人巨細胞病毒感染機體細胞免疫學機制的研究進展
2022-04-19 12:28:56韋愛萍宋雅琴彭薇謝政軍
醫學綜述 2022年7期
韋愛萍,宋雅琴,彭薇,謝政軍
(1.遵義醫科大學第五附屬(珠海)醫院血液內科,廣東 珠海 519100; 2.遵義醫科大學珠海校區,廣東 珠海 519000)
人巨細胞病毒(human cytomegalovirus,HCMV)即人皰疹病毒5型,是β皰疹病毒家族的一員。雖然HCMV在人群中的感染率較高,但在健康人群中多表現為潛伏感染,當機體免疫力下降時才會發生再激活引起病毒載量增加,從而播散演變成巨細胞病毒病。其受累人群主要是新生兒、老年人、器官移植及免疫缺陷者。HCMV感染機體往往較隱匿,在早期機體抗HCMV免疫反應過程中自然殺傷細胞(natural killer cell,NK細胞)占優勢,之后機體接觸來自HCMV抗原的持續性刺激導致T細胞活化[1],隨后產生HCMV特異性T細胞[2]。HCMV在機體內多以低病毒載量潛伏狀態存在,但在炎癥等誘因作用下,短期內病毒載量增加,在NK細胞、HCMV特異性T細胞等免疫細胞作用下又會恢復至低病毒載量的潛伏狀態。在這種不定時、間歇性的病毒抗原長期刺激下,HCMV特異性T細胞增殖擴張,從而導致T細胞亞群失衡,最終影響T細胞功能甚至出現T細胞衰竭[3-4]。機體感染HCMV后通過一系列復雜的免疫反應限制和清除HCMV。反之,HCMV通過編碼大量干擾主要效應機制的基因產物來逃避機體免疫攻擊[5]。機體對HCMV的免疫反應包括體液免疫和細胞免疫,細胞免疫的機制更為復雜。現就HCMV感染機體細胞免疫學機制的研究進展予以綜述。
1 NK細胞對HCMV感染的細胞免疫的應答過程
1.1NK細胞在HCMV感染過程中的作用及免疫過程 在固有免疫系統中,NK細胞充當第一道防線。HCMV感染后NK細胞被激活,并在病毒感染早期、T細胞反應產生前以及建立適應性免疫反應后發揮重要作用。NK細胞表面表達多種激活性和抑制性受體,受體均由生殖細胞編碼,通過整合其表面受體所接收的信號啟動機體免疫反應,從而為機體提供病毒感染的防御反應。NK細胞表達人類白細胞抗原(human leukocyte antigen,HLA)Ⅰ類分子的抑制性受體,HCMV通過降低靶細胞的HLAⅠ類分子的表達水平從而逃避T細胞的識別,而HLAⅠ類分子低表達則導致活化信號產生,激活NK細胞,識別和消除病毒感染的靶細胞。NK細胞與樹突狀細胞相互作用并產生一系列細胞因子和趨化因子(如γ干擾素、腫瘤壞死因子-α、巨噬細胞炎性蛋白-1α、巨噬細胞炎性蛋白-1β和正常T細胞表達和分泌的活性調節蛋白)直接參與免疫防御,這些細胞因子和趨化因子可以招募和調節適應性免疫反應,增強靶細胞的HLAⅠ類分子表達[6]。
