成齡轉基因銀中楊試驗林外源基因水平轉移及土壤微生物連年監測*
2022-03-23 06:54:54張偉溪王顏波丁昌俊朱文旭蘇曉華
林業科學 2022年1期
張偉溪 王顏波,2 丁昌俊 朱文旭,3 蘇曉華
(1.林木遺傳育種國家重點實驗室 國家林業和草原局林木培育重點實驗室 中國林業科學研究院林業研究所 北京 100091;2.南昌工程學院 南昌 330099; 3.沈陽農業大學 沈陽 110866)
自從Parsons等(1986)證實楊樹(Populus)可以進行遺傳轉化、外源基因可在林木細胞中表達以來,林木基因工程研究已經取得長足發展。據統計,全球至少有35個國家對百余個樹種進行了遺傳轉化研究,其中楊樹、番木瓜(Carica)、松樹(Pinus)、柳樹(Salix)、核桃(Juglans)、蘋果(Malus)、櫻桃(Cerasus)、樟樹(CinnamomumSect.Camphora)、懸鈴木(Platanus)、桉樹(Eucalyptus)、云杉(Picea)、美國栗(Castaneadentata)、美國榆(Ulmusamericana)等200多個轉基因林木已進入田間試驗階段或已批準商品化(FAO,2004;Straussetal.,2011;Sedjo,2005;Klockoetal.,2018;Muhretal.,2018;Changetal.,2018;康向陽,2020)。轉基因林木的改良性狀主要集中在抗病、抗蟲等抗生物脅迫,抗除草劑,抗旱、耐鹽等抗非生物脅迫,降低木質素合成、增加纖維素合成等材性改良以及發育調控等方面(Londoetal.,2010)。由此可見,轉基因林木主要應用于木材生產或在生態景觀建設等方面,并不像轉基因作物一樣直接或間接涉及人類生命安全相關問題。但是,轉基因林木的環境釋放及推廣應用的潛在生態風險仍不容忽視,其生態安全性監測仍需持續跟蹤進行,以便有效地規避其生態安全問題,從而加速轉基因林木的商業化應用和推廣。因此,加強轉基因生物安全的基礎研究,提高對轉基因生物安全性評價水平,制定有效的生態風險監測和管理策略尤為重要。
目前,轉基因植物生態安全評估主要集中在轉基因植物外源基因逃逸以及對生態系統的影響方面(Londoetal.,2010;Richardsonetal.,2011),如外源基因可能存在通過花粉、種子以及無性繁殖器官等媒介向其近緣種或其他生物群體進行基因漂移和水平轉移的風險(Wangetal.,2018;Klockoetal.,2018)。土壤微生物是土壤生態系統的一個重要組成部分,參與土壤結構形成、分解有機質和礦物質,與植物根部營養吸收密切相關。因此,轉基因植物的外源基因可能通過水平轉移整合到土壤微生物的基因組中,從而改變其遺傳特性與功能(Luetal.,2018)。例如,轉基因西瓜(Citrulluslanatus)中35S啟動子基因存在向土壤微生物轉移的可能(Keese,2008),在轉Bt玉米(Zeamays)、棉花(Gossypium)等作物的土壤中檢測到了不同水平的Bt 蛋白(Stotzky,2005;Icozetal.,2008)。但也有研究發現,外源基因發生水平轉移的頻率低于自然轉化的頻率,從而給環境帶來的風險可以忽略(Keese,2008),如對成熟期雄性轉Bt基因歐洲黑楊(Populusnigra)人工林基因流頻率和距離研究發現,距轉Bt楊樹人工林0 m處產生Bt種子的頻率為0~0.15%,500 m處產生Bt種子的頻率為0~0.02%,在自然條件下,轉Bt楊樹人工林通過花粉或種子產生的基因流水平極低(Huetal.,2017)。