轉錄組和iTRAQ技術聯合揭示玉米根系的耐旱機制
2022-03-26 07:39:22白明興莊澤龍姬祥卓彭云玲
干旱地區農業研究 2022年2期
白明興,莊澤龍,姬祥卓,彭云玲
(甘肅農業大學農學院,甘肅省干旱生境作物學重點實驗室/甘肅省作物遺傳改良與種質創新重點實驗室, 甘肅 蘭州 730070)
干旱作為全球最重要的非生物脅迫之一,極大地影響了作物的生長發育和產量。玉米作為我國主要的糧飼兼用作物,是典型的干旱敏感作物,各生育期需水量較大[1-3]。研究表明,干旱缺水不僅嚴重影響玉米的生長發育,造成玉米大幅減產,而且會阻礙玉米葉片對CO2的吸收,進而影響其光合性能[4-5]。此外,干旱脅迫會導致植物體內活性氧(ROS)不斷積累,ROS在植物生長發育中有著十分重要的作用,正常條件下主要參與調節植物的生長發育,維持動態平衡狀態[6];當遭受非生物脅迫時,植物體內的ROS快速積累,多余的ROS不能及時清除,動態平衡遭到破壞,從而影響植物的發育[7]。活性氧是氧分子沒有完全還原的產物,其毒性很強,在非生物脅迫下,它不僅可以破壞細胞的結構和功能,而且是信號轉導的重要調節因子[8]。由于ROS具有極高的氧化活性,植物體內蛋白質、磷脂和核酸很容易被氧化,造成細胞中蛋白質降解和核酸裂解,以及磷脂過氧化產生丙二醛,破壞細胞膜結構[9]。且大量研究也表明,ROS 作為信號分子在植物細胞防御響應和細胞程序性死亡等方面扮演著極其重要的角色[10]。當植物遭受輕度干旱脅迫時,會導致植物體內活性氧清除啟動一系列復雜的清除反應[11],促使超氧化物歧化酶(SOD)、過氧化氫酶(CAT)和過氧化物酶(POD)等酶活力提高,保護植物免受環境脅迫的危害。根系作為植物吸收和運輸土壤中水分及養料的主要器官,在遭遇非生物脅迫時,根系一般最先感受到逆境脅迫,從而改變根系的形態以及分布以適應不利環境。因此,深入剖析玉米苗期根系對干旱脅迫的分子基礎及響應機制,對改善干旱和半干旱地區作物的早期生長狀況及促進全球糧食生產安全有著極其重要的作用[12]。
近年來,隨著全基因組測序的快速發展,不同學者在研究植物抗逆性方面也逐漸采用轉錄組和蛋白質組學技術來揭示逆境脅迫中植物的生理、生化和分析響應機制,從而挖掘大量與抗逆相關的基因。ZENDA等[13]為了揭示不同玉米自交系對干旱脅迫的耐受性,采用耐旱自交系YE8112和干旱敏感自交系MO17進行比較轉錄組分析,通過分析獲得了一些耐旱基因和調控干旱脅迫的主要途徑,闡述了不同玉米自交系對干旱脅迫的響應機制,并依據目前研究構建了玉米幼苗抗旱的分子模型。LIU等[14]通過對兩個雜交品種(耐旱ND476和旱敏ZX978)在干旱脅迫下進行轉錄組分析發現,玉米對干旱脅迫的適應是一種品種特異性的反應,也是一種階段性的反應過程,并且該研究也挖掘了一些調控玉米干旱脅迫反應的品種特異性基因和生育期特異性基因。