基于生物信息學(xué)方法構(gòu)建重度抑郁癥發(fā)病的miRNA-mRNA調(diào)控網(wǎng)絡(luò)
2022-04-20 04:19:34李果張青萍劉永輝吳鵬吳成挺周嬌嬌
山東醫(yī)藥 2022年11期
關(guān)鍵詞:數(shù)據(jù)庫研究
李果,張青萍,劉永輝,吳鵬,吳成挺,周嬌嬌
1 廣西中醫(yī)藥大學(xué)研究生學(xué)院,南寧 530023;2 廣西中醫(yī)藥大學(xué)第一附屬醫(yī)院腦病科
重度抑郁癥(MDD)是一種常見的精神類疾病,日益威脅著人類的健康,但其確切的發(fā)病機制仍未闡明,亦無極其有效的干預(yù)方案[1]。因此,探討MDD的潛在發(fā)病機制、研發(fā)MDD 的有效治療方法是全世界亟需解決的公共衛(wèi)生問題和社會問題。微小RNA(miRNA)是具有調(diào)控功能的非編碼短小RNA,通過調(diào)控下游靶基因參與細(xì)胞的生長發(fā)育過程。研究表明,MDD的發(fā)生發(fā)展與miRNA異常表達(dá)密切相關(guān)[2],但尚未發(fā)現(xiàn)參與MDD 發(fā)生發(fā)展的特異性miRNA。生物信息學(xué)方法是聯(lián)合多學(xué)科領(lǐng)域信息的一種新興技術(shù),通過整合分析疾病相關(guān)數(shù)據(jù),可更高效、系統(tǒng)地揭示疾病的分子機制,并從宏觀層面揭示疾病發(fā)生發(fā)展的復(fù)雜過程。2021年1月—6月,本研究通過生物信息學(xué)方法挖掘MDD 患者外周血中差異表達(dá)的miRNA 和基因,建立潛在的miRNAmRNA 調(diào)控網(wǎng)絡(luò),并篩選出關(guān)鍵miRNA 和基因,為闡明MDD的發(fā)病機制提供理論依據(jù)。
1 材料與方法
1.1 miRNA 微陣列數(shù)據(jù)集選擇 通過國家生物技術(shù)信息中心(NCBI)基因表達(dá)綜合數(shù)據(jù)庫(GEO 數(shù)據(jù)庫),以“major depression”“blood”“miRNA”作為關(guān)鍵詞,搜索出關(guān)于MDD 患者外周血的miRNA 表達(dá)數(shù)據(jù)集,最終篩選出符合條件的數(shù)據(jù)集GSE81152。該數(shù)據(jù)集由GPL21814 平臺合成,包含19 例正常人和30 例MDD 患者的外周血(分別作為對照組和觀察組),正常人均無個人和家族精神病病史,MDD 患者來源于愛爾蘭科克市的精神衛(wèi)生服務(wù)機構(gòu)和圣帕特里克大學(xué)醫(yī)院,均未接受治療。
1.2 差異表達(dá)miRNA(DE-miRNA)篩選 下載GSE81152 數(shù)據(jù)集,并分出對照組和觀察組數(shù)據(jù),采用R軟件的“l(fā)imma”軟件包,對兩組表達(dá)矩陣數(shù)據(jù)進(jìn)行常規(guī)背景校正和分位數(shù)歸一化;通過高級的t檢驗方法比較兩組基因表達(dá)水平,以P<0.05 和|logFC|>0.15作為基本條件篩選DE-miRNA,最后使用R 軟件的“Gplot”軟件包對比較后的結(jié)果進(jìn)行可視化,即構(gòu)建差異分析的火山圖。
1.3 DE-miRNA 上游轉(zhuǎn)錄因子和下游靶基因預(yù)測 使用FunRich 軟件預(yù)測DE-miRNA 上游的潛在轉(zhuǎn)錄因子,并篩選出影響DE-miRNA 表達(dá)的主要上游轉(zhuǎn)錄因子,以P<0.05 作為鑒定轉(zhuǎn)錄因子的闕值。使用miRWalk 和TargetScan 數(shù)據(jù)庫分別預(yù)測上、下調(diào)DE-miRNA 的下游靶基因,再篩選出兩個數(shù)據(jù)庫共同的上調(diào)DE-miRNA 的下游靶基因和下調(diào)DEmiRNA的下游靶基因。