另外,外源基因表達對轉基因植物目標性狀改變有可能影響相關代謝途徑產物的積累,從而通過植物落葉、根系等對根際土壤微生物及其群落的多樣性產生潛在影響(Dauduetal.,2009;Gaoetal.,2014)。如轉基因水稻(Oryzasativa)生長旺盛時期土壤中細菌數量顯著增加(陳曉雯等,2011);轉MdSOS2L1基因蘋果(Malusdomestica)植株根際微生物多樣性研究表明,轉基因與野生型植株根際微生物群落豐富度和多樣性存在較大差異,轉MdSOS2L1植株根際酸桿菌門(Acidobacteria)豐度增加較多,藍藻門(Cyanophyta)、放線菌門(Actinobacteria)豐度略有下降,同時,轉基因植株根際土壤的蘋果酸、檸檬酸、草酸含量明顯升高,轉基因植株根際微生物的差異極有可能與MdSOS2L1基因介導的有機酸分泌有關(王曉娜等,2018)。因此,土壤微生物的種類和數量通常可作為轉基因植物生態安全性檢測的重要指標。目前有關轉基因植物系統生態安全性監測研究多集中在大豆(Glycinemax)、玉米、棉花、水稻等農作物方面(Icozetal.,2008;Keetal.,2014;Lietal.,2017)。轉基因林木雖有研究,但多以苗期或幼齡的轉Bt、Cry1A、Cry3A等基因林木為主,并未發現外源基因向土壤微生物轉移的現象(胡建軍等,2004;侯英杰等,2009;魏冰等,2009;張雁等,2012;李霞等,2011;Tranetal.,2018;Zuoetal.,2018;孫偉博等,2020),對成齡期轉基因林木監測研究較少(Zhuetal.,2016;朱文旭等,2017;Huetal.,2017;呂威等,2018)。與集約化程度較高的農作物相比,轉基因林木栽培環境復雜、生長周期長、經營管理相對粗放,且多為風媒傳粉,因此,對成齡期轉基因林木試驗林的土壤微生物進行長期持續性監測非常必要。
1 材料與方法
1.1 試驗材料
選取轉抗逆轉錄因子基因(JERF36)銀中楊(Populusalba×P.berolinensis)——抗逆1號楊(ABJ01)及非轉基因銀中楊(9#,對照)為研究材料,抗逆1號楊(ABJ01)是本實驗室早期通過農桿菌介導法將連有35S啟動子和NOS終止子的JERF36基因導入銀中楊中,并且通過PCR、RT-PCR、Southern 雜交等分子檢測獲得外源基因成功整合并穩定表達的轉基因銀中楊,其中JERF36基因是從番茄(Lycopersiconesculentum)中克隆獲得的茉莉酸乙烯應答元件基因,編碼AP2/EREBP類植物轉錄因子,能專一結合GCC-box,在植物中能夠激活下游抗逆相關基因表達,提高抗逆性。報告基因nptⅡ來自于細菌,編碼新霉素磷酸轉移酶,能賦予細胞抗卡那霉素能力(Lietal.,2009)。抗逆1號楊已獲得環境釋放及生產性試驗行政許可,與非轉基因對照相比,其形態未發生明顯變化,抗旱耐鹽性顯著提高,于2015年獲得國家植物新品種保護權。
1.2 試驗地概況及樣品采集
試驗林位于北京市房山區韓村河東營苗圃(115°58′E,39°37′N),2006年春季造林,轉基因株系及其對照均按隨機區組,正方形種植100株(行10 株、列10 株),株行距為2 m × 2 m,造林總面積為0.66 hm2。試驗地的地勢、地貌、氣溫、降雨、植被、栽培管理等自然條件和人為管理均一致,整個試驗階段林地不進行任何肥水及噴施農藥管理。