CHEN等[15]通過對兩份耐鹽性不同的玉米自交系根系進行蛋白質組學分析發現,當遭遇鹽脅迫時,不同耐鹽性自交系差異表達蛋白(DEPs)間雖存在明顯差異,但是不同自交系間對鹽脅迫的響應也存在一些相同的趨勢。JIANG等[16]利用iTRAQ(Isobaric tags for relative and absolute quantification)技術發現玉米幼苗在干旱脅迫下存在多重轉錄后調控和翻譯后修飾,并提出在干旱脅迫下玉米幼苗會通過提高其抗旱性來應對干旱的策略。ZHENG等[17]采用耐旱材料28M和旱敏感材料753F從基因調控水平揭示了287M抗旱性的機制,表明287M和753F的抗旱性差異可能與清除ROS能力、信號相互作用網絡和某些轉錄因子有關。HAO等[18]利用RNA測序技術對耐旱性不同的兩自交系根系干旱相關基因進行全基因組鑒定和比較分析,并表明玉米的抗旱性與ABA信號通路、海藻糖生物合成、活性氧清除和轉錄因子相關基因的表達模式有關。以上研究試圖從某一組學水平去解析一些生物學現象,不能深入剖析其響應機理。例如,轉錄組只能關注一些關鍵編碼轉錄本的表達量變化,而不能了解其轉錄調控以及對蛋白翻譯的影響。因此,利用多組學聯合分析剖析一些生物學問題十分必要。轉錄組和蛋白質組學可以從兩個不同層面反映基因的表達情況,所以要了解轉錄組與蛋白質組的相互調控作用,需要對mRNA 與蛋白質的表達進行同步監測。關聯分析的目的之一是實現數據互補,得到生物體更加完整的表達信息。
因此,為了進一步剖析玉米幼苗對干旱脅迫的響應機制,本研究以耐旱自交系昌7-2和干旱敏感自交系TS141的苗期根系為試驗材料,結合轉錄組測序和iTRAQ技術探究不同玉米自交系對干旱的響應機制并篩選一些耐旱候選基因,為進一步了解玉米耐旱的分子機制提供新的思路。
1 材料與方法
1.1 材料處理
用耐旱自交系昌7-2(Chang 7-2)和干旱敏感自交系TS141為試材,對兩自交系分別進行蒸餾水(CK)和20%PEG(D)處理。挑選整齊、一致的種子用0.5%NaClO表面滅菌10 min并用無菌蒸餾水洗滌5次。將種子浸泡在處理溶液中24 h后點播在營養缽(15 cm×13 cm)中,消毒蛭石(溶液∶蛭石=1∶5 v·m-1)用與各處理相同的處理溶液攪拌均勻,每缽10粒,試驗設置3次重復。將材料置于人工氣候箱,晝夜溫度分別為25℃、20℃,光照和黑暗時間分別為16、8 h,光照強度為600 μmol·s-1·m-2。對相應材料每隔1 d加入50 mL蒸餾水,每3 d施加50 mL對應處理液,處理12 d后收集根系樣品進行轉錄組測序和蛋白質組學測序。
1.2 轉錄組測序和蛋白質組測序分析