NK細胞活化包括以下步驟。①接觸:HCMV顆粒或被HCMV感染的靶細胞與NK細胞表面受體結合,觸發下游的轉導信號。②黏附:被認為是NK細胞效應功能的先決條件,并在NK細胞和靶細胞之間形成“免疫突觸”,為釋放細胞毒性顆粒提供一個封閉的微環境。當NK細胞與靶細胞之間的信號傳遞表現為抑制性信號,則NK細胞不被活化;相反,則形成穩定的激活突觸,NK細胞發生活化。③極化:NK細胞與靶細胞發生黏附后,NK細胞分泌性溶酶體(也稱為細胞毒性顆粒)即刻出現極化。磷脂酰肌醇-3-激酶在NK細胞細胞毒性中起關鍵作用,是第二信使形成的關鍵,顆粒極化由磷脂酰肌醇-3-激酶→鳥苷酸轉換因子→Ras相關C3肉毒桿菌毒素底物1→p21激活激酶→促分裂原活化的蛋白激酶激酶→胞外信號調節激酶途徑所觸發[7]。④脫顆粒:依賴于線粒體凋亡抑制因子vMIA-m38.5的表達,vMIA-m38.5是HCMV基因UL37.1編碼HCMV vMIA-UL37.1的同源基因。
1.2HCMV感染狀態下影響NK細胞功能的受體及相關分子通路變化 NK細胞通過其表面激活性和抑制性受體之間傳遞的信號來調節其功能,HLAⅠ類分子是NK細胞受體識別的主要分子家族,NK細胞功能的發揮取決于NK細胞抑制性受體與靶細胞表面HLAⅠ類分子的選擇性結合[8]。HLAⅠ類分子通常與抑制性受體[如Ly49A和CD94/NK細胞凝集素樣受體亞家族C成員1(NK cell lectin-like receptor subfamily C member 1,NKG2A)]結合而抑制NK細胞的功能,抑制性受體通過位于受體胞質結構域的一個或多個免疫受體酪氨酸抑制基序介導其作用。NK細胞表面受體與配體結合后,免疫受體酪氨酸抑制基序激活胞內含SH2結構域蛋白酪氨酸磷酸酶1,導致含SH2結構域蛋白酪氨酸磷酸酶1磷酸化和募集,從而使下游的刺激信號去磷酸化,為NK細胞提供阻斷激活信號的物理環境。
NK細胞利用其表面受體來識別靶細胞,這些NK細胞表面的受體需要靶細胞表面的HLAⅠ類分子的協助才能發揮作用,又稱為HLAⅠ類分子依賴性受體。HLAⅠ類分子依賴性受體有3個不同的家族:C型凝集素樣受體(如CD94/NKG2A或CD94/NKG2C)、白細胞相關免疫球蛋白樣受體1(leukocyte associated immunoglobulin-like receptor-1,LIR-1)和殺傷細胞免疫球蛋白樣受體(killer cell immnoglobulin-like receptor,KIR)[9]。CD94/NKG2受體可以激活激活性受體(如CD94/NKG2C)或抑制性受體(如CD94/NKG2A),但只識別HLA-E。LIR-1可識別所有的HLAⅠ類分子,并具有不同的特異性。而激活性和抑制性KIR則識別所有經典的HLAⅠ類分子(HLA-A、HLA-B和HLA-C),但具有等位基因特異性,在某些情況下以肽選擇的方式識別[10]。這些受體監測HLAⅠ類分子表達的變化,以誘導靶細胞溶解。NK細胞激活性受體與配體結合發揮效應功能,激活NK細胞受體的配體,包括HLAⅠ類分子結合肽、受體誘導的自配體、外源的病毒編碼分子以及免疫復合物(免疫球蛋白Fc段與CD16結合)[11]。
