纖維素的微生物降解研究進展
2022-04-26 10:38:18李明源王繼蓮
食品工業科技 2022年9期
李 靜,李明源,王繼蓮,張 甜,周 茜
(喀什大學生命與地理科學學院,葉爾羌綠洲生態與生物資源研究高校重點實驗室,新疆帕米爾高原生物資源與生態重點實驗室,新疆 喀什 844000)
纖維素是由葡萄糖以β-1,4糖苷鍵連接而成的線性鏈多糖[1],結構緊密、排列有序,具有高結晶度和難溶性等特點[2],是植物細胞壁的主要組成成分(圖1)。纖維素類物質是世界上年產量最多的二次資源[3],每年通過光合作用合成的有機物約有1×1011t,而纖維素就占50%,它們的再利用對社會的可持續發展和環境保護具有重要意義[4]。
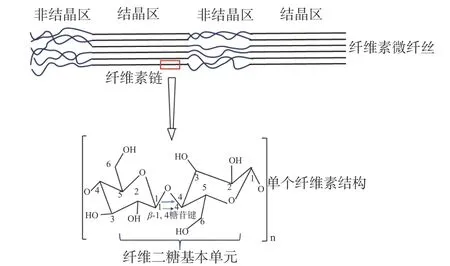
圖1 纖維素結構示意圖[5]Fig.1 The structure of cellulose[5]
纖維素的預處理是目前制約生物質資源高效利用的技術瓶頸之一。物理方法可以有效降低纖維素的結晶度,包括破碎、高溫熱解、微波處理和高能輻射等方式,具有操作簡單、污染少的優點,但同時能耗大,成本高。化學方法操作繁瑣程度較低,短時間就可達到降解效果,方式主要包括酸降解、堿降解、有機溶劑預處理和氧化預處理等。但所需試劑濃度較高,且后續需進行pH調節,生成的化學殘留物也會對環境造成污染,因此處理過的酸堿必須進行回收處理[6]。而微生物預處理是利用高產纖維素酶降解菌對其進行酶解,具有環保、專一性強、處理成本低、反應條件溫和、能耗低等優點,備受科研人員關注[7?8]。研究者們相繼開展了纖維素酶生產菌的選育及降解特性研究,得到了一批寶貴的菌種資源[9]。本文總結歸納了纖維素降解菌的篩選方法和研究策略,探討了各策略的優勢特點和應用價值,并闡釋了纖維素的微生物降解機理,對未來如何進一步高效利用纖維素進行了展望,以期為纖維素類物質的降解提供進一步理論支撐,促進二次資源的綜合利用。
1 篩選單一菌株降解纖維素
環境中存在著豐富的纖維素降解菌,主要包括真菌、放線菌及細菌等。它們的類群龐雜,相互協調,但絕大多數難以進行人工培養。目前已分離200多種,主要來源于枯腐樹葉或衣服、秸稈還田的土壤、反芻動物糞便等,多使用單一碳源(濾紙條或羧甲基纖維素鈉CMC-Na)進行富集培養[10?11]。由表1可見,降解菌種主要包括青霉屬(Penicillium)、曲霉屬(Aspergillus)、鏈霉菌屬(Streptomyces)、假單胞菌(Pseudomonas)、芽孢桿菌(Bacillus)等,以真菌居多,不同菌種對底物的降解效率參差不齊。

