lncRNA01535通過JAK/STAT3通路調節毛囊干細胞增殖和分化的作用研究*
2022-05-07 11:10:16姜金豆周建敏董溫云黃亞奔王佳雯
重慶醫學 2022年8期
關鍵詞:水平
朱 君,姜金豆,周建敏,董溫云,黃亞奔,王佳雯
(1.溫州醫科大學溫州市第三臨床學院/溫州市婦幼保健院/溫州市人民醫院整形外科,浙江溫州325000;2.廣東省婦幼保健院醫療美容科,廣州 510010)
毛囊干細胞(HFSC)屬于成年干細胞,在靜態下顯示出較強的增殖能力[1]。研究發現,HFSC具有多向分化潛能,可以分化為皮膚、毛囊、皮脂腺,并參與皮膚傷口的愈合過程[2]。研究表明,HFSC可以通過靶向Janus激酶(JAK)來誘導毛囊生長,HFSC通過上調信號轉導與轉錄激活子3(STAT3)激活而分化為轉運擴增細胞[3]。另1項研究表明,HFSC的體內轉錄調控可能受STAT3調控子調控:miR-128通過靶向STAT3來調節毛囊間充質干細胞向平滑肌細胞的分化[4]。長鏈非編碼RNA(lncRNA)是指一類長度大于 200 nt但不編碼蛋白的RNA分子,已被確定在細胞的增殖和分化中起作用。近年來,lncRNAs已成為許多人類疾病研究的重點,包括代謝和遺傳性疾病、癌癥和人類干細胞分化[5]。有證據表明,lncRNAs與細胞信號通路轉導相關,也是細胞療法的靶標[6-7]。分子信號傳導機制已證實lncRNA通過調節HFSC的不同信號途徑與多種細胞代謝過程相關[8]。截至目前,尚未見lncRNA01535在HFSC中的表達及其調控HFSC分化的機制研究,本研究擬分析lncRNA01535對HFSC增殖和分化的調控作用及JAK/STAT3通路在其中發揮的作用,為臨床皮膚病的治療提供理論依據。
1 材料與方法
1.1 主要試劑及儀器
胎牛血清、青霉素、鏈霉素、10%十二烷基鈉硫酸-聚丙烯酰胺凝膠(SDS-PAGE)、JAK一抗(1∶1 000)、STAT3一抗(1∶1 000)、GAPDH一抗(1∶2 000)購自美國Sigma公司(批號:ER-54863、KJ-5874.63、BV-5489.64、VB-471.65、F1555.52、G584.63、V6363.52),lncRNA01535 mimics、lncRNA01535 inhibitor購自武漢金開瑞生物公司(批號:XS-474.36、VF-748.36),DMEM培養基購自美國Hyclone公司(批號:DE-5596.56);Lipofectamine2000購自美國Thermo Fisher Scientific公司(批號:WR-474.26),細胞計數試劑盒8(CCK-8)、PrimeScript RT Master Mix試劑盒購自日本希森美康公司(批號:XC469.52、ZA-478.36);油紅O、細胞裂解液購自上海碧云天生物科技公司(批號:XQ-5858.45、XC-58.65);膜聯蛋白V-異硫氰酸熒光素(FITC)/碘化丙啶(PI)細胞凋亡檢測試劑盒購自美國Vazyme公司(批號:XE-471.36);細胞周期檢測試劑盒購自武漢AmyJet Scientific公司;TRIzol試劑盒購自美國Invitrogen公司(批號:SC-458.36),SYBR Select Master Mix、ABI 7300系統購自美國Applied Biosystems公司(批號:ED-585.36、EY-4741.65);聚偏氟乙烯(PVDF)膜、增強的化學發光(ECL)顯影劑購自美國Millipore公司(批號:MN-69853.2、DC-4523.54);辣根過氧化物酶標記的山羊抗兔二抗(1∶10 000)購自美國Merck KgaA公司(批號:W098.34)。Synergy Mx-89多模式酶標儀購自日本希森美康,WE-98流式細胞儀購自美國BD Biosciences公司,NanoDrop紫外分光光度計購自美國Thermo Fisher Scientific公司。