轉錄組測序參照CHEN等[19]的方法,對兩不同玉米自交系處理后12 d的幼苗根系進行總RNA提取,并進行轉錄組測序。測序所獲得的原始讀段(Raw reads)進行過濾,得到干凈讀段(Clean reads),后續分析都基于Clean reads,并采用HISAT和RSEM軟件以第三版B73玉米參考基因組對Clean reads進行比對(Version ZmB73_5a.59)(http://ftp.maizesequence.org/)和基因表達水平檢測。通過RPKM計算12個樣品的基因表達(每百萬Kbp讀數)方法,其測序的數據已提交到NCBI的SRA(登錄號:PRJNA698088)。為了提高DEG數據的準確性,差異基因篩選標準采用|log2FC(Fold change)|≥1且FDR≤0.001。而蛋白質組測序的具體方法及相關數據來自ZENG等[20]。采用WEGO軟件[21]對篩選出的DEGs在GO數據庫中的功能注釋進行分類。通路顯著性富集分析基于KEGG(Kyoto encyclopedia of genes and genomes)公共數據庫(http:/iwww.genome.jp/kegg/pathway.html)進行,對顯著富集的通路進行鑒定。其中把Q value≤0.05的Pathway定義為在DEGs中顯著富集的Pathway。
1.3 關聯分析
干旱脅迫下,利用LAN等[22]的方法計算了昌7-2和TS141蛋白質水平與轉錄水平的Pearson相關系數,確定蛋白質與mRNA的相關性,并對每個品種關聯了兩個生物學重復,其中Chang 7-2 D1vsCK1比較組表示耐旱自交系昌7-2轉錄組和蛋白組第一個生物學重復所關聯的結果,Chang 7-2 D2vsCK2比較組表示耐旱自交系昌7-2轉錄組和蛋白組第二個生物學重復所關聯的結果;而TS141 D1vsCK1和TS141 D2vsCK2比較組分別表示旱敏感自交系TS141中兩個重復的關聯結果。
1.4 qRT-PCR(Quantitative real time-PCR)驗證
采用RNA提取試劑盒獲取兩自交系不同處理下幼苗根系的總RNA,隨后利用FastKing RT Kit(天根生物技術,北京,中國)對提取的總RNA反轉錄為cDNA,備用。利用NCBI(https://www.ncbi.nlm.nih.gov/)官網進行引物設計,本試驗根據不同基因的表達量共設計了9個基因的特異性PCR引物對轉錄組數據和蛋白數據進行驗證,實時熒光定量(qRT-PCR)分析的具體方法參考CHEN等[19],所選基因的相對表達水平通過2-ΔΔCT方法計算,并以Actin基因的表達水平進行歸一化,每個處理使用3個生物重復。
2 結果與分析
2.1 轉錄組和蛋白質組數據分析
