基出現時間基本一致,與對照比,處理組插穗IAA快速積累,達到峰值,有利于根原基的誘導分化。處理組插穗IAA峰值出現的時間比對照提前了17天,IAA含量提前快速升高可能是插穗提前生根的主要原因。因此認為IAA是促進杜仲插穗不定根形成的主要內源激素(歐陽芳群等, 2015)。
GA3主要促進細胞分裂和伸長,對不定根的形成有抑制作用。在不定根的誘導期下降明顯(董勝君等, 2013; 劉明國等, 2010)。但也有研究認為GA3含量上升與插穗愈傷組織的誘導、不定根的形成呈正相關性(李朝嬋等, 2012; 趙云龍等, 2013)。可見GA3對扦插生根的影響因樹種差異而不同。本研究發現杜仲插穗GA3含量在不定根發生的關鍵時期呈下降趨勢,這與山杏(Armeniacasibirica)、芽變毛白楊(Populustomentosa)、興安圓柏(Juniperusdavurica)扦插過程中GA3的變化趨勢(遲彥等, 2007; 董勝君等, 2013; 郭英超等, 2012)一致。生根過程中處理組插穗GA3含量始終高于對照,可能是外源激素處理在一定程度上提高了插穗基部GA3的含量,進而促進了淀粉、蛋白質的水解,促進不定根的生成。ABA可增強植物抗逆性,被稱為脅迫激素,主要抑制扦插生根(侯江濤等, 2019)。在插穗剛脫離母體時,處理組ABA含量快速升高,而對照組含量卻先降低后升高,處理組在不定根誘導期內ABA與GA3含量變化趨勢一致,說明二者在感知受傷信號、提高插穗抗性方面具有協同作用,這與糙葉杜鵑(Rhododendronscabrifolium)結果(趙云龍等, 2013)一致。
ZR是木本植物體內細胞分裂素的主要運輸形式,具有促進細胞分裂、分化和影響不定根形成的作用(Ramirez-Carvajaletal., 2009; Druegeetal., 2016)。許多研究認為,低濃度的ZR有利于插穗不定根的形成與發育,例如茶條楓(Acertataricumsubsp.ginnala)和北美香柏的嫩枝扦插(蔡楚恒, 2019; 劉明國等, 2010)。ZR對無患子(Sapindusmukorossi)扦插過程的生理作用較復雜,低含量的ZR有利于根原基的發生,但高含量的ZR促進不定根形成(韓繼紅等, 2019)。在杜仲插穗生根早期,ZR含量持續降低,有利于根原基的形成和分化。不定根表達期ZR含量大幅度升高有利于不定根的生長發育,這與無患子、長柄扁桃(Amygdaluspedunculata)等(韓繼紅等, 2019; 李斌, 2017)研究結果一致。
3.2.2 氧化酶活性與生根的關系 吲哚乙酸氧化酶(IAAO)、過氧化物酶(POD)、多酚氧化酶(PPO)是與植物扦插生根關系最為密切的3種氧化酶類(Naokietal., 2011; 杜偉, 2016; 侯江濤等, 2019)。IAAO通過調節插穗內IAA含量水平,從而影響其不定根的形成和發育(張煥欣等, 2017; 吳文浩等, 2016)。在不定根誘導階段,較低活性的IAAO有利于插穗IAA的積累,以促進不定根的誘導和發生(Nagetal., 2001)。宋金耀等(2007)研究發現,易生根樹種扦插后IAAO活性基本均呈下降趨勢,而難生根樹種則變化趨勢相反,因此,IAAO活性的降低可使IAA含量升高,進而對根的形成過程有促進作用。本研究中,處理組杜仲插穗前期IAAO活性大幅度下降,正好對應IAA含量的大幅度升高。IAAO 活性變化與 IAA 含量變化高度吻合,呈此消彼長的關系,這與歐洲甜櫻桃(Prunusavium)、北美香柏、茶樹(Camelliasinensis)等扦插結果(劉明國等, 2010; 魏海蓉等, 2013; Rout, 2006)類似。