1.2 細胞培養及分組
HFSCs細胞系購自中國科學院上海典型培養物保藏中心,在37 ℃、5% CO2、2% O2、93% N2培養箱中,用10%胎牛血清、1%青霉素和鏈霉素(100 IU/mL)的DMEM培養基(美國Hyclone)培養。細胞分為HFSCs組、lncRNA01535 mimics組、lncRNA01535 inhibitor組。HFSCs組細胞不做任何處理;lncRNA01535 mimics、lncRNA01535 inhibitor組細胞分別轉染10 mL lncRNA01535 mimics、10 mL lncRNA01535 inhibitor。轉染方法:取10 mL HFSCs液(細胞濃度為5×106/mL)接種于6孔板,分別取100 pmol的 lncRNA01535 mimics、lncRNA01535 inhibitor于200 μL DMEM培養基中,再加4 μL Lipofectamine2000混勻,室溫孵育30 min,無菌PBS洗滌細胞,將上述混合液加入細胞孔內,6 h 后棄混合液,加入含10%胎牛血清的DMEM培養基培養。
1.3 細胞增殖存活的測定
CCK-8試劑盒分析細胞增殖。取對數期生長的HFSCs,將細胞以每孔5 000個細胞的密度接種于96孔板中,72 h后,將10 μL CCK-8溶液添加至每個孔中。在37 ℃下孵育1.5 h后,在酶標儀于450 nm處測量吸光度(A)值,存活率=(實驗組A值-HFSCs組A值)/(實驗組A值-蒸餾水空白調零組A值)×100%。
1.4 細胞分化水平測定
將HFSCs以5 000/孔的細胞密度接種于96孔板中。將細胞保持在1%瓊脂包被上,分化18 d。10%甲醛溶液固定細胞1 h,隨后用60%油紅O染色10 min。分化率=油紅O陽性細胞的比例/總細胞數×100%。
1.5 細胞周期、凋亡水平測定
將HFSCs與雙重染色FITC/PI細胞凋亡檢測試劑盒混合,在室溫下孵育10 min,上流式細胞儀分析細胞凋亡情況。根據細胞周期檢測試劑盒的操作說明,將收集的細胞在70%乙醇中固定過夜。將細胞與100 μL PI于4 ℃避光孵育30 min,上流式細胞儀檢測細胞周期分布。
1.6 細胞lncRNA01535、JAK、STAT3 mRNA水平測定
TRIzol法提取細胞總RNA,并使用NanoDrop紫外分光光度計測量RNA的純度和濃度。通過PrimeScript RT Master Mix將1.5 μg RNA反轉錄為cDNA,然后于-20 ℃保存。使用SYBR Select Master Mix在ABI 7300系統中進行實時熒光定量PCR(RT-qPCR)反應,GAPDH的表達水平作為標準化的內參。相關mRNA的表達水平通過2-ΔΔCt法計算。引物購自廣州富倫根有限公司,引物序列如下:GAPDH正向5′-CCA GCC GAG CCA CAT CGC TC-3′,反向5′-ATG AGC CCC AGC CTT CTC CAT-3′;lncRNA01535正向5′-GGG CGG CAG GTC ACT GAC AC-3′,反向5′-GCC AGC AGC CGC TGG CTT AG-3′;JAK正向5′-GCA ATA CTC GCC TTA CGG CT-3′;反向5′-TAC ACA CCT TGT AGT ACG CC-3′;STAT3正向5′-ACA ACT TTG GTA TCG TGG AAG G-3′,反向5′-ATG AGC CCC AGC CTT CTC CAT-3′。
1.7 細胞JAK、STAT3蛋白水平測定
各組HFSCs用細胞裂解液裂解以提取總蛋白。BCA法測定蛋白濃度。通過水浴變性后,加入蛋白上樣緩沖液,并使用10% SDS-PAGE電泳分離蛋白,將蛋白質轉移至PVDF膜上,并在室溫下用5%牛血清清蛋白封閉1 h。隨后,添加JAK、STAT3、GAPDH一抗在4 ℃下孵育過夜,然后添加辣根過氧化物酶標記的山羊抗兔二抗,在室溫下孵育2 h,然后借助ECL顯影劑進行顯影。使用ImageJ軟件(美國馬里蘭州國立衛生研究院)量化條帶強度。