2.1.1 轉錄組測序分析 采用BGISEQ-500測序平臺對兩份玉米自交系根系樣品進行RNA-Seq測序。結果表明測序樣品平均產生116025890.2條Raw reads,經過剔除低質量的Reads后, 每個樣品平均得到107905100.5條高質量的Clean reads。采用HISAT軟件將獲得的Clean reads與玉米參考基因組B73(http://ftp.maizesequence.org/)進行比對,結果表明不同樣品在參考基因組中的平均比對率約68.77%。此外,以|log2Ratio(FC, Fold Change)|≥1,Pvalue≤0.001作為兩樣品間差異表達基因(DEGs)的篩選條件對不同樣品間進行差異表達基因篩選,結果表明,在昌7-2中鑒定出7483個DEGs,其中3162個DEGs上調表達,4321個DEGs下調表達;在TS141中鑒定出6829個DEGs,其中3290個DEGs上調表達,3539個DEGs下調表達(圖1)。
2.1.2 蛋白質組數據分析 采用iTRAQ技術對抗旱性不同的兩自交系幼苗根系進行蛋白質組學分析,結果共產生了97374張光譜、39371個肽以及30148個獨特的肽,最終鑒定到7723個蛋白質。當蛋白質豐度比達到0.83倍以下或1.2倍以上,且P<0.05時,認為該蛋白是兩個不同樣品間的差異表達蛋白(DEPs)。結果表明,在Chang 7-2-DvsCK中鑒定出1243個DEPs,其中572個DEPs上調表達,671個DEPs下調表達;TS141-DvsCK中鑒定出419個DEPs,其中172個DEPs上調表達,247個DEPs下調表達(圖1)。

圖1 干旱處理下兩玉米自交系的DEGs和DEPs
2.2 轉錄組和蛋白質組學關聯性分析
2.2.1 轉錄組和蛋白質組差異表達基因與差異表達蛋白的關聯分析 對轉錄組和蛋白質組數據進行關聯分析,當某一個蛋白質在轉錄組水平有表達量時,被認為關聯到。關聯結果表明(表1),在鑒定、定量和顯著差異表達3個方面,Chang 7-2 D1vsCK1中7612個基因在mRNA和蛋白質水平被同時鑒定,并定量到7609個基因,有157個DEGs 同DEPs關聯。Chang 7-2 D2vsCK2中7621個基因在mRNA和蛋白質水平被同時鑒定,并定量到7619個基因,有98個DEGs 同DEPs關聯。TS141 D1vsCK1中7609個基因在mRNA和蛋白質水平被同時鑒定,并定量到7606個基因,有76個DEGs 同DEPs關聯。TS141 D2vsCK2中7619個基因在mRNA和蛋白質水平被同時鑒定,并定量到7616個基因,有116個DEGs 同DEPs關聯。

表1 鑒定、定量和顯著差異中被關聯的基因和蛋白質數量
2.2.2 轉錄組和蛋白質組相關性分析 基于mRNA水平和蛋白質水平的表達結果,使關聯到的基因在mRNA表達水平和蛋白質水平上存在多種類型,即mRNA和蛋白表達趨勢相同、mRNA和蛋白表達趨勢相反、mRNA表達有差異而蛋白表達無差異、mRNA表達無差異而蛋白表達有差異以及mRNA和蛋白表達均無差異5種關聯類型。