原發性HCMV感染產生強烈的NK細胞反應與激活性受體NKG2C的過表達有關[12]。HCMV感染會觸發NKG2C+NK細胞的擴增,這些細胞同時高表達LIR-1和KIR[13]。NKG2C是C型凝集素樣受體家族成員之一,其與CD94形成異二聚體,能夠與HLA-E分子相互作用。NKG2C表達的NK細胞選擇性擴增依賴于某些細胞因子的存在以及NKG2C和HLA-E與特定肽的相互作用[14]。另外,HCMV提供UL40衍生的肽,其在感染的靶細胞中維持HLA-E的表達,從而通過NKG2A抑制NK細胞活化。以NKG2C為特征的特定NK細胞群中,NKG2CbrightNK細胞被認為是一群HCMV特異性記憶性NK細胞,能夠識別特定的病毒蛋白,且NKG2Cbright細胞是一種長壽命的細胞,在初次感染后數量不斷增加,抗HCMV水平與NKG2C的擴增有直接關系[12]。HCMV能維持NKG2A的配體HLA-E表達,提供一種抑制肽負載的模擬肽[15]。HCMV感染的靶細胞的HLA-E也能被激活性受體CD94/NKG2C所識別,但抑制性受體CD94/NKG2A與HLA-E結合的親和力高,即抑制性受體的負性信號占主導地位,作用結果則傾向于抑制靶細胞HLA-E表達,故NKG2A與NKG2C協同作用可抑制NK細胞反應[16]。
LIR-1是一種抑制性受體,可與經典(HLA-A、HLA-B、HLA-C)和非經典(HLA-G)等HLAⅠ類分子結合。LIR-1可作為病毒復制的生物標志物,反映HCMV病毒載量變化。LIR-1與HCMV UL18結合的親和力遠高于其天然配體HLA-G,表達UL18的HCMV感染細胞能抑制LIR-1+NK細胞,UL18可以增強NK細胞的殺傷力。因此,在HCMV間歇性再激活過程中,UL18上調可誘導LIR-1表達[17]。HCMV感染能增加NK細胞LIR-1的表達,HCMV抗體水平與NK細胞中LIR-1表達呈負相關,HCMV抗體在高表達LIR-1+NK細胞的機體中表達水平降低[18]。
KIR屬于免疫球蛋白樣超家族成員,具有以下特點:①KIR是包含抑制性和激活性受體的配對受體家族;②HCMV感染后抑制性KIR配體的表達被下調;③KIR是高度多態的,許多多態性位于HLAⅠ類分子結合位點外;④許多活化的KIR不與宿主配體結合[19]。NK細胞通過其膜上的KIR識別HLAⅠ類分子,引起胞質免疫受體酪氨酸抑制基序發生磷酸化,募集Src同源區2蛋白酪氨酸磷酸酶1,產生抑制性或激活性信號,從而調節NK細胞和部分T細胞的免疫應答[20]。NK細胞通過KIR等受體的表達,區分健康的“自我”和多種被HCMV感染的靶細胞,進而發揮免疫功能[21]。在NK細胞中有兩種類型的KIR分子(激活分子和抑制分子),長胞質尾的KIR(KIR2DL1、KIR2DL2、KIR2DL3、KIR2DL5、KIR3DL1、KIR3DL2、KIR3DL3)具有抑制淋巴細胞效應功能,而短胞質尾的KIR(KIR2DS1、KIR2DS2、KIR2DS3、KIR2DS4、KIR2DS5、KIR3DS1)通過相關的DNA X激活蛋白12分子激活淋巴細胞,KIR-L由于帶有免疫受體酪氨酸抑制基序的細胞質尾具有受體抑制作用,而KIR-S受體具有短細胞質尾且有跨膜正電荷,從而與帶有免疫受體酪氨酸激活基序的適配器分子(如DNA X激活蛋白12)關聯[22]。