分別于2015年5—8月、2016年5—10月及2017年5—10月每月中旬取樣1次,選擇連續3天以上晴天的上午在試驗林地內使用土鉆鉆取非根際土壤。隨機選取抗逆1號楊(ABJ01)和非轉基因對照(9#)各3株進行取樣,以植株為中心,每株選取與主干50 cm距離的東南西北4個點進行取土,去除表層腐殖質層,留取 10~20 cm土柱,將土樣中的石塊和植物的根殘體等雜物去掉,裝入無菌的封口袋中混合均勻,放在冰盒內帶回實驗室,4 ℃冰箱內保存備用。該土壤樣品用于研究微生物數量變化。2017年7月分別隨機選取抗逆1號楊(ABJ01)和非轉基因對照(9#)各10株采集幼嫩葉片,并按非根際土壤取樣方法采集林下雜草,每種雜草取2~3片葉子混合;同時采集試驗林地旁(約10 m)行道樹毛白楊(Populustomentosa)、歐美楊(Populus×euramericana)雌株樹下自然生長的實生楊樹幼苗葉片,延路每隔10 m取1次樣品,取樣10次,每株楊樹幼苗取3~5片葉子混合。樣品用冰盒帶回實驗室,置于-80 ℃ 低溫冰箱內保存備用。
1.3 試驗方法
1.3.1 植物及土壤基因組DNA的提取 采用 DNeasy Plant Mini Kit (Qiagen,Germany)提取抗逆1號楊(ABJ01)、非轉基因對照(9#)、林下雜草以及試驗林旁行道樹下楊樹幼苗葉片基因組DNA;使用 PowerSoil? DNA Isolation Kit(Mobio,America)提取2017年7月非根際土壤中微生物的總DNA,具體操作參照試劑盒使用手冊。
1.3.2 細菌基因組DNA的提取 選取2017年7月采集的抗逆1號楊(ABJ01)、非轉基因對照(9#)土壤樣本的10-1土壤懸濁液涂布在含卡那霉素濃度50 mg·L-1的細菌選擇培養基平板中篩選抗性菌株,共獲得2株。分別挑取其單菌落使用牛肉膏蛋白胨液體培養基培養,吸取5 mL處于對數生長期的菌液5 000 r·min-1離心1 min,棄上清。采用細菌基因組DNA提取試劑盒(TIANGEN,China)提取2個菌株的基因組DNA。
1.3.3 土壤及細菌DNA質量檢測 以土壤微生物總DNA和細菌DNA為模板,去離子水為陰性對照。以細菌16S rRNA通用引物(上游引物:5′-ACTCCTACGGGAGGCAGCAG-3′;下游引物:5′-GGACTACHVGGGTWTCTAAT-3′)進行PCR擴增,反應條件為94 ℃ 3 min;94 ℃ 30 s,55 ℃ 30 s,72 ℃ 45 s,30個循環;最后72 ℃延伸10 min,預期擴增片段長度為468 bp。
1.3.4 外源基因水平轉移情況檢測 以上述提取的所有DNA為模板,質粒DNA為陽性對照,去離子水為陰性對照。根據JERF36序列設計特異引物(上游引物:5′-CTCCCTTGAACATTGCTTG-3′;下游引物:5′-TGATTGCCCGTCAACATAC-3′)進行PCR擴增,反應條件為95 ℃ 5 min;95 ℃ 1 min,58 ℃ 1 min,72 ℃ 1 min,共30個循環;最后72 ℃ 延伸10 min,預期擴增片段長度為278 bp。
1.3.5 土壤微生物數量測定 采用稀釋平板法培養土樣中的細菌、真菌、放線菌。稱取 10 g 土壤樣品倒入裝有90 mL無菌水和無菌小玻璃珠(10粒)的三角瓶中,置于搖床上(200 r·min-1)室溫振蕩30 min,隨后用無菌水稀釋土壤懸濁液(使用移液槍準確吸取0.1 mL土壤懸浮液注入0.9 mL的無菌水中即得到終濃度為10-1土壤懸濁液),其中,細菌稀釋液終濃度分別為10-3、10-4、10-5的土壤懸濁液,放線菌稀釋液終濃度分別為10-2、10-3、10-4的土壤懸濁液,真菌稀釋液終濃度分別為10-1、10-2、10-3的土壤懸濁液。