表1 單一菌株的纖維素降解特性Table 1 Characteristics of cellulose degradation activity of single strain
纖維素分子間主要靠氫鍵連接,鍵能相對較弱,其降解主要靠微生物產生的纖維素酶(包括外切β-1,4葡萄糖苷酶、內切β-1, 4葡萄糖苷酶和β-葡聚糖苷酶)破壞其氫鍵和β-1, 4糖苷鍵,將其多結構變為單結構。但纖維素酶活力受到諸多因素的影響,與培養溫度和培養時間密切相關,溫度過高會造成菌株提前失活[12],微生物培養基的pH、營養成分比例、接種量等也會對酶活力產生明顯影響,影響降解率。而相關研究報道證明通過向培養基中添加一定量的Fe2+和VB12等附加因子,可以起到穩定酶空間構象及提高微生物代謝效率的作用[20],有效提高纖維素降解率。此外,培養基中可添加適宜的產酶誘導物也可以提高纖維素酶產量,加速纖維素降解。槐糖是已報道誘導力最強的產纖維素酶誘導物,但大規模生產不具備經濟可行性。而以稻草粉代替槐糖作為碳源和誘導物,采用固體發酵法優化綠色木霉的產酶條件,纖維素酶活力得到有效提升[21]。
近年來,環境耐受性良好的高效降解纖維素菌株也被逐步發掘。某些冷適應降解菌的產酶條件經優化后在低溫下可保持高效產酶能力。放線菌Clavibacter michiganensisCF11在低于10 ℃、pH為6.5、培養時間為10 d的條件下纖維素酶活力顯著提升[22]。而一些耐高溫纖維素酶菌株具有高溫偏好性,熱穩定性強,在高溫環境下的纖維素降解能力較強[23]。如解淀粉芽孢桿菌Bacillus methylotrophicusY3-2在55 ℃條件下,不僅能高效降解纖維素,還具有抑制大腸桿菌、金黃色葡萄球菌等病原菌的能力,具有潛在應用前景[24]。但超高溫好氧發酵技術過程中會流失較多的營養成分,如何在高效降解和保持較多的營養成分之間找一個平衡點,有待深層次研究[25]。
2 微生物協同降解纖維素
單一菌株生成的纖維素酶種類相對單一,產酶能力薄弱,降解能力有限。1950年,Gilligan[26]首次發現木霉所產的纖維素酶可通過協同作用,實現對纖維素的高效降解。微生物協同降解的優勢在于產酶種類多、耗時少、降解覆蓋率高、針對性強[27],但對培養條件、培養基成分、比例等的要求相對嚴格。工業生產中,常利用微生物的協同作用達到降解纖維素作物的目的[28]。
2.1 篩選復合微生物菌系
