橘小實蠅發生特點及影響因素研究進展*
2022-04-21 00:35:18郭騰達孫瑞紅葉保華宮慶濤姜莉莉武海斌
中國果樹 2022年4期
關鍵詞:影響
郭騰達,孫瑞紅,葉保華,宮慶濤,姜莉莉,武海斌
(1 山東省果樹研究所,泰安271000)(2 山東農業大學)
橘小實蠅[Bactrocera dorsalis(Hendel)]屬雙翅目實蠅科,廣泛分布于熱帶和亞熱帶地區,是世界范圍內危害果蔬的重要害蟲[1]。該蟲繁殖力高,適應能力強,寄主范圍極為廣泛,可為害番石榴、芒果、柑橘、石榴、棗、桃、蘋果、柿等46 科250多種水果和蔬菜,常造成減產甚至絕產,故被列為世界性檢疫害蟲[2]。近年來,隨著我國氣候變暖和果蔬大范圍往來調運,橘小實蠅的分布范圍不斷向北擴展[3],有可能逐漸演變成落葉果樹上的主要害蟲,威脅蘋果、桃、梨等果業的健康發展。為科學防控該蟲,綜述了橘小實蠅的發生特點、影響其發生的因素以及自然天敵等方面的國內外研究進展。
1 橘小實蠅發生特點
1.1 生活習性
橘小實蠅雌成蟲產卵器末端尖銳,可直接在果皮和果肉內產卵,雌蟲更喜歡將卵產于果實縫隙處、傷口處、凹陷處以及軟組織等地方。單雌產卵1 200~1 500 粒,孵化率為80%~90%,產卵期40 d以上[4]。在田間其卵期長短隨季節溫度變化而不同,夏季僅需1 d,春秋季需2~3 d,冬季需7~20 d。幼蟲孵出后通常會在被害果實內生長發育,不轉果為害。幼蟲在1~2 齡時不會彈跳,當幼蟲生長到3齡老熟后鉆出果實,從果實表面彈跳至地面,幼蟲在地面會繼續不斷地彈跳,尋找適合場所入土化蛹,部分老熟幼蟲也可在被害果實內化蛹。幼蟲期發育時長一般為9~23 d,幼蟲老熟后在土表下經1~2 d 預蛹后化蛹,蛹期發育時長一般為8~23 d[5]。
橘小實蠅成蟲全天均可羽化,以8:00—10:00羽化最盛。成蟲雌雄比為1∶0.79,羽化后即取食花蜜、果實或露水補充營養[6]。成蟲具有趨光性,在田間,成蟲取食通常在上午天氣較涼爽期間,而中午或下午一般只是在樹干枝條或是葉叢中活動和停歇,夜晚則停止活動[7]。在室內,成蟲可全天取食,如有光照條件夜間也不停止取食。橘小實蠅在整個成蟲階段需要不斷覓食來維持自身的生長發育和達到性成熟,所需要的食物必須含有糖類和蛋白質等。當成蟲性器官發育成熟后才開始交尾,交尾為重疊式,一般在傍晚至黎明前進行[2],在該階段雌雄成蟲均可多次交尾[8]。雌成蟲產卵時,將產卵管刺入果實內1~2 mm 處產卵,產卵孔直徑0.2~0.3 mm,每個產卵孔產1~24 粒卵[9]。
1.2 發生代數