1.8 統計學處理
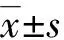
2 結 果
2.1 各組HFSCs細胞存活率比較
與HFSCs細胞組[(60.85±9.56)%]比較,lncRNA01535 mimics組細胞存活率[(73.54±9.26)%]升高,lncRNA01535 inhibitor組細胞存活率[(52.21±5.33)%]降低(P<0.05);與lncRNA01535 mimics組比較,lncRNA01535 inhibitor組細胞存活率降低(P<0.05),差異均有統計學意義。
2.2 各組HFSCs細胞分化率、凋亡率及G1期細胞比例比較
與HFSCs細胞組比較,lncRNA01535 mimics組細胞分化率升高,細胞凋亡率和G1期細胞比例降低(P<0.05),lncRNA01535 inhibitor組細胞分化率降低,細胞凋亡率和G1期細胞比例升高(P<0.05);與lncRNA01535 mimics組比較,lncRNA01535 inhibitor組細胞分化率降低、細胞凋亡率和G1期細胞比例升高(P<0.05),差異均有統計學意義。見表1,圖1、2。

表1 各組HFSCs細胞分化率、凋亡率及G1期比較
2.3 各組HFSCs細胞lncRNA01535、JAK、STAT3 mRNA表達水平比較
與HFSCs細胞組比較,lncRNA01535 mimics組lncRNA01535、JAK、STAT3 mRNA表達水平升高(P<0.05),lncRNA01535 inhibitor組lncRNA01535、JAK、STAT3 mRNA表達水平降低(P<0.05);與lncRNA01535 mimics組比較,lncRNA01535 inhibitor組細胞lncRNA01535、JAK、STAT3 mRNA表達水平降低(P<0.05),差異均有統計學意義。見表2。

A:HFSCs細胞組;B:lncRNA01535 mimics組;C:lncRNA01535 inhibitor組。

A:HFSCs細胞組;B:lncRNA01535 mimics組;C:lncRNA01535 inhibitor組。

表2 各組HFSCs細胞lncRNA01535、JAK、STAT3 mRNA表達水平比較
2.4 各組HFSCs細胞JAK、STAT3蛋白表達水平比較
與HFSCs細胞組比較,lncRNA01535 mimics組細胞JAK、STAT3蛋白表達水平升高(P<0.05),lncRNA01535 inhibitor組細胞JAK、STAT3蛋白表達水平降低(P<0.05);與lncRNA01535 mimics組比較,lncRNA01535 inhibitor組細胞JAK、STAT3蛋白表達水平降低(P<0.05),差異均有統計學意義。見表3,圖3。
表3 各組HFSCs細胞JAK、STAT3蛋白表達水平 比較

A:HFSCs細胞組;B:lncRNA01535 mimics組;C:lncRNA01535 inhibitor組。
3 討 論
成體干細胞可以替代垂死的細胞并盡快修復受損的組織,因此在維持組織穩態方面起著至關重要的作用。HFSC對于毛囊的產生、維持和更新至關重要。除了體內正常的生理功能外,HFSC還被認為是深層皮膚損傷,組織工程和再生醫學中傷口組織修復必不可少的干細胞。HFSC是血管工程和干細胞療法的重要來源。更重要的是,HFSC具有增殖、集落形成、再生的特征,此外HFSC易于獲取且侵入性小,因此備受關注。闡明HFSC增殖,分化,凋亡的分子機制至關重要。