直接從自然界篩選的天然復合菌群,基于微生物間的協同作用能有效增強纖維素降解速度,不僅擁有健全的酶系,還可以將不同pH的培養液緩沖到中性,更適合菌系生存。崔宗均等[29]采用酸堿反應互補原則,在傳代時重新優化組合馴化出的復合菌系MC1可在72 h內將濾紙和脫脂棉等較純的纖維素材料完全降解,對麥稈、鋸末等材料也有較強的降解能力。
復合體系中微生物產生的纖維素酶可降解纖維素生成葡萄糖,起到為菌供能、促進增殖等作用,多組分酶體系在降解過程中互補互利,共同完成降解過程。而復合菌系的構建增加了酶的環境適應性和功能穩定性,彌補了單一菌種的纖維素酶種類不全和纖維素酶配比不協調等缺點,從而提升降解速度。因此可以說,各組成菌之間、組成菌與纖維素酶、各酶與酶之間的協同和拮抗作用,是保持復合菌系穩定的重要機制。即使混入其它雜菌,也很難將這個平衡系統破壞。
復合菌系雖然在降解效果上優于單一菌株,但降解過程中出現的不確定因素也較多,如微生物群落和代謝類型不明確、高溫或低溫環境中部分酶系易失活、菌系的組成和結構會因培養條件的差異而改變等。因此,要想達到快速、低成本降解纖維素的目的,高效降解菌的篩選還存在一些亟待解決的瓶頸問題:a.明確復合菌系中菌株組成類別,進而準確判斷菌株習性,構建最優培養條件;b.深入解析各酶組分之間的重要協同作用關系;c.解析復合菌系中的代謝調控;d.增強對環境耐受性的復合菌系的研究。現階段可對菌株培養條件(溫度、pH、培養時間等)進行變量-非變量研究,觀察纖維素降解效率及纖維素酶活峰值的變化,從而確定菌群的最優培養條件[30?31]。或重新組合已有的復合菌群和功能菌群,檢測、對比新復合菌系的降解效率、結構穩定性及功能性,從而進一步明確復合體系中各組分間的協同作用關系。此外,還可聯合生物工程技術構建基因工程菌,將難培養的微生物中的纖維素酶編碼基因克隆到大腸桿菌或釀酒酵母上進行表達,以期獲得產率更大、酶系更完整的高活性降解菌。而通過分子生物學技術提取復合菌群的微生物總DNA,從構建的基因組文庫中篩選高產纖維素酶基因,也可提升降解菌系的結構和功能分析效率。
2.2 優化組合純培養菌株
將單一高活性產酶菌株混合培養,基于不同微生物類群的優化組合可以彌補菌種間的性質差異,所構成的完全酶系可充分發揮各酶系間的協同作用,綜合提高酶活性,最大限度發揮纖維素降解潛力,毫不遜于直接篩選到的復合菌系(表2)。多數學者對霉菌的組合研究較多[32],對細菌和放線菌的協同作用鮮見報道。細菌的基因無內含子,可直接克隆目的基因,方便構建高效纖維素酶工程菌[33]。而放線菌作為土壤微生物的重要組成部分,參與了大部分高分子聚合物的降解,還可以產生具有抑制雜菌生長的抗菌素[34]。因此,構建細菌與放線菌的復合菌系具有廣闊的研究前景。此外,菌株的培養環境也會深刻影響降解效果,原位生長環境的模擬能顯著提升其降解效果。