本研究發現,Chang 7-2 D1vsCK1、Chang 7-2 D2vsCK2、TS141 D1vsCK1和TS141 D2vsCK2等4個比較組中所關聯到DEGs和DEPs的相關性系數分別為0.3063、0.2208、0.2295和-0.0203(圖2)。其中,mRNA和蛋白表達趨勢相同的基因分別為98(重復基因2個)、64、55(重復基因2個)、62個(重復基因1個)(表2),去除組內重復基因后分別為96、64、53、61;而mRNA和蛋白表達趨勢相反的基因分別為59(重復基因1個)、34(重復基因2個)、21(重復基因1個)、54個(表2), 去除組內重復基因后分別為58、32、20、54。

圖2 顯著差異蛋白質和顯著差異基因表達關聯圖
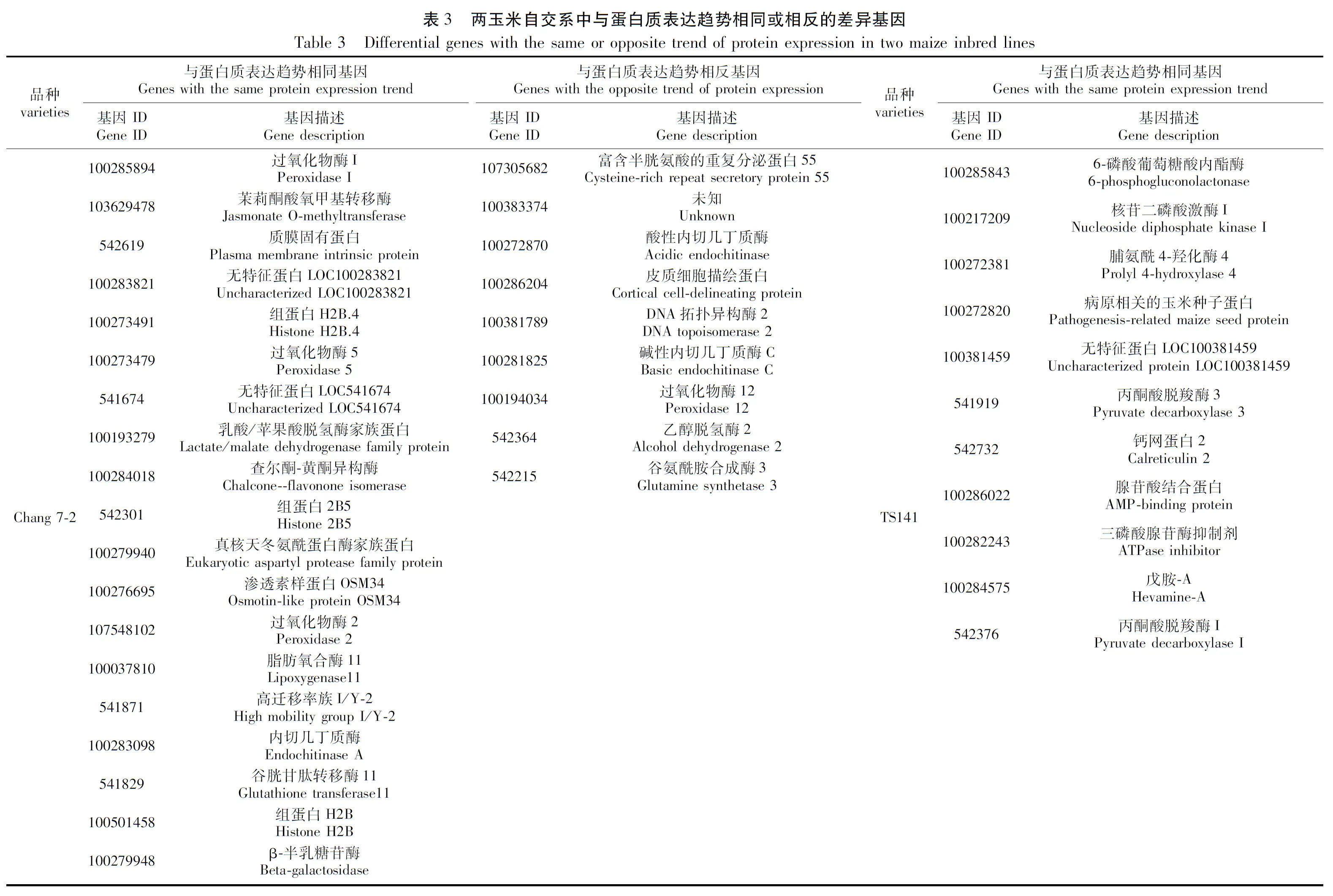
為了進一步剖析兩自交系在干旱脅迫下基因表達的調控情況,對每個自交系D1vsCK1和D2vsCK2的關聯結果進行了整合,以獲得在干旱脅迫下穩定表達的基因信息。結果表明,Chang 7-2的D1vsCK1和D2vsCK2樣品中mRNA和蛋白表達趨勢相同的基因有19個(表3;圖3A),mRNA和蛋白表達趨勢相反的基因有9個(表3;圖3B);TS141 D1vsCK1和D2vsCK2中mRNA和蛋白表達趨勢相同的基因有11個(表3),mRNA和蛋白表達趨勢相反的基因在兩次重復中沒檢索到較為穩定表達的基因(圖3B)。
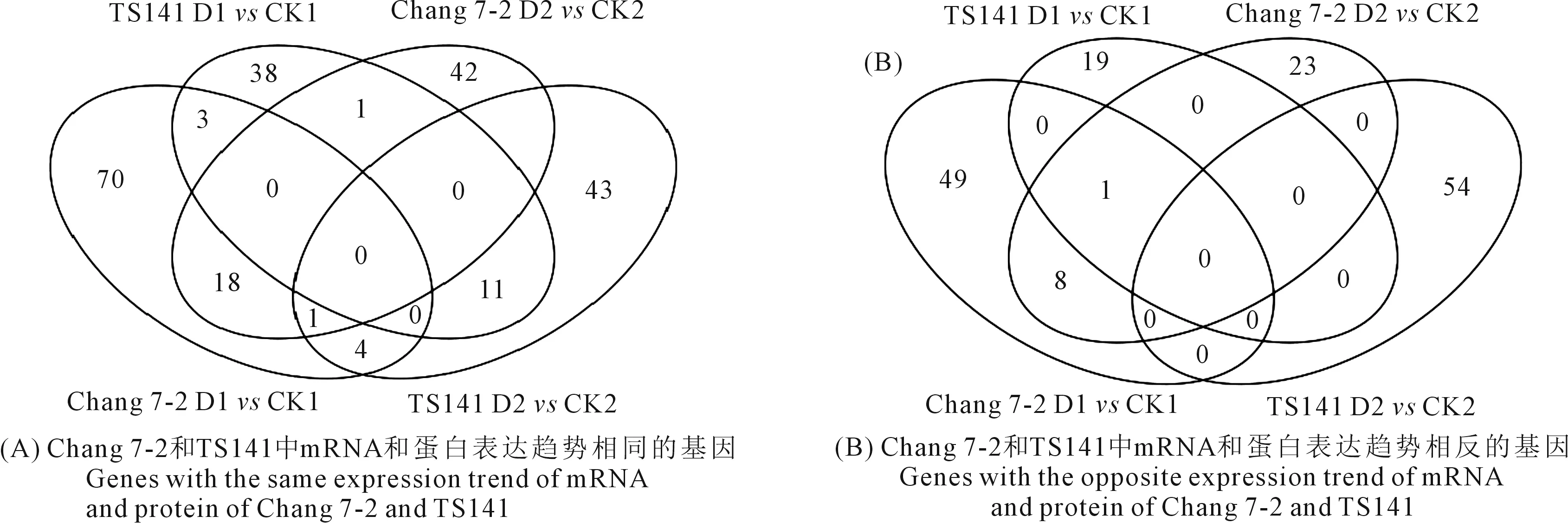
圖3 兩玉米自交系在不同處理下DEGs和DEPs的Venn圖
2.2.3 GO富集關聯分析 對兩個組學中差異表達蛋白和基因進行GO功能注釋分析,結果表明Chang 7-2 D1vsCK1、Chang 7-2 D2vsCK2、TS141 D1vsCK1和TS141 D2vsCK2等4個比較組的GO功能富集總體分為細胞成分(Cellular component, C)、分子功能(Molecular function, F)和生物過程(Biological process , P),而干旱處理下,比較組Chang 7-2 D1vsCK1和Chang 7-2 D2vsCK2所關聯DEGs/DEPs的主要細胞組分包括細胞內非膜-b(Intracellular non-membrane-b)、非膜結合細胞器(Non-membrane-bounded organel)、細胞外圍(Cell periphery);生物學過程主要有氧化-還原過程(Oxidation-reduction process)、脅迫反應(Response to stress)、碳水化合物代謝過程(Carbohydrate metabolic proce);分子功能主要包括金屬離子結合(Metal ion binding)、陽離子結合(Cation binding)、氧化還原酶活性(Oxidoreductase activity)。