1.4 DE-miRNA 下游目標(biāo)基因篩選 通過GEO 數(shù)據(jù)庫檢索并下載MDD 患者外周血的基因表達(dá)芯片GSE98793 和GSE76826,其中GSE98793 數(shù)據(jù)集包含64 例份健康對照人群和128 例份MDD 患者的全血樣本,GSE76826 數(shù)據(jù)集包含12 例份健康對照人群和10 例份MDD 患者的外周血樣本。采用R 軟件的“l(fā)imma”軟件包分別對下載整理好的兩個數(shù)據(jù)集進(jìn)行校正和歸一化,分別比較兩組外周血相關(guān)基因的表達(dá)水平,并篩選出同時滿足P<0.05 和|logFC|>0.15 的差異表達(dá)基因(DEG),并構(gòu)建火山圖。將上調(diào)DE-miRNA 的下游靶基因和下調(diào)的DEG 進(jìn)行交集,將下調(diào)DE-miRNA 的下游靶基因和上調(diào)的DEG進(jìn)行交集,最終獲得DE-miRNA下游目標(biāo)基因。
1.5 京都基因與基因組百科全書(KEGG)富集分析 通過4個不同的數(shù)據(jù)庫(Enrichr數(shù)據(jù)庫、DAVID數(shù)據(jù)庫、Metascape 數(shù)據(jù)庫和GlueGO 數(shù)據(jù)庫),對DE-miRNA 下游目標(biāo)基因分別進(jìn)行KEGG 富集分析,篩選出主要的信號通路,設(shè)置條件為P<0.05。
1.6 miRNA-mRNA 網(wǎng)絡(luò)構(gòu)建 利用String 數(shù)據(jù)庫構(gòu)建目標(biāo)基因的蛋白質(zhì)—蛋白質(zhì)互作(PPI)網(wǎng)絡(luò),設(shè)置網(wǎng)絡(luò)最低要求互作得分為0.15。通過miRNA 預(yù)測的靶基因結(jié)果找到目標(biāo)基因?qū)?yīng)的miRNA,使用cytoscape3.6.1 軟件構(gòu)建miRNA-mRNA 調(diào)控網(wǎng)絡(luò)圖;最后用插件“cytohubba”進(jìn)行網(wǎng)絡(luò)拓?fù)浞治觯鶕?jù)MCC、MNC 和Degree 三種不同的計算方法篩選出核心miRNA和核心基因。
2 結(jié)果
2.1 DE-miRNA 篩選結(jié)果 在獲得的DE-miRNA中,上調(diào)的DE-miRNA 有8 個(hsa-miR-3129-3p、hsamiR-4428、hsa-miR-3122、hsa-miR-4783-5p、hsa-miR-186-5p、hsa-miR-24-3p、hsa-miR-185-5p、hsa-miR-3156-5p),下調(diào)的DE-miRNA 有10 個(hsa-miR-668、hsa-miR-496、hsa-miR-655、hsa-miR-3664-3p、hsamiR-1208、hsa-miR-129-5p、hsa-miR-214-5p、hsamiR-1275、hsa-miR-539-5p、hsa-miR-1185-2-3p)。見OSID碼圖1、2。
2.2 DE-miRNA 上游轉(zhuǎn)錄因子和下游靶基因預(yù)測結(jié)果 在FunRich 軟件篩選出的DE-miRNA 主要上游轉(zhuǎn)錄因子有5個,分別是特異性蛋白1(Sp1)、特異性蛋白4(Sp4)、早期生長反應(yīng)1(EGR1)、鋅指蛋白143(ZNF143)和鋅指蛋白161(ZFP161),見OSID 碼圖3。miRWalk 和TargetScan 數(shù)據(jù)庫共同的上調(diào)DE-miRNA 和下調(diào)DE-miRNA 的下游靶基因分別有13 351和15 549個。