吸取50 μL稀釋懸濁液均勻涂在牛肉膏蛋白胨培養基(培養細菌)、馬丁(Martin)孟加拉紅-鏈霉素培養基(培養真菌)、改良淀粉銨鹽培養基(培養放線菌)上,放線菌置于28 ℃的培養箱內進行培養,細菌和真菌置于35 ℃的培養箱內進行培養。2天后統計真菌的菌落數,2~4天后統計細菌的菌落數,5天左右統計放線菌菌落數。每個菌種每個濃度培養3次。同時稱取50 g的待測土樣,用牛皮紙信封裝好,放于烘箱中,105 ℃烘烤,烘至恒質量后稱質量,計算土壤含水量,從而計算出每克干土中土壤微生物的菌落數,菌落數=(菌落平均數×稀釋倍數)/[接種量毫升數×(1-含水量)]。采用SPSS 17.0 軟件進行方差分析和多重比較。
2 結果與分析
2.1 外源基因JERF36的水平轉移
采用外源基因JERF36的特異性引物分別對抗逆1號楊(ABJ01)、非轉基因對照(9#)、林下雜草、林下土壤、試驗林旁行道樹下楊樹實生苗葉片、土壤中篩選出的抗性菌株基因組DNA進行PCR檢測(圖1A),采用細菌16S rRNA通用引物對林下土壤、土壤中篩選出的抗性菌株基因組DNA進行PCR檢測(圖1B)。結果顯示,在抗逆1號楊(ABJ01)的基因組DNA中能擴增出JERF36目的片段(278 bp),而非轉基因對照(9#)、林下雜草、林下土壤、試驗林旁行道樹下楊樹實生苗葉片以及2個抗性菌株基因組DNA均沒得到擴增產物,且在土壤及菌株DNA,均能檢測到微生物DNA,說明土壤及抗性菌株基因組DNA質量合格。以上說明種植11年后,JERF36基因仍穩定存在于轉基因植株基因組中,并未發現向林地土壤、周邊雜草等發生水平轉移。
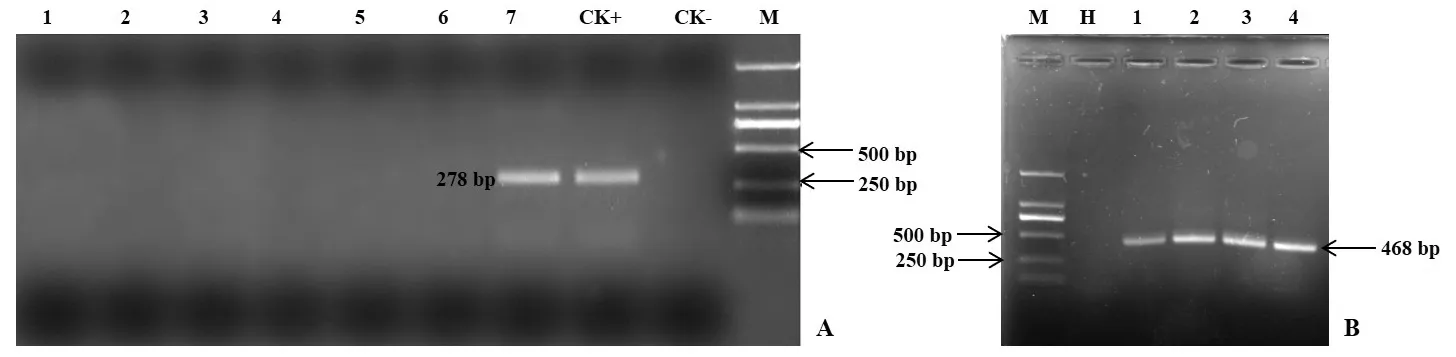
圖1 外源基因JERF36 PCR檢測Fig.1 PCR detection of exogenous gene JERF36A:外源基因JERF36 PCR檢測(1:非轉基因對照9#;2:林下雜草;3:林下土壤總DNA;4:試驗林旁行道樹下楊樹實生苗葉片;5:抗性菌株1;6:抗性菌株2;7:轉基因植株ABJ01;CK+:質粒陽性對照;CK-:去離子水陰性對照);B:16S rRNA PCR檢測(H:去離子水陰性對照;1:林下土壤總DNA 1;2:林下土壤總DNA 2;3:抗性菌株1;4:抗性菌株2;M:DNA marker)。A:PCR detection of exogenous gene JERF36 (1:Non-transgenic control 9#;2:Understory weed;3:Total DNA in soil;4:Seedling leaves of roadside poplar beside the test field;5:Kanamycin-resistant strain 1;6:Kanamycin-resistant strain 2;7:Transgenic poplar ABJ01;CK+:Plasmid positive control;CK-:Deionized water negative control);B:PCR detection of 16S rRNA(M:DNA marker;H:Deionized water negative control;1:Total DNA in soil 1;2:Total DNA in soil 2;3:Kanamycin-resistant strain 1;4:Kanamycin-resistant strain 2).