一般情況下,KIR2D受體結合HLA-C配體,KIR3D受體結合HLA-A和HLA-B配體。當靶細胞表面表達HLAⅠ類分子,通過NK細胞上KIR抑制性受體與HLAⅠ類分子結合并釋放抑制信號,阻斷NK細胞的殺傷活性,保護靶細胞不被NK細胞消滅,而激活的KIR-S數量對HCMV感染有潛在保護效應,對HCMV的保護程度也隨著KIR-S激活數目的增多而增加[23]。
2 T細胞對HCMV感染的細胞反應的應答過程
2.1HCMV感染后機體T細胞反應的特點及輔助參與細胞 一定數量的HCMV抗原暴露后,單核巨噬細胞、樹突狀細胞及B細胞等抗原呈遞細胞參與啟動機體適應性免疫反應,促使T細胞識別特定的病毒抗原(磷酸化蛋白65、包膜糖蛋白B等),隨后產生HCMV抗原特異性T細胞。T細胞受體通過識別其靶表位產生HCMV表位特異性T細胞,該亞群細胞為單克隆性,從而決定HCMV抗原表位特異性T細胞抗病毒的精確性。HCMV表位特異性T細胞的產生來自幼稚的T細胞受體記憶庫,該記憶庫具有多樣性,可以針對不同的抗原表位刺激提供有效的免疫反應,并維持免疫效應所需的免疫表型。記憶性T細胞再次接受抗原表位刺激后,表位特異性T細胞增殖分化,從而以定量和定性的方式影響隨后的T細胞亞群。T細胞受體的多樣性反映了針對 HCMV抗原表位有效免疫力的強弱。理論上T細胞受體具有多樣性,但胸腺選擇限制了這一多樣性,使相應的抗原特異性前體數量也受到限制,經過一系列細胞免疫過程最終形成針對不同表位的單克隆特異性T細胞。針對HCMV抗原表位特異性T細胞免疫形成過程見圖1。
有數據表明,在首次檢測到HCMV-DNA后,機體對HCMV的早期T細胞反應平均在第7天達到峰值,T細胞反應最初以CD4+T細胞為主,具有輔助性T細胞1細胞因子特征,CD4+T細胞分泌γ干擾素和腫瘤壞死因子-α等細胞因子,CD4+T細胞反應數天后,HCMV患者外周血中可檢出具有HCMV特異性的CD8+T細胞[24]。HCMV特異性CD4+和CD8+T淋巴細胞分別識別HLAⅡ類分子和HLAⅠ類分子所表達的病毒肽,CD8+T細胞能產生高水平效應分子[如白細胞介素(interleukin,IL)-2、γ干擾素、腫瘤壞死因子-α、穿孔素和顆粒酶],CD4+T細胞分泌大量的細胞因子輔助CD8+T細胞對HCMV產生反應[25]。
2.2HCMV感染后CD4+T細胞應答的主要分子變化 CD4+T細胞是HCMV感染的適應性免疫反應的重要效應因子,在機體初次感染后,HCMV特異性CD4+T細胞群持續存在于機體的T細胞庫中,以控制病毒在宿主生命周期內的再激活。CD4+T細胞在抗病毒免疫應答中的作用大致可分為3類:淋巴細胞向感染部位募集;介導其他效應細胞的擴張;通過細胞因子的產生或細胞介導的細胞毒性發揮抗病毒作用。T細胞表達CD28和CD40配體等共刺激分子,這些共刺激分子聚集是引起CD4+T細胞胞內信號傳遞和隨后激活所必需[26]。
