GSK-3β參與氯化鋰修飾顱內感染后癲癇發作的可能機制
2022-03-23 07:11:44李志民余巨明蔣國會
重慶醫學 2022年3期
李志民,曹 興,余巨明,蔣國會
(川北醫學院附屬醫院神經內科,四川南充 637000)
癲癇是一種腦部神經元異常放電且反復發作為特征的常見腦部疾病[1],目前還沒有太多辦法能夠抑制癲癇的發生。顱內感染作為癲癇的重要病因[2],抑制顱內感染炎性環境從而抑制癲癇的發生無疑是研究的方向。雖然抗感染治療能降低腦炎病死率,但不能改變腦炎后形成癲癇的結果。目前亦推薦預防性抗癲癇治療[3-5]。盡管顱內感染是如何導致癲癇發作和癲癇形成的具體機制迄今還不十分清楚,但推測可能與促炎細胞因子[白細胞介素(IL)-1β、 腫瘤壞死因子(TNF)-α]、抗炎細胞因子(IL-10)失衡,膠質細胞異常激活及神經元的變性壞死共同導致腦神經網絡引起異常興奮有關[6-7]。
糖原合成激酶-3β (GSK-3β)是炎癥TLR4信號通路的關鍵調節器,抑制GSK-3β可以達到明顯抗炎作用,而激活GSK-3β則具有明顯促炎作用[8],因而對維持機體炎癥平衡起到至關重要的作用。它作為絲氨酸/蘇氨酸蛋白激酶功能并不單一。GSK-3β通過絲氨酸/蘇氨酸蛋白激酶去磷酸化激活可導致細胞凋亡,然而通過促進絲氨酸/蘇氨酸蛋白激酶磷酸化(P -GSK-3β)而達到細胞保護的作用,均通過pI3K/Akt通路影響GSK-3β的活性[9]。Wortmannin(WT)抑制pI3K使絲氨酸/蘇氨酸蛋白激酶去磷酸化而激活[10],因此被視為GSK-3β的激動劑。氯化鋰(LiCl)和丙戊酸(VPA)能使GSK-3β磷酸化失活,故被視為GSK-3β的抑制劑。LiCl在精神疾病中發揮了重要作用,目前研究其在神經系統疾病如腦和脊髓損傷[11]及神經變性疾病如帕金森病[12]的治療也發揮一定作用。然而,LiCl對大鼠顱內感染及顱內感染后癲癇發作的影響還鮮見報道。本實驗發現LiCl對癲癇發作的影響可能與劑量相關,小劑量LiCl(小于40 mg/kg)可能抑制癲癇發作,而大劑量LiCl(大于60 mg/kg)可促進癲癇的發作[13]。本課題組前期實驗得出LiCl對大鼠顱內感染后癲癇發作的影響具有劑量依賴效應,但具體機制尚不清楚。因此本研究采用不同水平的LiCl、VPA及WT干預顱內感染模型大鼠,通過免疫組織化學、Western blot、ELISA、電生理檢測的方法,探討GSK-3β在LiCl影響顱內感染大鼠癲癇發作的關系,進而為顱內感染后癲癇的預防提供方向。
1 材料與方法
1.1 一般材料