2.2 林下土壤微生物的數量
分別對9年生(2015年)、10年生(2016年)、11年生(2017年)抗逆1號楊(ABJ01)及非轉基因對照(9#)林下土壤中微生物總數量進行檢測并統計分析(圖2A)。3年連續性觀測顯示,各年份成齡抗逆1號楊與非轉基因對照間林下土壤中微生物總數量均無顯著差異;不同年份間的抗逆1號楊和非轉基因對照的林下土壤微生物數量均略有差異,但無明顯變化規律。其中6—8月是土壤中微生物快速增長期,其總量呈現2015年>2016年>2017年;而3個年份間6—8月氣溫并無顯著差異(圖2B),但降雨天數差異明顯且呈現2015年(44天)>2016年(37天)>2017年(31天)(圖2B)。在2016年和2017年的9—10月土壤中微生物總量、氣溫迅速下降,但在2016年9—10月的降雨天數維持穩定,2017年降雨天數先下降后上升,2017年9—10月試驗林中土壤微生物總數量顯著高于2016年。由此推測可能由于降雨差異導致土壤濕度變化,進而造成不同年份間抗逆1號楊和非轉基因對照的林地土壤微生物數量差異,而抗逆1號楊未對林地土壤微生物總量造成影響。
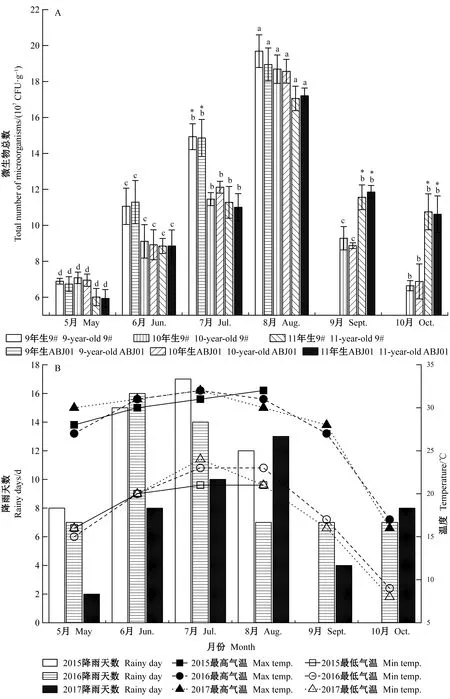
圖2 9~11年生(2015—2017年)楊樹林地土壤微生物總數量以及不同月份氣溫和降雨天數Fig.2 Total number of microorganisms of 9-11 year-old(2015-2017)poplar test field and the temperature and raining days in different months9#:非轉基因對照;ABJ01:轉基因植株。下同。不同小寫字母表示不同月份微生物數量在0.05水平上存在顯著性差異;“*”表示不同年份間微生物數量在0.05水平上存在顯著性差異。9#:Non-transgenic plants;ABJ01:Transgenic plants.The same below.Different lowercase letters indicate significant differences in the number of microorganisms in different months at the 0.05 level;“*”means significant differences in the number of microorganisms in different years at the 0.05 level.