HCMV特異性CD4+T細胞的表型在感染的不同階段具有不同的表型特征,在HCMV感染高峰期,HCMV特異性CD4+T細胞表型為CD45RA/CD45RO雙陽性細胞,表達CD27、CD28、CD38和CD40配體。而潛伏期,HCMV特異性CD4+T細胞表型為CD27/CD28雙陰性細胞。HCMV特異性CD4+T細胞在感染高峰期和潛伏期均能產生經典的輔助性T細胞1細胞因子(如IL-2、γ干擾素和腫瘤壞死因子-α)。Roquin是一種細胞因子,在穩定狀態下控制各種抗病毒基因的表達,以調節免疫穩態,其功能在感染早期和晚期保持不變。Roquin是HCMV誘導免疫調節的關鍵調節因子,在HCMV感染過程中被誘導表達,HCMV積極利用Roquin進行高效的病毒合成。干擾素調節因子1是一種轉錄因子,主要通過誘導促炎細胞因子和促進抗病毒免疫反應來調節免疫應答,而Roquin抑制干擾素調節因子1的表達則可促進HCMV的高效復制[27]。如能尋找到抑制這兩個分子的靶點可能抑制HCMV復制,從而達到治療目的。HCMV可編碼免疫抑制IL-10同源基因,IL-10是輔助性T細胞1細胞因子的一種強有力的抑制劑,也能抑制單核細胞產生炎癥細胞因子,從而導致表面HLAⅡ類分子表達減少,并降低CD4+T細胞抗原的表達。IL-10能明顯抑制MCMV感染慢性期T細胞的增殖[28]。

注:TCR為T細胞受體,HCMV為人巨細胞病毒,pp為磷酸化蛋白,IE1為立刻早期蛋白1,gB為包膜糖蛋白B
2.3HCMV感染后CD8+T細胞應答的主要分子變化 HCMV已被證明可誘導抗原特異性CD8+T細胞克隆擴增,HCMV感染能誘導宿主產生大量效應記憶CD8+T細胞[29]。CD8+T細胞能限制HCMV復制,但不能消除或抑制其傳播。HCMV多輪抗原刺激導致機體HCMV特異性CD8+T細胞的克隆性擴增,隨著年齡的增長HCMV特異性CD8+T細胞不斷累積,增加的CD8+T細胞數量能有效控制HCMV潛伏感染再激活。因此,在活躍的免疫反應中,CD8+T細胞快速克隆擴增可作為免疫介導疾病持續感染的標志。T細胞受體識別細胞表面HLAⅠ類分子所表達的同源抗原肽,介導HCMV表位特異性CD8+T細胞的產生。HCMV特異性CD8+T細胞通過釋放γ干擾素和顆粒酶(中性粒細胞、生長因子受體結合蛋白、G蛋白偶聯受體激酶、代謝型谷氨酸受體)來控制HCMV感染,這些細胞因子水平在受刺激后數小時內升高[30]。HCMV特異性CD8+T細胞可產生趨化因子,將免疫細胞(如CC趨化因子配體4、CC趨化因子配體5)吸引到感染部位,上述免疫細胞可趨化NK細胞、單核細胞以及活化的T細胞。因此,HCMV特異性CD8+T細胞是具有強大效應功能的多功能細胞,能誘導和維持免疫反應。已有多項臨床試驗證實HCMV特異性T細胞的過繼細胞免疫治療具有抗HCMV作用(表1),HCMV特異性T細胞過繼細胞免疫治療在造血干細胞移植后100 d內防治巨細胞病毒病方面具有良好的應用前景;HCMV特異性T細胞在實體器官移植受者中被證明具有保護作用[37];HCMV特異性T細胞治療巨細胞病毒病有一定的療效,如HCMV視網膜炎[38];HCMV特異性T細胞在預防先天性 HCMV感染中也起到一定作用[39]。但HCMV特異性T細胞過繼細胞免疫治療存在持續時間短、效應功能發揮有限等局限性,仍需進一步研究。