比較組TS141 D1vsCK1和TS141 D2vsCK2所關聯到的DEGs/DEPs主要細胞組分包括Intracellular non-membrane-b、Non-membrane-bounded organel和核糖核蛋白復合物(Ribonucleoprotein complex);分子功能主要包括Oxidoreductase activity和Metal ion binding;其生物學過程主要有脅迫反應(Response to stress)和刺激反應(Response to stimulus)。此外,我們還發現兩玉米自交系幼苗根系所關聯到的DEGs/DEPs均富集到脅迫響應(Response to stress)過程,即遭遇干旱脅迫后,其幼苗根系會立即對干旱脅迫做出響應,并通過促進根系內過氧化物酶活性、氧化還原酶活性等多種酶活性來抵御干旱。
2.2.4 Pathway富集關聯分析 利用KEGG pathway數據庫對Chang 7-2 D1vsCK1、Chang 7-2 D2vsCK2、TS141 D1vsCK1和TS141 D2vsCK2等4個比較組的轉錄組和蛋白質組數據以及所關聯到的DEGs/DEPs進行通路富集分析,結果表明,差異基因和差異蛋白在各代謝通路中富集顯著性分為3大類:轉錄組與蛋白質組都顯著富集、轉錄組與蛋白質組任意一個組學數據顯著富集和轉錄組與蛋白質組均無顯著富集。而我們僅對轉錄組與蛋白組兩個組學都顯著富集的通路進行了分析,發現4個比較組所關聯到的DEGs/DEPs主要富集的通路包括苯丙素的生物合成(Phenylpropanoid biosynthesis)、次生代謝產物的生物合成(Biosynthesis of secondary metabolites)和核糖體(Ribosome)(表4)。由此發現,玉米幼苗在應對干旱脅迫時會發生一系列復雜的代謝活動,而苯丙素的生物合成、次生代謝產物的生物合成和核糖體等相關代謝通路可能是根系響應干旱脅迫的主要通路。
2.2.5 qRT-PCR驗證 為了驗證干旱脅迫下兩玉米自交系轉錄組和蛋白質組數據對干旱脅迫響應的可靠性,我們通過qRT-PCR驗證了在干旱處理下昌7-2和TS141中表達水平不同的9個基因(表5、圖4)。我們發現這9個基因在兩個自交系中的基因表達水平與RNA-Seq和iTRAQ結果呈現相同趨勢,這表明兩組學測序數據是可靠的。
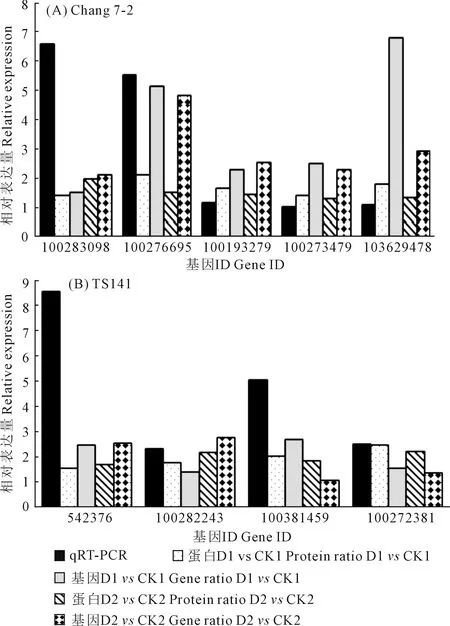
圖4 qRT-PCR驗證所選基因的相對表達量
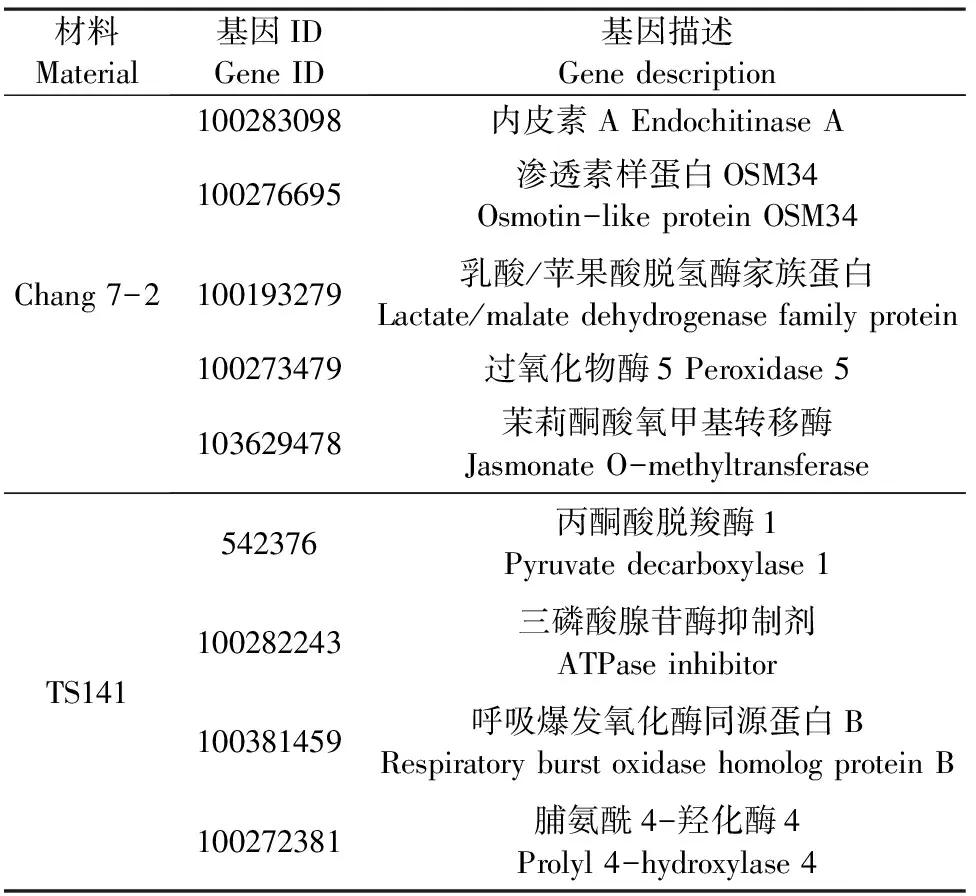
表5 用于qRT-PCR分析的基因
3 討 論
為了進一步剖析不同玉米自交系對干旱脅迫的響應機制,采用耐旱自交系昌7-2和干旱敏感自交系TS141作為試驗材料,在干旱處理12 d后對其幼苗根系進行轉錄組學分析和蛋白質組學分析,并對其根系差異基因與差異蛋白組表達趨勢相同的部分基因進行了進一步分析。