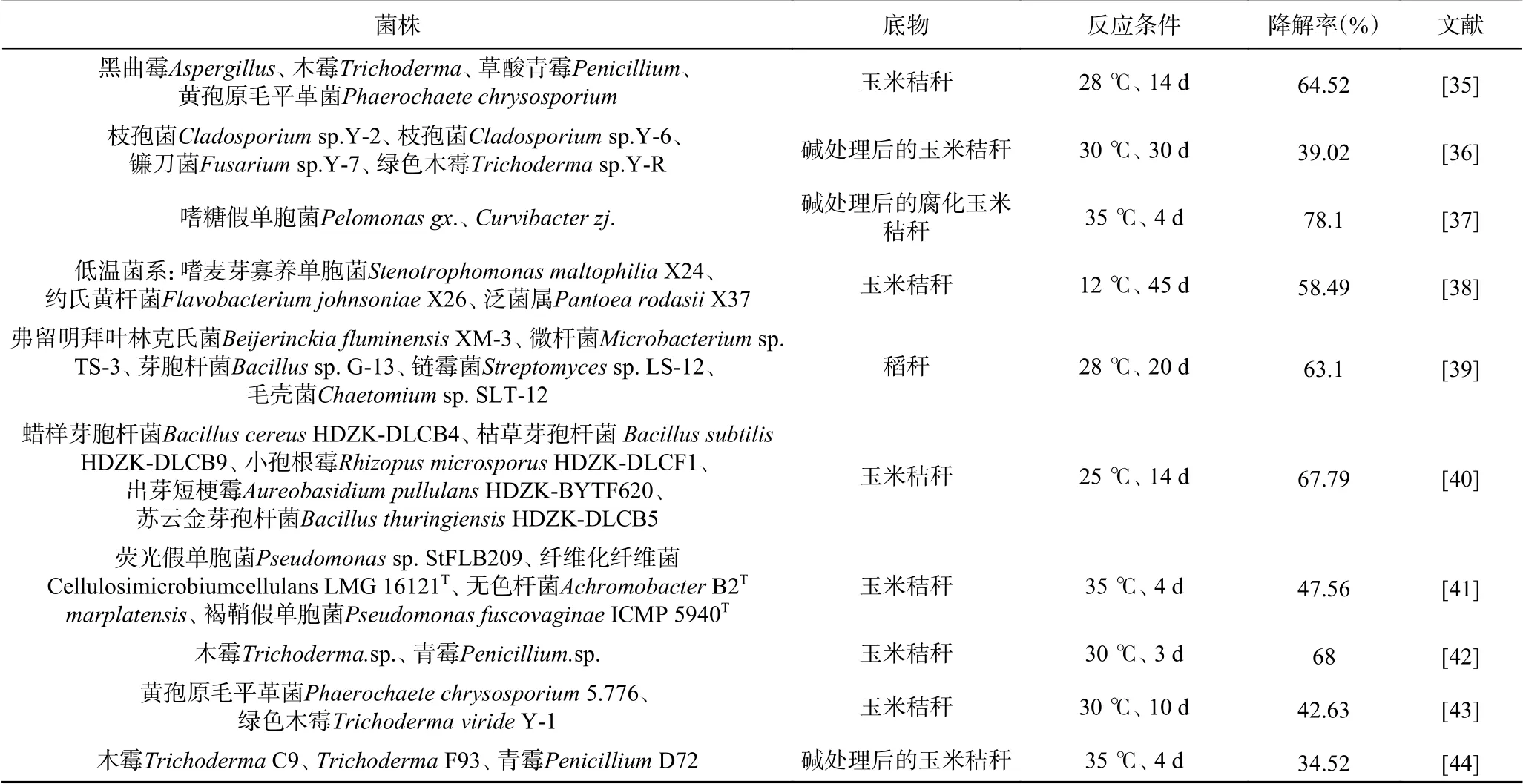
表2 組合菌株的纖維素降解特性Table 2 Characteristics of cellulose degradation activity of mixed strains
針對分離到的不同單一菌株,總結其相互間的拮抗活性、酶種類和代謝產物,將具有互補差異的幾種高酶活菌株進行復配,介導菌株之間的協同作用可起到提高菌系的降解能力和穩定性目的。同時此類復合體系每個菌株的特性明確,在實際應用中能較好地控制其活性,可操作性強。但缺陷在于優化組合單一菌株的過程相比天然復合菌系繁瑣復雜。另外需注意的是,并不是所有混合菌的發酵一定比單菌株發酵能力強,因復合菌系之間可能存在競爭或拮抗作用,不利于菌株的生長和產酶。因此,在進行不同組合的菌株培養時,要充分考慮菌株間的拮抗作用、物種組成和比例、菌株的生長周期和整體酶系的穩定等因素[45]。
3 微生物降解纖維素機理
微生物直接通過胞吞方式將纖維素等大分子物質轉移至細胞膜內進行代謝是很難實現的,因此,其對纖維素的降解主要靠分泌至胞外的纖維素酶氧化分解。由于纖維素結構的復雜性,其確切的降解機理至今尚無定論。目前被大家普遍接受的主要有3種理論[46],分別是C1-CX假說、協同理論和原初反應假說。
3.1 C1-CX假說
此假說認為在纖維素降解過程中,纖維素酶與各個酶或物質要依據一定的步驟順序才能完成整個水解過程[47]。相關研究表明,需要C1酶(外切β-1,4葡萄糖苷酶)、CX酶(內切β-1, 4葡萄糖苷酶)和CB酶(β-葡聚糖苷酶)同時存在才能將天然纖維素水解為葡萄糖。如果在降解結晶纖維素時首先加入C1酶,反應過后除去C1酶的同時再加入CX酶,則反應過程無法完成。根據C1-CX假說,先是結晶纖維素在C1酶的作用下將已形成結晶結構的纖維素鏈斷裂,使長鏈分子的末端脫離,形成非結晶無定型纖維素或可溶性低聚糖,從而導致纖維素鏈易水解。接著CX酶作用于非結晶無定型纖維素或可溶性低聚糖,產生纖維二糖,最后在CB酶的水解作用下生成葡萄糖(圖2)。
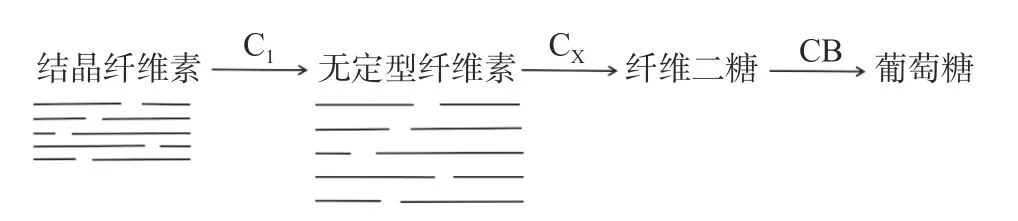
圖2 C1-CX假說理論[48?49]Fig.2 The C1-CX hypothesis theory [48?49]