2.3 DE-miRNA 下游目標(biāo)基因篩選結(jié)果 在GSE98793 數(shù)據(jù)集中上、下調(diào)的DEG 分別有231、241個(OSID 碼圖4A),在GSE76826 數(shù)據(jù)集中上、下調(diào)的DEG分別有3 290、3 377個(OSID碼圖4B)。上調(diào)DE-miRNA 的下游靶基因與下調(diào)DEG 的交集得到47 個基因、下調(diào)DE-miRNA 的下游靶基因與上調(diào)DEG 的交集得到48 個基因,最終共得到95 個下游目標(biāo)基因(OSID碼圖5)。
2.4 下游目標(biāo)基因的KEGG 富集分析結(jié)果 下游目標(biāo)基因在造血細(xì)胞系、Th17細(xì)胞分化、人類T細(xì)胞白血病病毒1感染、mTOR信號通路中顯著富集。見OSID碼圖6。
2.5 miRNA-mRNA 網(wǎng)絡(luò)構(gòu)建結(jié)果 PPI 網(wǎng)絡(luò)的網(wǎng)絡(luò)節(jié)點數(shù)為95、邊數(shù)為205、平均節(jié)點度為4.32、平均局部聚類系數(shù)為0.325。miRNA-mRNA 調(diào)控網(wǎng)絡(luò)圖,見OSID 碼圖7。網(wǎng)絡(luò)拓?fù)浞治龊蠛Y選出3 個核心miRNA(hsa-miR-4428、hsa-miR-3664-3p 和hsamiR-539-5p)和5 個核心基因(IL-7R、IKZF3、CD3D、CD3G和MS4A1)。見表1。
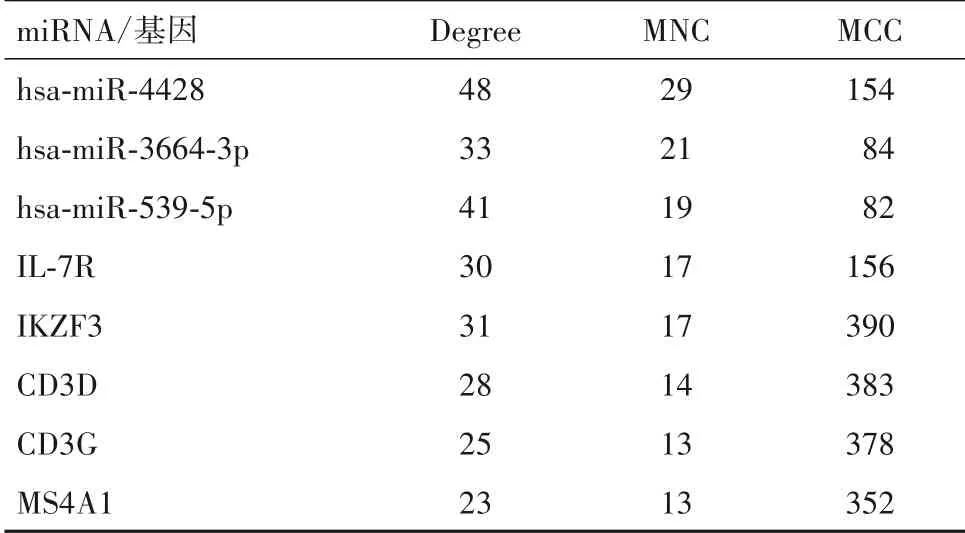
表1 核心miRNA和基因在3種計算方法中的得分(分)
3 討論
研究顯示,miRNA 通過調(diào)節(jié)基因的表達(dá)在神經(jīng)精神疾病中起著重要作用,其失調(diào)導(dǎo)致下游基因表達(dá)水平的改變也是MDD 的發(fā)病原因之一[2-3]。但是目前關(guān)于MDD 患者比較全面的miRNA-mRNA 調(diào)控網(wǎng)絡(luò)相關(guān)研究報道較少。因此本研究先對GEO 數(shù)據(jù)庫中MDD 患者的miRNA 數(shù)據(jù)芯片進(jìn)行差異分析,最終得到8 個上調(diào)的DE-miRNA 和10 個下調(diào)的DE-miRNA,其部分分析結(jié)果也有相關(guān)的研究報道。miR-24-3p在MDD 患者的腦組織(腹側(cè)前額葉皮層)中表達(dá)上調(diào),予抗抑郁治療后血液中miR-24-3p 表達(dá)則明顯下降[4];MDD 患者血清中miR-185 表達(dá)明顯高于健康對照人群[5];另外,miR-185-5p 在雙相情感障礙患者(含有MDD 患者)中的表達(dá)也明顯上調(diào)[6]。有研究表明,miR-185可通過抑制額葉皮質(zhì)中TrkB-T1 的表達(dá),導(dǎo)致精神類疾病發(fā)生[7]。miR-129-5p在MDD小鼠腦內(nèi)合成顯著降低,其表達(dá)改變可能與TRPC3/6 和TREK1 活性變化導(dǎo)致神經(jīng)元離子失衡有關(guān)[8]。雖然目前暫未找到miR-214-5p 在MDD中的相關(guān)研究,但DENG 等[9]通過動物實驗發(fā)現(xiàn)在抑郁小鼠內(nèi)側(cè)前額葉皮層中miR-214-3p 表達(dá)明顯增高,且認(rèn)為該miRNA 是通過抑制了β-catenin 表達(dá)而導(dǎo)致了小鼠的抑郁行為,有效降低miR-214-3p 表達(dá)可達(dá)到治療MDD 的效果。這與本研究篩選的結(jié)果有所不同,這種差異可能與所取的樣本、種類或?qū)嶒炇曳椒ú煌嘘P(guān),后期仍需更多實驗研究來探討。