3.1 耐旱玉米自交系昌7-2根系對干旱的響應機制
Jasmonate O-methyltransferase作為催化茉莉酸合成MeJA最重要的限速步驟[23],是MeJA合成途徑遺傳改造的最重要位置。研究表明,茉莉酸類化合物作為信號分子參與調控植物的生長發育與代謝途徑,響應低溫、干旱和鹽等多種非生物脅迫[24]。此外,茉莉酸類物質在植物對逆境防御及脅迫傳遞信號等方面有著極其重要的作用。MA等[25]和WU等[26]研究結果也表明,茉莉酸甲酯緩解物質通過調節小麥(Triticumaestivum)和甘藍(Brassicaoleracea)氣孔運動及相關基因表達來調節干旱缺水對作物所帶來的損傷,提高植物的抗旱性。本研究在昌7-2中也發現一個編碼Jasmonate O-methyltransferase的基因(103629478)在轉錄組和蛋白質組中均上調表達,推測其在干旱脅迫下通過調節幼苗根系的氣孔開度抗氧化酶活性來抵御干旱。
幾丁質酶(Endochitinase)是一種能夠將幾丁質分解成幾丁質單糖—N-乙酰基葡萄糖的酶,幾乎在植物各器官中都有分布。有研究表明,當植物遭受干旱、低溫、凍害及重金屬離子等多重脅迫時,其組織內幾丁質酶的活性會快速升高并水解病原菌細胞壁的幾丁質,抑制病原菌在植物體內的生長繁殖[27]。近年來,關于幾丁質酶基因的研究在不同植物中也不斷被報道,陳鵬等[28]通過對小麥進行干旱脅迫后發現,隨著干旱脅迫程度的增加,其體內幾丁質酶的活性也在不斷上升,復水處理后又逐漸恢復至正常水平,表明在干旱脅迫下幾丁質酶可以提高植物抗旱性。MUOKI等[29]通過對茶樹進行環境脅迫后發現,幾丁質酶基因Chi與Thaumatin蛋白可共同響應干旱脅迫,進而促進茶樹的抗旱性,減弱干旱對茶樹的損害。在本研究中,我們也發現當昌7-2在遭遇干旱脅迫后其根系內幾丁質酶相關基因出現差異上調表達。
此外,當昌7-2遭受干旱脅迫時,其體內與熱激蛋白(100283821,Hsp20)、滲透蛋白(100276695,Osmotin-like protein OSM34)及過氧化物酶(100285894、100273479、541674、107548102)相關基因及蛋白也發生上調或下調表達。研究表明,熱激蛋白作為植物響應非生物脅迫的重要蛋白之一,在植物耐寒耐旱、保護植物相關蛋白完成正確的折疊、翻譯和聚集及結構功能方面發揮著重要作用[30],目前已發現多種類型熱激蛋白,例HSP60、HSP70和HSP90等家族[31-32]。另外,干旱脅迫下,滲透調節蛋白基因和過氧化物酶(POD)對植物應對逆境脅迫有非常重要的作用。Osm基因(即ProBA基因)是一種可增強植物細胞抗滲透脅迫能力的蛋白質,其主要功能為在逆境脅迫下促進脯氨酸在細胞內的合成和累積[33],而在逆境脅迫下不同類型的POD相互作用可有效清除植物體內活性氧。因此,我們推測昌7-2受到干旱脅迫后,其幼苗根系會通過調節體內熱激蛋白和滲透蛋白相關基因上調,同時通過調控不同類型的過氧化物酶上或下調來減輕干旱所帶來的損傷。