C1酶可作用于纖維素的結晶區,使之轉變為可被CX酶作用的形式。但該學說沒有很好地解釋C1酶的作用機理。有學者對其提出了種種推測及猜想,如C1酶可能作用于連接纖維素分子間的氫鍵、纖維素分子中的β-1, 4糖苷鍵、其它一些不規則的鍵或薄弱鍵等,但都沒有得到證實[50]。CX酶在水解過程中起著舉足輕重的作用,在一定程度上影響著纖維素大分子水解為纖維二糖的程度,可以用來表征纖維素酶降解纖維素的能力。C1酶和CX酶對纖維素的水解作用是分階段協調的,C1酶只起到轉化纖維素的結晶區但不水解的作用,而CX酶可將部分無定型纖維素水解為可溶性產物。
3.2 協同理論
協同理論是3個理論中接受度最高的理論[51],其降解過程也是由CX酶、C1酶和CB酶3種酶協同完成[52]。其中CX酶的作用是可以給C1酶和CB酶創造酶切位點,再通過水解作用,作用于β-1,4糖苷鍵,長鏈寡糖被裂解為短鏈寡糖,此時一些帶有非還原性末端的纖維素分子就會顯現出來。最后,C1酶作用于這些帶有非還原性末端的小分子纖維素末端,使纖維二糖分子脫下,再在CB酶的水解作用下水解為葡萄糖分子[53](圖3)。Wood[54]發現赤木霉Trichoderma reesei和青霉菌Penicillium funiculosumde產生的纖維素酶在水解微晶纖維素和棉纖維時具有協同性。這種現象可能是由于兩種纖維素酶底物的立體特異性不同所致。在協同過程中,每種纖維素酶作用于兩個不同的非還原性鏈末端中的一個。當一種纖維素酶切斷一種非還原鏈末端的一個纖維二糖單元時,另一種非還原鏈將暴露出來,并且具有不同底物立體特異性的第二種纖維素酶將切斷纖維二糖單元。在這個過程中,這兩種酶的協調可以解釋協同效應。而在晶態幾丁質降解過程中,通過幾丁質降解酶系的內外協同,促進了外切幾丁質酶在幾丁質纖維上“持續奔跑”的耐力,進而增強昆蟲在蛻皮過程中表皮的快速降解[55]。
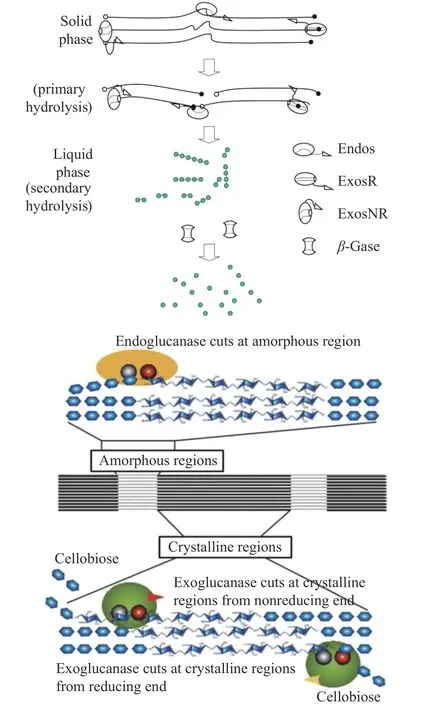
圖3 協同作用降解纖維素示意圖[56?57]Fig.3 Schematic diagram of synergistic cellulose degradation[56?57]
與C1-CX假說不同的是,在協同理論中,纖維素酶的順序沒有固定性和單一性。由于每種纖維素酶組分在水解過程中的確切作用還不清楚,纖維素酶的類型也沒有嚴格的區別。因此,雖然協同理論可以解釋大多數纖維素的降解過程,但對起始反應是如何開始的及天然纖維素結晶區的降解機制尚不能完全闡明[58]。