2.4HCMV感染后CD4/CD8相互關系 HCMV特異性免疫應答與CD4/CD8密切相關,HCMV感染研究中CD4/CD8發生倒置,CD8+T細胞擴張性增多[40]。CD4+T細胞作為輔助細胞,在抗病毒反應中通過CC趨化因子配體3和CC趨化因子配體4促進CD8+T細胞與樹突狀細胞結合,從而促進CD8+T細胞進入感染部位。CD4+T細胞還能促進幼稚的CD8+T細胞進入引流淋巴結,并通過產生γ干擾素和分泌局部趨化因子,將固有性或抗原特異性的效應物導入病毒復制部位[41]。HCMV特異性CD4+T細胞的存在是維持HCMV特異性CD8+T細胞所必需。CD4+T細胞通過多種機制促進記憶性CD8+T細胞的發育,如下調腫瘤壞死因子相關凋亡誘導配體的表達,產生IL-2等細胞因子[26,42]。但目前這兩種T細胞亞群的失衡及其在抗HCMV過程中的關鍵分子事件尚不清楚。
3 HCMV免疫逃逸及免疫衰老
3.1HCMV免疫逃逸特點及主要分子事件 HCMV是一種對病原免疫逃逸的范式,針對復制的不同步驟而產生抗病毒作用。首先,HCMV編碼細胞因子模擬物和誘餌受體,以調節細胞因子活性,限制細胞遷移和分化。其次,HCMV通過阻斷轉錄因子的激活來抑制細胞因子的產生。第三,HCMV微RNA直接抑制機體信使RNA的轉錄,抑制趨化因子和細胞因子表達。
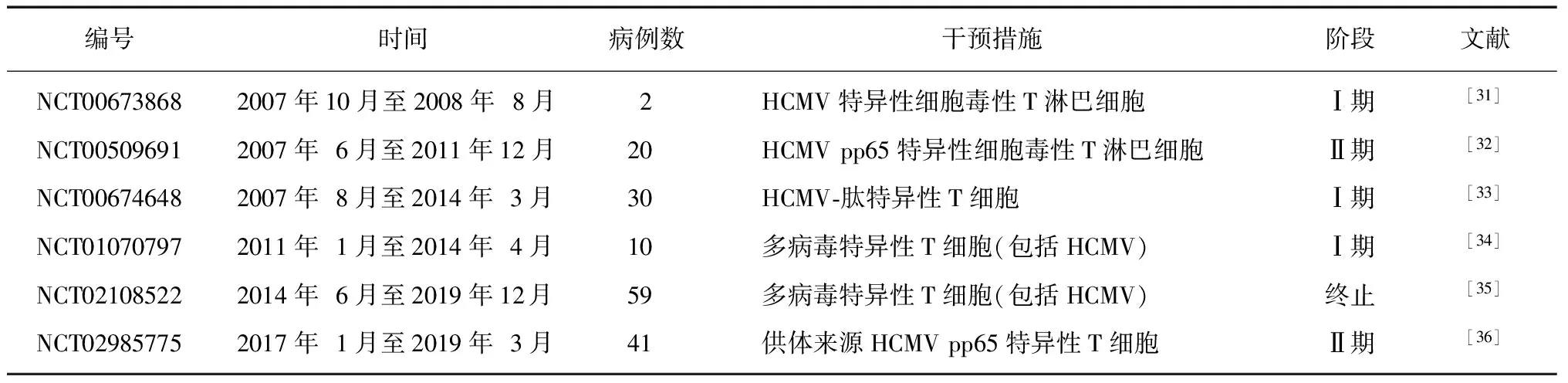
表1 HCMV特異性T細胞過繼細胞免疫治療HCMV血癥和巨細胞病毒病的臨床試驗
HCMV基因組在感染過程中編碼多種逃逸蛋白,使病毒能夠調節固有性和適應性免疫反應,最終即使在免疫能力強的宿主中病毒血癥也會持續存在,病毒排泄可長達數月(健康成人)甚至數年[27]。HCMV有多種方法來避免適應性免疫反應,其中一種方法是通過HCMV特異性表位抗原pp71下調靶細胞HLAⅠ類分子的表達,促進HCMV增殖復制。pp71是由UL82基因編碼的磷酸化蛋白,其表觀分子量為71 000。HCMV pp71蛋白被包裝在病毒的被膜內,阻止轉錄抑制性異染色質在病毒基因組上的組裝,促進病毒基因的轉錄翻譯,通過抑制固有性和適應性免疫反應的成分來幫助免疫逃逸[43]。
3.2HCMV致免疫衰老的免疫細胞改變 與其他慢性病毒感染相比,HCMV感染可引起一種不典型的CD8+T細胞過度增殖,稱為記憶膨脹,而導致CD8+T細胞膨脹的主要原因之一是重復或持續的抗原暴露刺激。在HCMV感染過程中,T細胞識別病毒表位并經歷增殖性擴張,主要以HCMV特異性CD8+T細胞數量增多為主,這種現象在老年人中更為突出,大部分HCMV特異性CD8+T細胞為CD45RA+效應記憶細胞。