分別對9年、10年、11年生抗逆1號楊(ABJ01)及非轉基因對照(9#)林地土壤中細菌、真菌和放線菌數量進行檢測并統計分析(圖3)。結果表明:9年、10年、11年生抗逆1號楊和非轉基因對照林地土壤3大類微生物數量在樹木生長季呈現先上升后下降的變化趨勢,其中在生長旺季的7月和8月林地土壤微生物數量最高,顯著高于開始生長階段(5月和6月)和停止生長階段(9月和11月)。3個年份中,細菌和放線菌的數量均在每年的8月達到最高值;9年生、10年生抗逆1號楊和非轉基因對照林地土壤中真菌數量也在8月達到最高值,11年生的林地土壤中真菌數量在7月峰值最高。抗逆1號楊與非轉基因對照間林地土壤中3大類微生物數量均無顯著差異,變化趨勢一致。表明抗逆1號楊未對林地土壤中細菌、真菌和放線菌數量造成影響,而其中的微小差異可能是由氣候等其他因素造成的。
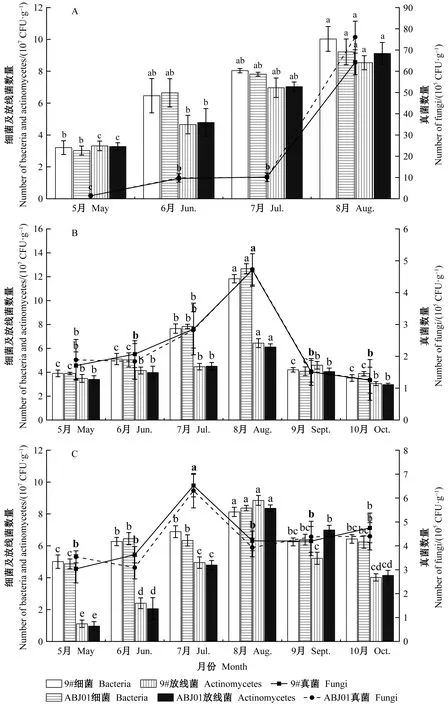
圖3 9~11年生(2015—2017年)林地土壤細菌、真菌和放線菌數量Fig.3 Total numbers of bacteria,actinomycetes,and fungi in soil of 9-11 years old (2015—2017)poplar test fieldA:9年生楊樹5—8月土壤微生物數量;B:10年生楊樹5—10月土壤微生物數量;C:11年生楊樹5—10月土壤微生物數量。不同小寫字母表示不同月份微生物數量在0.05水平上存在顯著性差異。A:Total number of soil microorganisms in 9-year-old poplar plantation from May to August;B:Total number of soil microorganisms in 10-year-old poplar plantation from May to October;C:Total number of soil microorganisms in 11-year-old poplar plantation from May to October.Different lowercase letters indicate significant differences in the number of microorganisms in different months at the 0.05 level.
通過對不同年份間細菌、真菌和放線菌的比例分析(圖4)發現,3個年份中,抗逆1號楊和非轉基因對照林地土壤中3類微生物的比例相似,從高到低依次是細菌、放線菌和真菌;不同年份間3類微生物所占比例略有差異,但并沒有出現某一類微生物急速增加或降低的現象。說明抗逆1號楊并沒有破壞主要微生物種類之間的平衡。
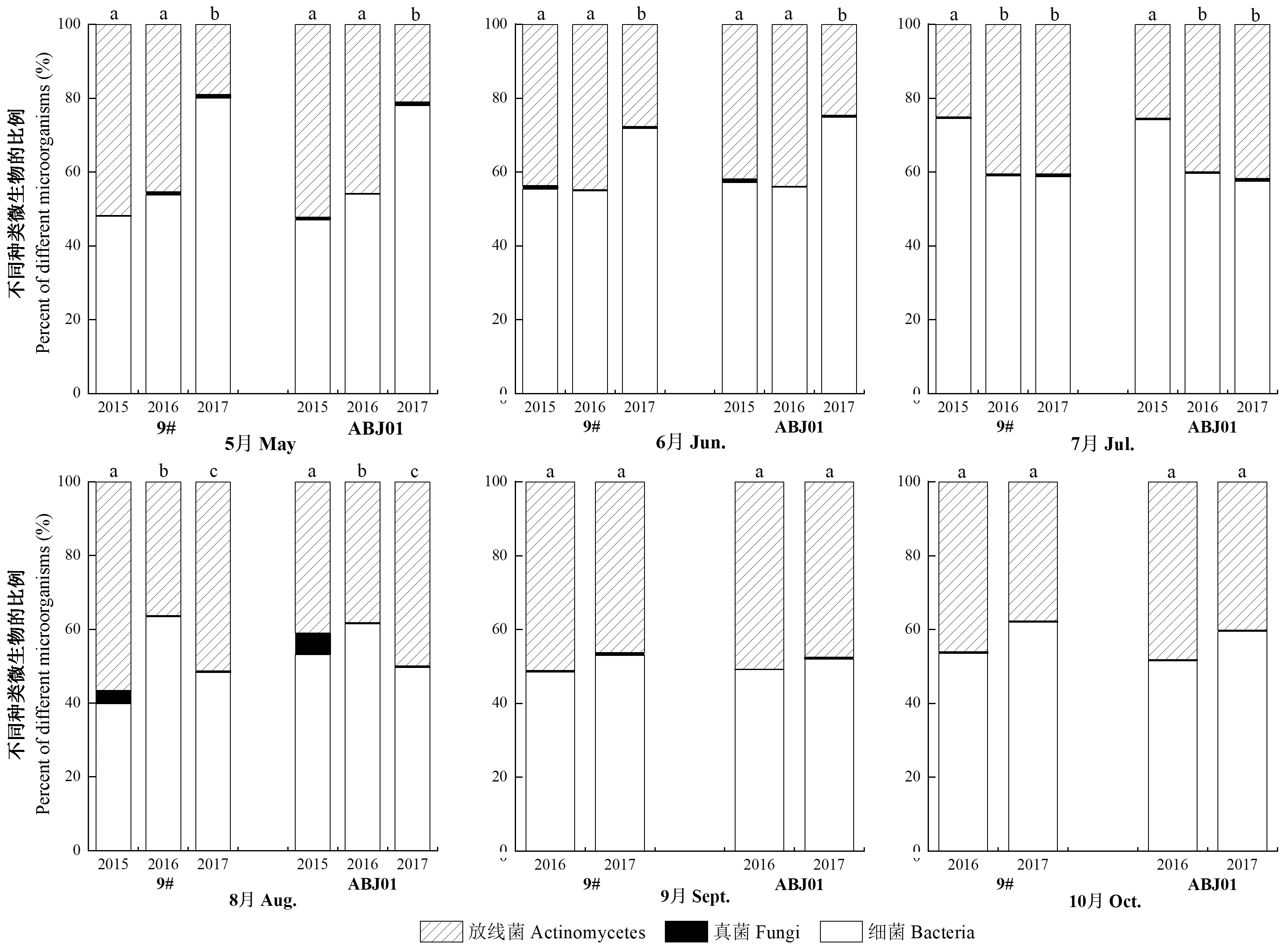
圖4 不同年份(2015—2017年)林地土壤中微生物比例Fig.4 Proportions of microorganisms in plantation soil in different years(2015—2017)不同小寫字母表示不同年份間微生物比例在0.05水平上存在顯著性差異。The different lowercase letters mean significant deference of the microbial proportion in different years at the 0.05 level.