選取8~10周齡約200 g的成年雄性Sprague-Dawley大鼠(均由川北醫學院實驗動物中心提供);LiCl、LPS(美國Sigma 公司)、小膠質細胞抗體 (IBA-1,日本Wako 公司)、毛果蕓香堿(Pilo,美國Sigma公司)、Wortmannin (美國Abmole公司)、抗-Phospho-GSK-3α/β抗體、抗-GSK-3α/β抗體(美國R&D公司)、IL-10、IL-1β、TNF-α檢測試劑盒(武漢博士德公司)、神經元特異性核蛋白抗體(NeuN,美國Millipore公司)、β-actin多克隆抗體(北京中杉金橋公司)。川北醫學院實驗動物倫理委員會批準該實驗。
1.2 方法
1.2.1模型制備及分組
制備脂多糖(LPS)顱內感染大鼠模型[14]:在大鼠側腦室 (前囟后0.8 mm,旁開1.3 mm,入腦 3.5 mm) 采用注射LPS 50 μg,建立大鼠顱內感染模型。對照組(5只)給予側腦室注射同體積生理鹽水5 mL/kg。觀察記錄實驗鼠行為活動,并于24 h檢測實驗鼠海馬組織中炎癥因子和GSK-3β的水平。
側腦室注射LPS誘導顱內感染大鼠105只,然后將其均分為LPS組,LPS-10、20、40、80 mg/kg LiCl組,以及LPS-VPA組和LPS-WT組。對應分組分別進行腹腔注射生理鹽水5 mL/kg,LiCl 10、20、40、80 mg/kg,以及VPA 30 mg/kg和WT 0.6μg/kg[15],LiCl<40 mg/kg時為LPS-小劑量LiCl組。每24小時1次,連續3 d。
1.2.2癲癇大鼠模型造模及其電生理檢測[16]
根據3.3 mL/kg的比例,采用水平為10%水合氯醛對大鼠進行腹腔注射麻醉,然后將其置于腦立體定位注射儀上。乙醇消毒后對大鼠進行備皮,沿顱骨中縫剪開頭皮后以Bregma十字縫為參照向左移2.6 mm,向后移3.6 mm為中點用顱骨鉆開一邊長為3 mm骨窗。在骨窗近鼻端鉆2小孔并插入固定小螺栓用于連接參考電極。參照大鼠腦圖譜,以Bregma十字縫為參照向后3.6 mm,向左2.6 mm移動 4×4微陣列排列的16導微絲電極至左側海馬CA1區并用微推進器向下刺入腦中3.5 mm,然后連接相應電極。待大鼠麻醉蘇醒后,使用16導在體電生理記錄儀(美國Plexon公司)和OmniPlex?D神經信號采集系統(美國Plexon公司)記錄大鼠海馬CA1區神經元的電活動信號。首先,采集15 min大鼠清醒狀態下的腦電信號作為基準線;然后按照360 mL/kg的劑量腹腔注射毛果蕓香堿誘發癲癇持續狀態(SE)并記錄30 min。采用腦電分析系統(NeuroExplorer?v4.0)對實驗大鼠腦電信號進行ripples振蕩和SE海馬局部場電位的FRs的功率譜密度均值。