研究表明,橘小實蠅的發生代數在不同地區不一致,代數多的地區存在世代重疊現象。我國由南向北,橘小實蠅年發生代數逐漸減少[10]。在海南1年發生11~12 代,在北京1 年發生4 代,發生代數與氣候條件和寄主種類有關[11-12]。為此,有人把橘小實蠅在國內分布劃分為4 個層次的適生區,不同適生區的發生代數不同[13]。近年來對橘小實蠅潛在地理分布范圍預測表明,其適生范圍不斷向北擴散,部分地區的適生程度加重[14]。表1 匯總了橘小實蠅在國內部分地區的發生代數、發生時間及發生高峰期,顯示其發生時間與水果生長時間同步,果實成熟前后多是該蟲的發生高峰期,收獲后轉移到下一個近成熟的寄主上取食,故該蟲在果品四季豐富的海南、廣東存在2 個發生高峰,在果品成熟期集中在夏末和秋季的北方發生高峰期為8—10 月。
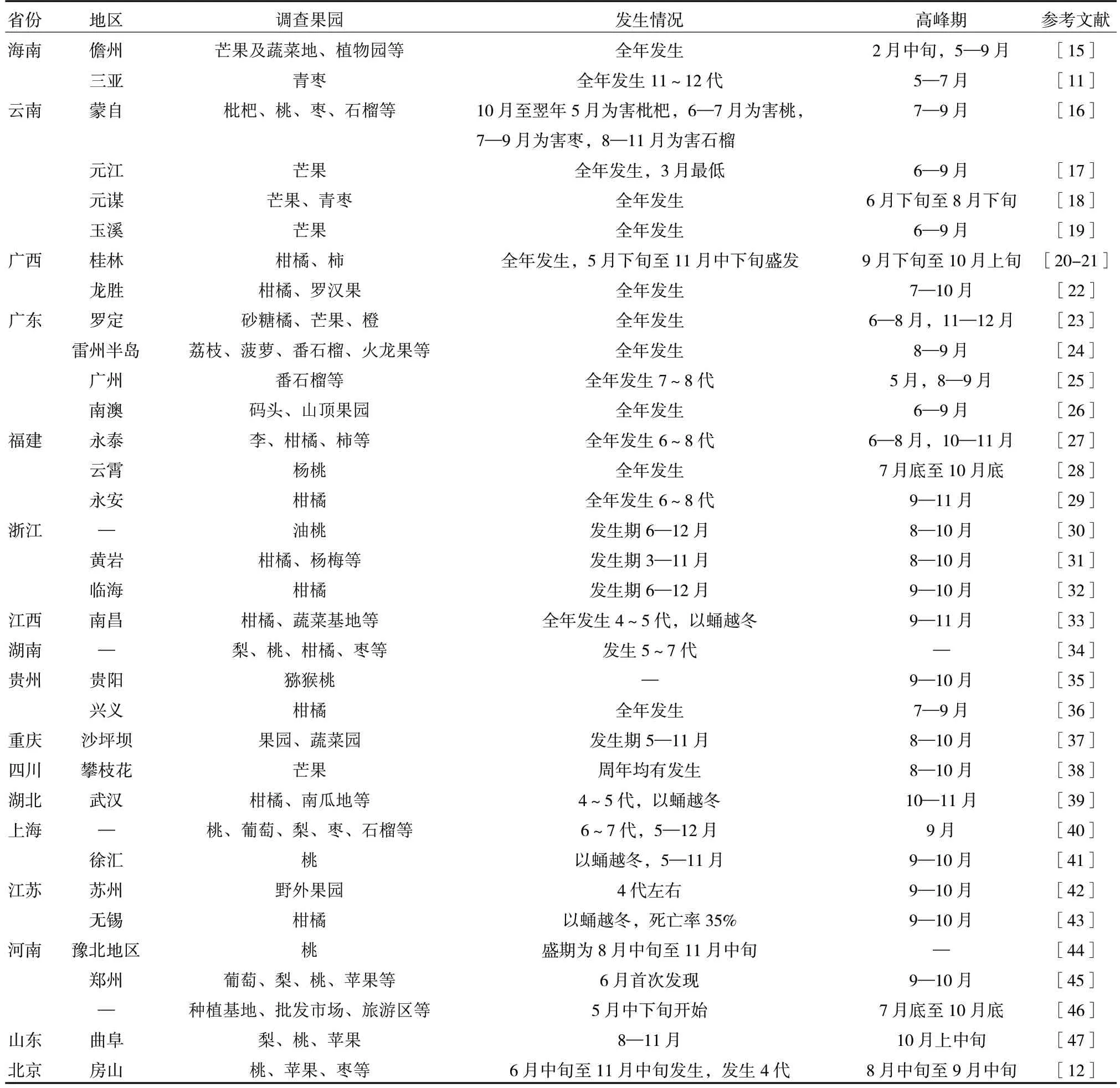
表1 橘小實蠅在國內部分地區的發生情況及發生高峰期
2 影響橘小實蠅發生的因素
2.1 溫濕度的影響
溫度是影響昆蟲生長發育、分布和種群動態的主要因素之一。橘小實蠅的卵、幼蟲和成蟲的生長發育主要受大氣溫度影響,而蛹的生長發育則主要受土壤溫度影響[48]。由于受到該蟲地理種群、食料、試驗條件等因素影響,獲得的發育起點溫度、過冷卻溫度、有效積溫等結果有差異。袁盛勇等利用云南種群進行室內不同溫度試驗,發現橘小實蠅卵、幼蟲、蛹、成蟲的發育起點溫度分別為11.95、11.70、12.83、12.44 ℃,完成對應的各蟲態分別需要有效積溫25.82、175.13、138.12、325.83 d·℃,完成從卵到成蟲的整個生活史的起點溫度和所需的有效積溫分別為10.21 ℃和356.72 d·℃[49]。羅智心等在室內人工飼養入侵無錫地區的橘小實蠅,發現橘小實蠅卵、幼蟲和蛹的發育所需溫度分別為10.5、9.6、10.3 ℃,完成相應蟲態發育所需有效積溫分別為24.1、178.6、176.7 d·℃,完成整個生活史的起點溫度和所需的有效積溫分別為9.9 ℃和381.0 d·℃[50]。因此不同地區的年有效積溫不同會導致橘小實蠅的年發生代數有很大差異,而冬季低溫不同會造成部分地區橘小實蠅不能越冬和完成世代發育[51]。
溫度不僅影響發育速率,同樣影響橘小實蠅各蟲態的存活率與雌雄成蟲性比,在25 ℃時的雌蟲比率最高[50]。由于溫度不同橘小實蠅產卵前期會隨著季節的變化而變化,在夏季一般為10~20 d,溫度越低產卵前期越長,到冬季為3~4 個月。該蟲最適的發育溫度為18~30 ℃;當環境溫度低于18 ℃時,卵、幼蟲和蛹的生長發育時間會延長,蛹的羽化率會下降;當環境溫度高于34 ℃或低于15 ℃時,成蟲和幼蟲的生長發育就會受到影響,甚至會發生大量死亡,且幼蟲不能夠正常化蛹[52]。22~30 ℃是最適宜成蟲飛翔的溫度,在該溫度下雄成蟲飛翔比率為100%,而當環境溫度低于10 ℃時,絕大數雄成蟲會停止飛翔[53]。成蟲和幼蟲的臨界熱最大值分別為46.16、45.23 ℃[54],卵期、幼蟲期、蛹期和卵-蛹期的最適溫度分別為20.7、21.8、21.1、22.4 ℃[55]。