綜上所述,對試驗林內抗逆1號楊與非轉基因對照在成齡期后的連續監測顯示,試驗林地土壤微生物的數量和種類平衡并未發現顯著變化,而環境變化如降雨差異可能是不同年份微生物數量差異的主要原因,抗逆1號楊與非轉基因對照林地微生物數量略微差異需要結合環境因素做進一步研究分析。
3 討論
隨著越來越多轉基因植物的大面積田間種植,其對生態環境的影響一直備受矚目。而外源基因在轉基因受體植物中,特別是多年生林木中是否會發生逃逸現象是檢測轉基因植物安全性的首要問題(Frankenhuyzenetal.,2004)。盡管早期研究發現一些轉基因林木中存在外源基因沉默現象(Jouaninetal.,2000;Kumaretal.,2001),但眾多研究表明外源基因能在轉基因林木中穩定存在并表達。例如,對5年生轉基因741楊[Populusalba×(P.davidiana+P.simonii)×P.tomentosa]田間檢測發現,外源基因Cry1Ac、Cry3A仍能穩定表達,未發現丟失和沉默現象(Zuoetal.,2018);轉基因雜種山楊(Populustremula×P.tremuloides)在溫室生長18年后,外源基因rolC依然穩定表達,并未出現外源基因丟失和沉默現象(Fladungetal.,2013);經過7年田間試驗的轉Bt基因歐洲黑楊(Populusnigra)的外源基因仍存在于轉基因楊的基因組中(胡建軍等,2004);轉基因長春花(Catharanthusroseus)在5年后依然檢測到外源基因rolA、rolB、rolC的存在和穩定表達(Vermaetal.,2015)。本試驗使用外源基因特異引物對轉基因銀中楊(抗逆1號楊)和非轉基因對照(銀中楊)的基因組DNA進行PCR檢測結果表明,抗逆1號楊在田間種植11年后,外源基因仍穩定存在于轉基因楊樹的基因組中,未發生外源基因丟失現象。
基因水平轉移現象在原核生物中廣泛存在,在真核生物中也有一些相關報道(Kyndtetal.,2015;Quispe-Huamanquispeetal.,2019;Matveevaetal.,2019)。林木生長周期長,轉基因樹木種植大田后,每年試驗林中產生大量根茬、落葉、花粉、果實等,經過多年累積,外源基因可能通過根際分泌物、土壤微生物降解和吸收有機物的過程進入土壤體內,影響周圍土壤環境中的微生物群落結構及其多樣性等(Gaoetal.,2014;Aslametal.,2017;Figueredoetal.,2018)。因此外源基因是否發生水平轉移,土壤中微生物的種類、群落數量等是否發生變化是對轉基因林木進行安全性評價的重要方面。對包含3 種編碼不同抗性蛋白的外源基因(PeTLP、Cry1Ac、AtGols)的轉基因南林895楊試驗林進行安全性評估,未發現外源基因向土壤微生物中轉移的現象(孫偉博等,2020)。