1.2.3IL-1β、TNF-α和IL-10水平檢測
用水合氯醛(3.3 mL/kg,腹腔注射)麻醉大鼠后斷頭取腦,快速分離雙側海馬組織(冰上操作)迅速放入勻漿管制作勻漿,而后將勻漿液移入離心管,于4 ℃、 12 000 r/min離心15 min。取上清液,用Bradford 法測定蛋白水平后分裝于0.5 mL離心管。根據ELISA試劑盒說明書進行IL-1β、TNF-α和IL-10檢測。
1.2.4p-GSK-3β和GSK-3β水平檢測[16]
采用Western blot對p-GSK-3β和GSK-3β兩個靶蛋白進行定量分析。將實驗大鼠海馬區提取的蛋白質裂解液用10%的十二烷基磺酸鈉-聚丙烯酰胺凝膠電泳進行分離。然后,采用半干式轉印法將蛋白質條帶轉移至硝酸纖維素薄膜上。檢測條件:5%脫脂牛奶,室溫封閉硝酸纖維素薄膜2 h;然后,將封閉液更換為一抗溶液,分別為1∶1 000稀釋的兔抗鼠的p-GSK-3β多克隆抗體溶液、GSK-3β多克隆抗體溶液和β-actin多克隆抗體溶液,4 ℃冰箱孵育過夜。次日,回收一抗溶液,并用PBS洗滌硝酸纖維素薄膜3次,每次5 min。然后,室溫用HRP標記的羊抗兔IgG孵育1.5 h。PBS洗滌硝酸纖維素薄膜3次后,用新鮮配制的化學發光顯色劑對蛋白質條帶進行顯影。用Image J軟件測定各蛋白質條帶的灰度值,以靶蛋白與內參蛋白β-actin的灰度值之比表示靶蛋白的相對表達水平。
1.2.5神經元和小膠質細胞檢測
采用水平為10%的水合氯醛,按照3.3 mL/kg的劑量,對大鼠實施腹腔注射麻醉。將小鼠固定后用生理鹽水灌流大鼠洗滌血細胞。待右心耳流出液呈無色后再用4%多聚甲醛灌流大鼠。大鼠灌流完成后小心取出腦組織并置于4%多聚甲醛溶液中后固定24 h。將后固定好的腦組織經脫水處理后,用石蠟包埋并制作連續的冠狀組織切片(每片4 μm厚)。海馬區腦組織切片每間隔4張取1張進行免疫組織化學染色。使用小膠質細胞特異性抗體IBA-1(1∶200)檢測活化的小膠質細胞;用神經元特異性抗體NeuN(1∶1 000)檢測神經元細胞。所有組織切片均用蘇木精進行復染。