先前有研究表明,miRNA 的表達(dá)可受轉(zhuǎn)錄因子調(diào)控,并進(jìn)一步共同調(diào)節(jié)下游mRNA的轉(zhuǎn)錄和翻譯,是細(xì)胞代謝的主要調(diào)節(jié)劑[3]。因此本研究利用Fun-Rich 軟件預(yù)測了調(diào)節(jié)DE-miRNA 并可能在MDD 中發(fā)揮作用的轉(zhuǎn)錄因子。Sp1 和Sp4 屬于轉(zhuǎn)錄因子家族的知名成員,均參與多種基本生物學(xué)過程,如細(xì)胞生長、分化、凋亡等,在多種精神疾病中的調(diào)節(jié)作用也已得到充分證明[10],但其在MDD 中的影響尚不明確。EGR1 被定義為鋅指轉(zhuǎn)錄因子,可通過參與大腦中神經(jīng)元活動的關(guān)鍵過程而影響神經(jīng)精神疾病的發(fā)展,包括MDD。COVINGTON 等[11]發(fā)現(xiàn),EGR1 在MDD 患者大腦前額皮質(zhì)中呈低表達(dá),因此上調(diào)EGR1 表達(dá)可能是抗抑郁治療的一種有效手段。但ZHANG 等[12]卻發(fā)現(xiàn),EGR1 在大腦海馬中的表達(dá)是上調(diào)的,且通過抑制EGR1 表達(dá)可快速發(fā)揮抗抑郁效果。這可能是實驗方法及所取樣本不同而導(dǎo)致的,有關(guān)這些的轉(zhuǎn)錄因子在MDD 中的作用尚有爭議,有待于將來進(jìn)一步研究。
本研究通過對DE-miRNA 靶基因與DEG 的整合,共篩選出95 個目標(biāo)基因并進(jìn)行了KEGG 富集分析,其結(jié)果顯示其在造血細(xì)胞系、Th17 細(xì)胞分化、人類T 細(xì)胞白血病病毒1 感染和mTOR 信號通路中顯著富集。造血細(xì)胞系、Th17 細(xì)胞分化和人類T 細(xì)胞白血病病毒1 感染途徑均與體內(nèi)免疫應(yīng)答過程相關(guān),體內(nèi)免疫炎癥反應(yīng)與MDD 有密切聯(lián)系,而神經(jīng)炎癥是導(dǎo)致情緒障礙(包括MDD)的原因之一,且抑制炎癥反應(yīng)在抗抑郁治療中也有不錯的效果,但免疫反應(yīng)與MDD 的復(fù)雜網(wǎng)絡(luò)關(guān)系還有待更深入研究[13-14]。mTOR 信號通路是由含有mTOR 的復(fù)合物引起下游反應(yīng)的一個途徑,因mTOR 可與多種蛋白質(zhì)結(jié)合形成兩種不同的復(fù)合體(mTORC1 和mTORC2),可將mTOR 信號通路分為mTORC1 途徑和mTORC2 途徑。mTORC1 途徑通過控制蛋白質(zhì)的磷酸化調(diào)節(jié)多種生物過程,且mTORC1 通路失調(diào)可涉及到多種神經(jīng)系統(tǒng)疾病,如MDD、精神分裂癥[15]。因此,激活mTORC1 信號引起下游反應(yīng)能夠抑制抑郁樣行為[16]。而mTORC2 途徑則主要調(diào)節(jié)細(xì)胞存活、細(xì)胞代謝和細(xì)胞骨架組織,但其可通過影響下游AKT/GSK3β 途徑導(dǎo)致MDD 的發(fā)生[17]。研究發(fā)現(xiàn),抗抑郁藥可先通過促進(jìn)大腦海馬及前額葉皮層內(nèi)AKT 和ERK1/2 因子的磷酸化,激活MAPK/ERK 和PI3K/AKT途徑而誘導(dǎo)下游的mTOR信號通路,最后介導(dǎo)腦組織中的突觸途徑發(fā)揮抗抑郁的作用[18]。