CD45RA+效應記憶細胞群隨著年齡的增長而逐漸擴大,在老年人群中占比達總效應記憶CD8+T細胞群的一半以上[28]。持續抗原刺激,如潛伏或持續的HCMV感染,被認為是CD45RA+效應記憶T細胞擴展的主要原因。T細胞受體直接識別HLAⅠ類分子呈遞的病毒肽,介導HCMV特異性T細胞的產生限制和清除HCMV[19]。在受感染的細胞中,源自蛋白酶體加工病毒蛋白的抗原肽從胞質溶膠運輸至內質網,加載到HLAⅠ類分子上并通過分泌途徑輸出至細胞表面,HCMV干擾HLAⅠ類分子抗原呈遞途徑的幾乎所有階段,從而避免CD8+T細胞的識別。
3.3HCMV引起免疫衰老主要分子事件 HCMV免疫反應的一個特征是效應記憶表型CD8+T細胞的積累(即CD27-CD28-/CD45RA+),其他特征包括幼稚CD8+T細胞的頻率和數量下降(即CD27+CD28+/CD45RA+)和炎癥活性增強(如IL-6水平)[44]。T細胞的主要共刺激受體是CD28,然而終末分化的CD8+T細胞失去表達CD28的能力。相反,抗原驅動的T細胞活化后,其他共刺激受體(如4-1BB、OX40)在T細胞中表達,并導致小鼠巨細胞病毒特異性T細胞膨脹。老年人的HCMV特異性CD8+T細胞群的這種特殊性質與T細胞的衰老有關。
在慢性感染情況下,NK細胞和CD8+T細胞暴露于持久性抗原和(或)炎癥信號,這種過多的信號常導致宿主出現慢性免疫衰竭狀態[45]。免疫衰竭主要表現為效應功能(細胞因子產生和殺傷功能)的逐漸喪失、多種抑制受體(如程序性細胞死亡受體1和淋巴細胞活化基因-3)的表達、代謝失調和穩態增殖。NK細胞在HCMV感染的早期免疫反應中必不可少,隨后HCMV進化出多種機制來逃避NK細胞介導的病毒清除,通過下調細胞毒性功能或引發細胞凋亡,導致NK細胞耗竭。研究顯示,在慢性淋巴細胞性脈絡膜腦炎病毒感染過程中,程序性細胞死亡受體1在功能耗竭的病毒特異性CD8+T細胞中表達上調,阻斷程序性細胞死亡受體1通路可促進病毒特異性T細胞的效應功能,顯著降低實驗動物的病毒載量[30]。但程序性細胞死亡受體1在HCMV感染狀態下能否促進HCMV特異性T細胞擴增發揮抗病毒效應及發生機制尚不清楚,尚待進一步闡明。
4 小 結
HCMV與宿主免疫系統之間的相互作用是一個復雜的過程,從病毒黏附進入宿主靶細胞開始,隨后引起機體的細胞免疫應答,最終形成一種特異性的適應性免疫反應。NK細胞在HCMV感染早期和潛伏期對病毒的控制起重要作用,通過產生細胞因子攻擊并清除HCMV感染的細胞,并在適應性免疫過程中與T細胞共同作用發揮抗病毒作用,經過一系列適應性細胞免疫反應過程產生HCMV特異性T細胞。隨著感染時間的推移,HCMV通過改變T細胞亞群比例、受體與配體結合、關鍵信號轉導通路的阻斷等方式產生免疫逃逸,這種狀況隨年齡增長表現愈發明顯甚至導致免疫衰竭。因此,在HCMV感染宿主的過程中,宿主抵御病毒的免疫機制復雜多樣,病毒免疫逃逸的策略也復雜多變,仍有很多機制尚未闡明。深入了解HCMV致病的細胞免疫機制、免疫逃逸機制及其對機體免疫衰老的影響,可為HCMV感染的防控提供新思路。