3.2 玉米干旱敏感自交系TS141根系對干旱的響應機制
當對TS141進行干旱處理后,我們發現其mRNA和蛋白表達趨勢相同的基因主要編碼6-磷酸葡萄糖酸內酯酶(100285843)、核苷二磷酸激酶I(100217209)、脯氨酸-4-羥化酶(100272381)及丙酮酸脫氫酶(541919、542376)等一系列基因上、下調表達來減輕其對干旱脅迫的響應,而這些基因的功能多數與糖酵解生成丙酮酸的途徑相關聯。丙酮酸脫羧酶(Pyruvate decarboxylase, PDC)是一種以焦磷酸硫胺素為輔酶的羧基裂解酶,是發酵途徑中的關鍵酶。PDC通過與PDH競爭代謝丙酮酸參與植物體內能量代謝和物質代謝,響應植物多種非生物脅迫,如缺氧、高鹽及低溫等[34-35]。研究表明,在逆境脅迫下丙酮酸對植物的呼吸作用有交替作用,而植物保持較高的交替呼吸本身便是一種抗逆機制,對植物抗旱等方面具有重要意義[36-37]。因此,我們推測當TS141感受到干旱脅迫后,主要通過誘導其糖酵解途徑來促進根系進行交替呼吸,進而抵御干旱所帶來的損傷。
4 結 論
本研究以兩份耐旱性不同的玉米自交系昌7-2和TS141為試材,對其幼苗進行12 d干旱脅迫后進行轉錄組分析和蛋白質組學分析,以剖析不同自交系對干旱脅迫的響應機制及其抵御反應。結果表明,當玉米幼苗根系遭遇干旱脅迫后,為抵御干旱脅迫其體內會發生一系列生理生化反應,且不同自交系的根系在轉錄和翻譯變化方面也存在明顯的差異。此外,在干旱脅迫下,我們在昌7-2和TS141中分別鑒定了19、11個mRNA和蛋白表達趨勢相同的基因、9和0個趨勢相反的基因,并且這些基因主要涉及苯丙素的生物合成、次生代謝產物的生物合成和核糖體等代謝通路,可為今后了解玉米耐旱性的分子機制提供一定的參考。
猜你喜歡
當代陜西(2021年17期)2021-11-06 03:21:36
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
學苑創造·A版(2018年11期)2018-02-01 06:29:20
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
讀者(2017年5期)2017-02-15 18:04:18
爆笑show(2016年7期)2017-02-09 09:36:13
山東工業技術(2016年15期)2016-12-01 05:31:22
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