5年生轉基因741楊的生態安全性評價表明,在土壤和林下雜草中未檢測到外源基因,外源基因沒有發生基因轉移,多年生轉基因楊樹對土壤理化性質和土壤微生物群落結構沒有產生影響,微生物群落結構主要受地理位置和季節變化的影響(Zuoetal.,2018)。大田種植12年后轉基因毛白楊暫未對土壤可培養微生物數量造成顯著影響(呂威等,2018)。轉Bt基因棉花根際土壤微生物的活性并不受Bt蛋白的影響,其主要的影響因素是土壤的鹽分(Luoetal.,2017)。本實驗室早期研究顯示,8年生轉多基因歐美楊 (Populus×euramericana‘Guariento’)未發現外源基因轉移,其與非轉基因對照之間林地土壤細菌多樣性和群落結構無顯著差異,其對土壤微生物系統沒有明顯的影響,轉基因楊樹的土壤微生物數量主要受季節變化影響(朱文旭等,2015;2017;Zhuetal.,2016)。對不同區域抗逆1號楊及非轉基因對照內生菌多樣性研究表明,外源基因導入并未對抗逆1號楊根、莖中內生菌(細菌和真菌)群落結構及多樣性產生影響,微生物的群落結構主要取決于土壤pH值和有機質含量(Wangetal.,2019)。本試驗也未在抗逆1號楊試驗林內的雜草和林地土壤微生物基因組中檢測到外源基因。由于本試驗材料在轉JERF36基因的同時引進了報告基因nptⅡ,為了進一步驗證土壤中抗卡那霉素的細菌是否被整合上外源基因,本試驗篩選出2個抗性菌株,但均未得到外源基因的擴增產物,充分表明外源基因未發生水平轉移。此外,抗逆1號楊與非轉基因對照林地土壤中的細菌和真菌數量無顯著差異;放線菌僅在2015年(楊樹9年生)的7月份略有差異,可能是環境變化引起,隨后的2年中均無顯著差異。也進一步證實11年生抗逆1號楊并未對土壤微生物的種類、數量造成顯著影響。
此外,還有通過檢測試驗林內根際土壤微生物群落結構、表層土凋落物、花粉中外源基因表達等方面對多年生轉基因楊樹對土壤環境的安全性進行檢測,但均未發現顯著影響。如在內蒙古地區轉基因試驗林地內的轉基因銀中楊安全性檢測顯示,轉基因銀中楊并未影響其根際土壤微生物的數量及主要類群的數量(呂秀華,2017;2018);種植13年的轉基因三倍體毛白楊枯落物中的外源基因暫未水平轉移到根際土壤可培養細菌基因組中(呂威等,2019)。雖然上述研究并未發現轉基因楊樹對根際土壤等產生顯著影響,但本研究中并未進行相關方面安全性檢測,因此在今后的研究中可加強本試驗林在該方面的相關檢測內容,以期更加全面地進行轉基因楊樹試驗林的安全性檢測,為轉基因林木安全性評價提供依據。
4 結論
轉基因林木對生態的影響是一個長期而復雜的過程,目前尚沒有公認的評估轉基因植物生態安全性的方法和標準。本試驗對轉JERF36基因銀中楊生態安全性的初步分析表明,外源基因在轉基因銀中楊中穩定存在,沒有發現向周邊環境水平轉移,并未發現外源基因導入對林地土壤微生物的數量和群落結構造成影響。森林中土壤微生物的組成和結構非常復雜,關于轉基因林木生態及生物安全性監測需要進行更加系統、全面和長期的研究。