在自然界中,橘小實蠅也會受到濕度的影響。當田間土壤含水量低于40%或高于80%時,老熟幼蟲入土化蛹變慢,死亡率升高[56]。橘小實蠅在溫度恒定為25 ℃的條件下,空氣相對濕度為83%~86%時,成蟲羽化率最高[57]。據Andrei 研究,空氣相對濕度在60%~80%的范圍內最適合橘小實蠅飛翔活動,當空氣相對濕度高于或低于該范圍時不利于橘小實蠅飛翔活動[58];國內研究也發現,在橘小實蠅飛翔活動最活躍時氣溫應處于20~30 ℃,空氣相對濕度應大于60%[7]。在干旱季節,由于空氣相對濕度低,雌蟲產卵量會受到影響,種群增長受到抑制,剛羽化的成蟲從干旱土壤中掙扎出來變得更加困難,且無法充分展翅,導致新羽化的成蟲死亡率極度增加,降水帶來的氣溫與空氣相對濕度的變化對橘小實蠅羽化、飛翔活動有顯著的影響,從各地的田間誘蟲高峰看,基本符合這一規律。干旱的環境還會造成橘小實蠅蛹體的暫時性發育遲緩甚至休眠,一旦遇到雨水充足即可大量羽化[59]。
2.2 氣流的影響
橘小實蠅在室外自然條件下可進行遠距離擴散轉移,其分布和發生區域不斷擴張的主要原因之一在于該蟲的強遷飛能力。在云南怒江河谷地區,橘小實蠅在7 d 的時間最遠遷飛距離可達97 km,表明氣流擴散有助于其遷飛[60]。通過我國北方各地區對橘小實蠅的研究表明,該蟲在北方尚無法進行安全越冬[51],故可推斷其初發蟲源應是從長江以南可越冬地區遷飛而來。天氣系統的變化和大氣環流會影響到昆蟲的遷飛,通常在一個地區內發生大型的天氣過程均會引起該地區部分昆蟲的遷飛。我國所處地區較容易發生熱帶風暴和臺風,據統計,平均每年有7 次臺風登入我國境內,臺風發生后勢必會帶來強風、強降水及使陸地大氣環流出現重大調整,因此必然會影響到昆蟲的遷飛和降落[61]。臺風不僅會影響橘小實蠅的遷飛,也會造成當地橘小實蠅消長動態的變化。臺風登陸時由于風大雨大會導致橘小實蠅誘集數量減少,而當臺風過后的第1 d會出現該蟲活動的高潮,導致誘集量大增[62]。臺風的破壞還會造成果袋破裂和果實傷口,均有利于橘小實蠅產卵和取食,蟲量大幅度提高。
2.3 寄主的影響
昆蟲依靠視覺、觸覺等器官對植物的物理、生理等特征和性狀做出相應的判斷,以檢測其是否適宜作為產卵寄主,依靠嗅覺系統對寄主散發的氣味進行選擇。橘小實蠅的寄主有250 余種果蔬,寄主種類之間顏色、形態、氣味不同會導致它們對橘小實蠅的吸引力有顯著差異。任荔荔等的室內試驗結果表明,橘小實蠅對不同水果的產卵量由多到少依次為:柑橘、香蕉、芒果、棗、石榴、蘋果、桃、梨、李、葡萄、柿,對不同蔬菜的產卵量由多到少依次為:絲瓜、豇豆、苦瓜、黃瓜、茄子、南瓜、青椒、番茄,雌蟲更偏好于橘黃色、綠色以及圓球形的寄主進行產卵[63]。取食不同寄主對橘小實蠅雌成蟲的產卵數量也存在較大的影響,在25 ℃條件下,取食芒果、番石榴、香蕉、枇杷、青棗、楊桃、柑橘的橘小實蠅單頭雌蟲的產卵量分別為1 634.3、1 520.6、1 309.1、1 019.8、983.3、966.3、912.9 粒[64]。蔡子堅等試驗發現,橘小實蠅對黃熟楊桃的產卵量要顯著高于綠熟楊桃,幼蟲數量隨著果實成熟度的提高而增加,原因是黃熟楊桃氣味揮發性物質種類和含量遠高于綠熟楊桃,特別是存在具有引誘橘小實蠅產卵的幾種活性物質[65-66]。以上表明,橘小實蠅在寄主選擇過程中,寄主顏色、形狀、氣味等都在發揮作用。