本研究最后通過構(gòu)建miRNA-mRNA 調(diào)控網(wǎng)絡(luò),篩選出核心miRNA(hsa-miR-4428、hsa-miR-3664-3p和hsa-miR-539-5p)和 核 心 基 因(IL-7R、IKZF3、CD3D、CD3G 和MS4A1)。目前關(guān)于miRNA 的研究越來越多,但miR-4428 和miR-3664-3p 在MDD 中的研究相對甚少。CIUCULETE 等[19]認(rèn)為,miR-3664-5p 表達(dá)改變是導(dǎo)致MDD 發(fā)生的重要因素,而miR-3664-3p是否有相同的作用尚無定論。miRNA(包括miR-539)可通過靶向調(diào)節(jié)HTR2A 基因多態(tài)性導(dǎo)致MDD 的發(fā)生[20]。研究表明,大腦組織中miR-539-3p在精神類疾病的發(fā)生中發(fā)揮重要作用[21]。IL-7R 基因編碼的蛋白質(zhì)是白細(xì)胞介素7(IL-7)的受體,參與體內(nèi)免疫炎癥反應(yīng),而免疫炎癥反應(yīng)與MDD 的發(fā)生密切相關(guān)[13]。另外,IKZF3 表達(dá)對CD4+T 淋巴細(xì)胞中抗炎細(xì)胞因子IL-10 的表達(dá)有影響,CD3D 和CD3G 基因在T 淋巴細(xì)胞的增殖發(fā)育過程中發(fā)揮作用,而MDD 又與IL-10、T 淋巴細(xì)胞比例改變有關(guān)[22-24]。此外,MS4A1(即CD20)在B 淋巴細(xì)胞增殖和分化中具有重要的調(diào)節(jié)作用,可通過影響B(tài) 淋巴細(xì)胞數(shù)量導(dǎo)致MDD 的發(fā)生[25]。這些重要的基因主要在免疫系統(tǒng)中發(fā)揮作用,而免疫反應(yīng)又是導(dǎo)致MDD 的重要途徑,因此針對這些基因通過免疫系統(tǒng)來參與MDD 的發(fā)生可能是研究其機制的相關(guān)切入點。
綜上所述,本研究成功構(gòu)建了MDD 患者的
miRNA-mRNA 調(diào) 控 網(wǎng) 絡(luò),其 中hsa-miR-4428、hsamiR-3664-3p 和hsa-miR-539-5p 等 核 心miRNA 通 過影響IL-7R、IKZF3和CD3D等核心基因表達(dá),并進(jìn)一步調(diào)節(jié)免疫系統(tǒng),在MDD 的發(fā)生發(fā)展中發(fā)揮重要作用。本研究通過生物信息學(xué)方法構(gòu)建了涉及MDD發(fā)病機理的miRNA-mRNA 調(diào)控網(wǎng)絡(luò),有利于更全面地闡明其潛在機制,也為MDD 的診斷及開發(fā)新的治療藥物提供了一個新思路。但是本文是根據(jù)大數(shù)據(jù)預(yù)測出其可能存在的分子機制,部分預(yù)測結(jié)果仍需進(jìn)一步實驗驗證。
猜你喜歡
體育科技文獻(xiàn)通報(2022年3期)2022-05-23 13:46:54
天津外國語大學(xué)學(xué)報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設(shè)計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學(xué)報(2017年2期)2017-07-05 08:13:02
財經(jīng)(2017年15期)2017-07-03 22:40:49
財經(jīng)(2017年2期)2017-03-10 14:35:35
財經(jīng)(2016年15期)2016-06-03 07:38:02
財經(jīng)(2016年3期)2016-03-07 07:44:46