PPO和POD參與植物體內多種生理生化過程。在細胞分裂分化、根原基的形成和生長以及逆境環境適應過程中起重要作用(Hayrullahetal., 2003)。POD參與生長素的代謝、呼吸、細胞壁的合成和傷害反應,與不定根的誘導和生長密切相關,活性升高是植物具有生根能力的標志(Barceloetal., 1990; 杜學梅等, 2019; Rout, 2006)。研究證明,在生根過程中,堿性和酸性POD先后被激活用于誘導生根和發育(Gasparetal., 1985; 杜學梅等, 2019)。扦插初期插穗感知受傷信號,POD活性升高,不僅響應逆境脅迫,還可清除插穗內部自由基等物質,由此認為扦插生根前高水平的POD活性對于生根有利(趙云龍等, 2013; 李斌, 2017; 魏海蓉等, 2013)。酚類物質抑制生根,PPO能夠催化酚類物質合成一種生根的輔助因子—“IAA-酚酸復合物”以促進不定根形成(鄭巧巧等, 2020)。PPO可在不定根起始部位影響細胞分裂分化及根原基的形成和生長(張煥欣等, 2017)。處理組杜仲插穗POD和PPO活性在前期升高,峰值出現時間正好對應插穗不定根發育的關鍵時期,不定根伸長階段下降。說明杜仲生根劑提高了POD和PPO的活性,有效促進插穗生根(王新建等, 2009)。這與葡萄(Vitisvinifera)、歐洲甜櫻桃扦插的結果(Hayrullahetal., 2003; 魏海蓉等, 2013)一致。
3.2.3 營養物質與生根的關系 插穗生根是一個需要消耗大量營養物質和能量的過程(張煥欣等, 2017)。可溶性糖、可溶性蛋白及含氮化合物作為能量及物質基礎供應的主要來源,與根原基的形成關系密切(侯江濤等, 2019)。可溶性糖可直接作為呼吸底物被利用,為插穗生根提供必須的物質和能量(Husenetal., 2007; 張煥欣等, 2017); 可溶性蛋白為細胞生長提供物質基礎,在扦插生根過程中一方面提供插穗生根所需的能量,另一方面在不定根發育過程中以蛋白酶類的形式參與調控多種生理生化過程(曹凡等, 2016; 王新建等, 2009); 氮元素化合物可誘導根原基的發生并為不定根的發育中核酸、蛋白質的合成提供原料(王青, 2019)。許多研究認為營養物質在生根前需要有一個積累的過程,在生根過程中被大量消耗(易詠梅, 2001; 陳寧, 2011; 許曉崗等, 2013)。杜仲插穗生根過程中全氮含量和可溶性蛋白含量先后在第18天和22天出現峰值,可能與氮類化合物是蛋白質的合成原料有關。可溶性糖含量在第39天達到峰值后才開始下降,生根數量較多的處理組下降幅度更大,可能與大量不定根自身較強的代謝和呼吸作用消耗大量糖類有關(王新建等, 2009; 陳寧, 2011)。
4 結論
杜仲不定根發生屬于皮部誘導生根類型,根原基來源于形成層。其生根過程可劃分為誘導期、啟動期、表達期和伸長期4個階段。杜仲扦插生根困難的主要原因之一是內源IAA含量低,抑制了根原基起始細胞的誘導。ABA雖抑制生根,但在不定根誘導期參與信號傳導,提高插穗抗性; ZR在不定根表達期含量顯著升高; GA3則主要在扦插初期和不定根伸長期發揮作用。POD和PPO活性整體呈“升高—下降”趨勢,IAAO活性整體呈“下降—升高”趨勢。杜仲生根劑可提高POD和PPO活性、抑制IAAO活性。可溶性糖、可溶性蛋白、氮素化合物主要表現為在生根前不斷積累,含量升高,在生根后因大量消耗而含量降低。