田間調查結果顯示,在不同寄主之間橘小實蠅可以轉移為害,如在主要種植柑橘的地區,橘小實蠅每年可發生3~5 代,如果柑橘園周圍栽有番石榴、芒果、番木瓜或是梨、桃等其他寄主植物時,則每年發生代數9~10 代[67]。橘小實蠅在上海地區7—8 月主要在桃上為害,8—9 月主要在桃、葡萄、梨、棗上為害,10 月以后主要在柑橘和柿上為害[40]。
2.4 其他因素影響
此外,橘小實蠅發生量還會受到光照、降水、海拔以及天敵等因素的影響。研究發現,中長波光照有利于該蟲的種群增殖,光照強度會影響橘小實蠅的活動能力,其飛行活動的基本條件是要有光刺激,只有在白天有光照的條件下雄成蟲才會飛行,夜晚停止飛行,在光照強度500~2 000 lx 條件下其平均飛行距離、時間最長[68]。由于橘小實蠅老熟幼蟲需要進入土壤化蛹,而降水量與降水頻率會影響到土壤濕度,從而會影響其幼蟲化蛹和成蟲羽化。此外,降水還會影響到空氣濕度,進而影響成蟲的交配產卵和種群數量。如在蘇州地區,6—7 月過多的降水導致土壤過濕板結,從而影響其化蛹、羽化和繁殖,而到8—9 月降水量下降,適宜的溫濕度和土壤疏松度導致其種群數量逐漸上升[42]。同時,海拔也會影響橘小實蠅的發生,在浙江黃巖地區監測結果發現,山區高海拔地區和丘陵地的成蟲誘集數量相對少于平原和沿海低海拔地區[31]。
3 橘小實蠅的天敵種類及其利用
天敵是影響橘小實蠅田間種群數量大小的關鍵因素之一,利用天敵控制害蟲是一種安全有效的手段。橘小實蠅的天敵種類主要包括寄生性天敵和捕食性天敵,但目前關于橘小實蠅的捕食性天敵研究報道較少,且防治效果并不顯著,據報道,螞蟻、雞可以捕食裸露在地表的老熟幼蟲、蛹以及剛羽化的成蟲,隱翅蟲、螨類等可以捕食脫落果實內的幼蟲[69]。
對于橘小實蠅寄生性天敵的研究報道較多,目前發現該蟲的寄生天敵種類有70 余種,其中已知寄生蜂34 種,我國有12 種[70]。橘小實蠅的幼蟲潛藏在果實內部取食,老熟后脫果落入土壤內化蛹,化學防治很難取得較好的效果,因此要對橘小實蠅進行可持續治理,利用寄生蜂是非常有效的手段之一。早在1951 年,美國夏威夷就已經推廣應用阿里山潛蠅繭蜂(Fopius arisanus)、長尾全裂繭蜂(Diachasmimorpha longicaudata)、凡氏費氏繭蜂(F.vandenboschi)和切割潛蠅繭蜂(Psyttalia incisi)防治橘小實蠅,平均寄生率可達70%。我國相關研究起步較晚,1985 年成功引進前裂長管繭蜂,經應用發現它在橘小實蠅種群數量控制上可起到一定作用[69]。