3.3 原初反應假說
原初反應是一個無序反應,最早是在褐腐菌的水解研究中提出的。褐腐菌菌絲處于靠近次生木質細胞壁的S3層,在降解纖維素過程中菌絲可產生降解劑,擴散到S2層,從而導致擴散性腐爛(圖4)。當褐腐菌開始降解纖維素時,機械強度急劇下降,在此過程中只有少量的重量損失和少量的還原糖生成[59],這與纖維素酶水解過程中產生的聚合度降低而生成的還原糖含量增加的變化明顯不同。因此Coughlan[60]提出原初反應可以改變纖維素的結晶狀態,更有利于纖維素的水解。褐腐菌的獨特之處在于,當木材完全降解時,木質素很少被降解,這有利于纖維素和半纖維素的快速解聚。鑒于褐腐菌在降解天然纖維素方面的獨特性,有學者推測,褐腐菌可能存在一種特殊的非酶木材降解系統,其中一小部分物質可進入S2解聚纖維素[61]。
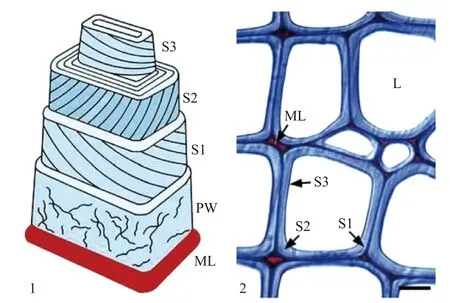
圖4 木質細胞壁模型Fig.4 Wood cell wall model
另外霉菌在降解時首先會進入纖維細胞壁里面,通過CX酶和C1酶的協同作用降解纖維素。而褐腐菌不含C1酶,CX酶的作用可能只打開一小部分的糖苷鍵,能降低纖維素的強度,削弱纖維的表面,所以只能看到機械強度的下降等物理變化,其他變化很小,比如其還原糖含量用還原糖分析方法則無法測出。
4 展望
不同菌種生成的纖維素酶之間可能存在某種程度的拮抗作用,從而影響纖維素降解效率。單一菌株易培養、易獲取,但酶活性較低,降解不徹底。而多種微生物協同作用在降解中表現出的優勢更明顯,降解效果更理想,尤其是復合菌系的環境耐受性和穩定性,對纖維素的降解意義重大。但天然復合菌系和優化組合單一菌株的各方面條件相對嚴格,因此篩選過程中還需注重培養環境的優化。同時,聯合基因工程組學,通過分子生物學技術分離纖維素酶編碼基因,促進基因表達,包括排除對阻遏敏感的操縱基因、提高轉譯效率和將基因連到強啟動子上等,都可有效改善纖維素酶穩定性,達到提高降解率目的[63?64]。
天然纖維素在降解過程中涉及到的能量變化、新陳代謝復雜多變,具體的生物降解過程尚不完全明確。因此,仍需進一步探究纖維素酶類的不同組分和小分子活性物質之間的作用,從酶學、動力學等多個角度綜合揭示纖維素的降解機理。待深入闡明其降解機制過后,就可以實現菌群的人工調控,基于理論分析和應用研究的有機結合,使纖維素酶的水解達到極致,實現纖維素作物的最大限度利用。總之,實現纖維素的微生物降解和資源化利用,是將閑置纖維素類作物變廢為寶、保護環境、緩解世界能源危機、推動綠色可持續發展的重要手段。