LncRNA已證明在HFSCs多系分化進程中具有潛在作用。LncRNA PCAT1通過調節miR-329/Wnt10b軸參與間充質干細胞分化[9];LncRNA5322通過PI3K/AKT信號途徑調節毛囊干細胞增殖和分化[10];lncRNA-H19被證明通過充當miR-199a-5p的內源性競爭RNA(ceRNA)來促進間充質干細胞的存活和血管生成能力[11]。有關基因治療和新型傷口治療的研究報道表明,有必要將表皮細胞和HFSCs視為不同的種群,lncRNA01535可以通過充當miR-19b-3p的誘餌來促進HFSCs的增殖[12];此外,lncRNA01535可促進毛囊源性神經干細胞向雪旺細胞分化[13]。本研究分析了lncRNA01535對HFSCs增殖和分化的調控作用。結果顯示:與HFSCs細胞組比較,lncRNA01535 mimics組、細胞存活率、細胞分化率升高,細胞凋亡率、G1期細胞比例降低;lncRNA01535 inhibitor組細胞存活率、細胞分化率降低,細胞凋亡率、G1期細胞比例升高(P<0.05);與lncRNA01535 mimics組比較,lncRNA01535 inhibitor組細胞存活率、細胞分化率降低,細胞凋亡率、G1期細胞比例升高(P<0.05)。這表明lncRNA01535過表達刺激了HFSCs的增殖和分化,抑制了HFSCs凋亡,而lncRNA01535表達抑制了HFSCs的增殖和分化,促進了HFSCs凋亡;表明lncRNA01535可調控HFSCs的增殖和分化。
為了研究lncRNA01535調節HFSCs增殖和分化的機制,本研究評估了JAK/STAT3通路。結果表明,在lncRNA01535 mimics轉染的HFSCs中JAK、STAT3基因與蛋白的表達顯著增加。有研究報道JAK能夠調節人骨髓干細胞中骨形態發生蛋白2誘導的β-catenin活化[14]。還有研究表明JAK/STAT3通路在人類神經干細胞向神經元分化中起關鍵作用[15]。本研究結果顯示,與HFSCs細胞組比較,lncRNA01535 mimics組lncRNA01535 mRNA,JAK、STAT3 mRNA和蛋白表達水平升高,lncRNA01535 inhibitor組細胞lncRNA01535 mRNA,JAK、STAT3 mRNA和蛋白表達水平降低(P<0.05);與lncRNA01535 mimics組比較,lncRNA01535 inhibitor組細胞lncRNA01535,JAK、STAT3 mRNA和蛋白表達水平降低(P<0.05)。這說明lncRNA01535過表達導致HFSCs中JAK/STAT3通路的表達上調。JAK是胚胎成纖維細胞中lncRNA01535的直接靶基因[16]。此外,已有研究證明lncRNA01535可以與JAK特異性結合以調節人間充質干細胞的成骨分化[17]。此外,在肝細胞癌(HCC)中發現lncRNA01535表達與JAK表達呈正相關,并且JAK升高可預示HCC患者的預后不良[18]。此外已有研究證明lncRNA01535靶向STAT3參與人毛囊來源的間充質干細胞(HF-MSC)向平滑肌細胞(SMC)的分化過程,可作為SMC的組織工程和再生醫學的潛在靶標[19]。綜上所述,lncRNA01535促進HFSCs的增殖和分化;其機制可能與lncRNA01535激活HFSCs中JAK/STAT3通路有關。
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年6期)2019-10-08 08:55:48
人大建設(2019年12期)2019-05-21 02:55:32
雜文月刊(2018年21期)2019-01-05 05:55:28
人大建設(2017年6期)2017-09-26 11:50:44
學苑創造·A版(2015年11期)2016-01-14 09:03:27
俄羅斯問題研究(2012年1期)2012-03-25 09:54:45
中國火炬(2010年12期)2010-07-25 13:26:22
中國火炬(2010年8期)2010-07-25 11:34:30