目前發現,我國橘小實蠅的寄生蜂分布較廣的主要是長尾全裂繭蜂、凡氏費氏繭蜂和印嚙小蜂(Aceratoneuromyia indica),其中長尾全裂繭蜂、凡氏費氏繭蜂主要分布在福建和廣東省,印嚙小蜂分布于廣東、海南[71]。有研究發現,阿費氏繭蜂(阿里山潛蠅繭)、凡氏費氏繭蜂和長尾全裂繭蜂偏好黃色和白色球體[72],說明橘小實蠅取食果實的顏色和形狀會影響其寄生蜂的選擇和定位。此外,利用切割潛蠅繭蜂也有很好的防治效果,在失管番石榴果園中的寄生率可高達20%。對于不同的水果,切割潛蠅繭蜂的寄生橘小實蠅總量為番石榴>楊桃>蓮霧[73]。
橘小實蠅的齡期、蟲態以及蟲口密度會影響其寄生蜂的發生。在福建漳州,梁光紅等通過對該地區橘小實蠅及其主要寄生蜂的研究發現,寄生蜂種群的發生期和高峰期與橘小實蠅的種群消長表現一致,且有明顯的跟隨效應,在橘小實蠅發生數量達到高峰1~2 個月之后寄生蜂的數量隨即也達到高峰[73]。凡氏費氏繭蜂對1~4 日齡的橘小實蠅幼蟲寄生率明顯高于4 日齡以上的幼蟲[74];長尾全裂繭蜂對3~4 日齡的橘小實蠅幼蟲有較高的寄生率[75]。因此,在釋放寄生蜂時,必須做好橘小實蠅蟲情監測,做到精準釋放、高效生防。另外橘小實蠅的殺蟲劑也會影響其寄生蜂存活和發育,Stark 等研究發現,滅蠅胺、多殺菌素對寄生蜂的安全性高于二嗪膦,在防治橘小實蠅時需要合理使用農藥,協調化學防治與生物防治,以保護寄生蜂[76-77]。
寄生蜂的成功利用離不開自然保護和人工繁殖。目前,一些國家和地區在橘小實蠅的寄生蜂繁育和釋放上做了大量工作,遇到的困難重重,主要表現在室內飼養過程中雌雄蜂比例失調,雄蜂數量遠多于雌蜂,無法快速得到大量可以發揮寄生效果的雌蜂,同時增加生產成本[78]。但是,經過研究者的持續努力,在工廠化繁殖阿里山潛蠅繭蜂方面大獲成功,已在美國、阿根廷、澳大利亞、中國等多個國家使用該寄生蜂防治實蠅害蟲,并取得良好成效[79]。今后寄生蜂作為防治橘小實蠅的重要天敵,應當進一步加強其利用與保護,深入對寄生蜂、橘小實蠅以及寄主植物之間的關系研究,結合性誘、套袋等物理防治,有效地防控橘小實蠅,提高產量,增加經濟收入。在未來其他橘小實蠅天敵的規模化繁育與釋放技術也將會被攻克,生物防治在生產上發揮的作用愈來愈大。
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
家庭影院技術(2020年10期)2020-12-14 07:54:18
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(2016年3期)2016-03-26 07:13:38
知識經濟·中國直銷(2016年3期)2016-02-27 16:15:49
現代檢驗醫學雜志(2014年6期)2014-02-02 03:02:04
閱讀與作文(小學低年級版)(2011年3期)2011-01-01 00:00:00