參照YANG等[17]的方法,對免疫組織化學染色的腦組織切片進行陽性細胞計數。在40倍放大的顯微鏡視野下,選取海馬CA1及鄰近皮質區中具有代表性的,非重疊區域(400 μm×400 μm)進行NeuN+和IBA-1+細胞計數。所有的細胞計數區域均由觀察者預先設定,并采用Image-Pro Plus Media Cybernetics系統對NeuN+和IBA-1+細胞自動計數。計數視野中,只有形態結構完整,細胞清晰可辨的NeuN+和IBA-1+細胞才會被計數。由于大鼠左、右大腦半球的細胞數目無明顯差別,故不特意區分左、右半球,并將Image-Pro Plus Media Cybernetics系統獲得的結果用于統計分析。
1.3 統計學處理
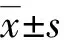
2 結 果
2.1 顱內感染大鼠模型中海馬GSK-3β和p-GSK-3β的變化
2.1.1LPS顱內感染大鼠模型
相比對照組大鼠而言,LPS組活動明顯減少、食物攝入量降低、飲水量增加、體溫上升明顯。在LPS組的海馬組織中存在明顯的促炎性細胞因子IL-1β和TNF-α的水平增高,與對照組比較差異有統計學意義(P<0.05),見圖1。
a:P<0.05,與對照組比較。
2.1.2大鼠海馬組織中GSK-3β和p-GSK-3β的表達
Western blot分析表明LPS組的大鼠海馬組織中GSK-3β水平較對照組升高(P<0.05),而p-GSK-3β水平卻明顯比對照組降低,見圖2。
2.2 不同干預對顱內感染模型大鼠各種檢測指標的影響
2.2.1炎性因子在不同干預處理組中的表達
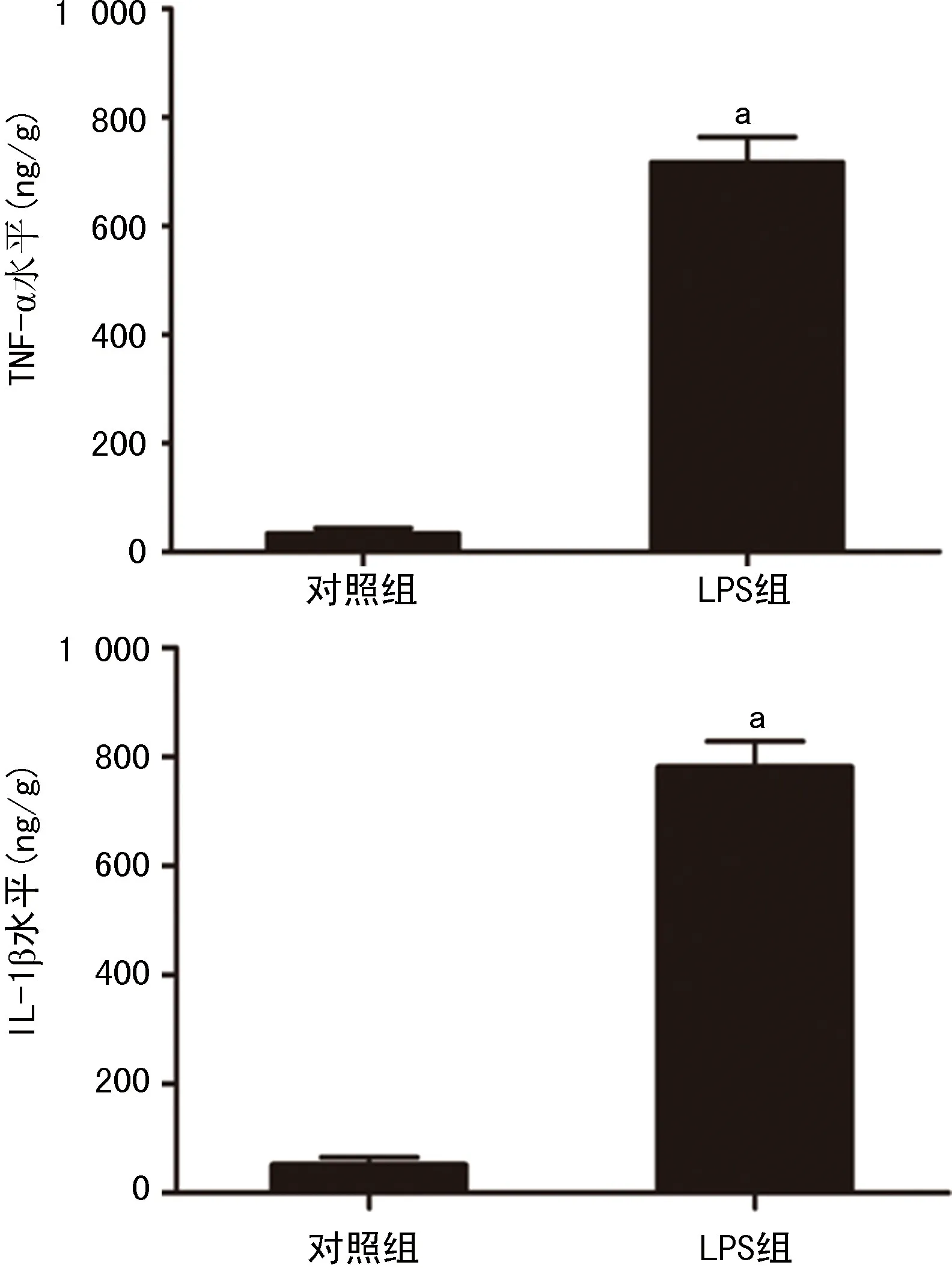
與LPS組相比LPS-小劑量LiCl組及LPS-VPA組的海馬組織中IL-10的表達水平明顯升高(P<0.05)。LPS-80 mg/kg LiCl組和LPS-WT組的海馬組織中IL-10的表達水平明顯降低(P<0.05),見圖3A。LPS-小劑量LiCl組、LPS-VPA組的海馬組織中IL-1β和TNF-α的表達水平明顯降低(P<0.05),LPS-80 mg/kg LiCl組及LPS-WT組的海馬組織中IL-1β和TNF-α的表達水平明顯升高(P<0.05),見圖3B、C。

A:Western blot檢測;B:GSK-3β相對表達水平;C:p-GSK-3β相對表達水平;a:P<0.05,與對照組比較。
A:IL-10;B:TNF-α;C:IL-1β;1:LPS組;2~5:分別為LPS-10、20、40、80 mg/kg LiCl組;6:LPS-VPA組;7:LPS-WT組;a:P<0.05,與LPS組比較。
2.2.2不同干預對顱內感染大鼠模型海馬CA1區神經元的影響
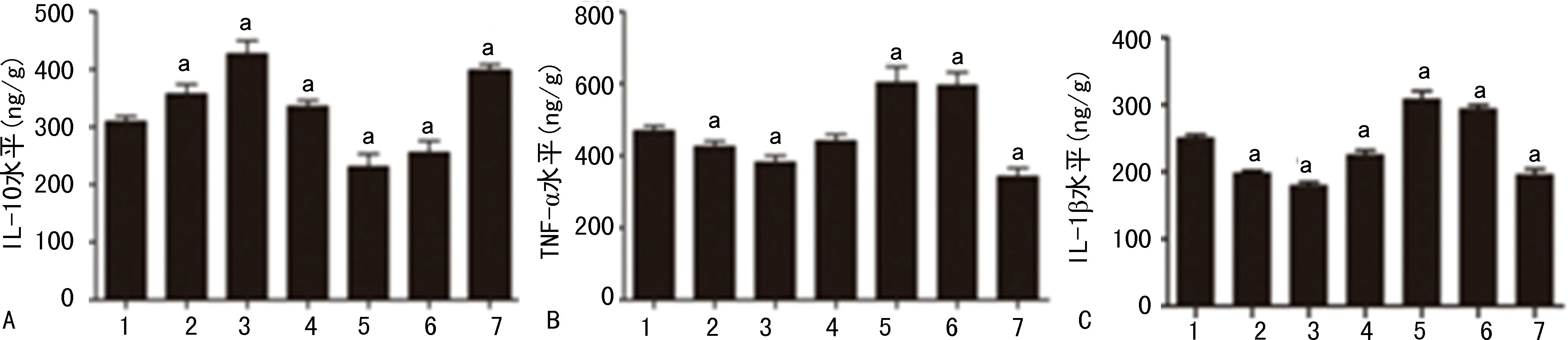
LPS-小劑量LiCl組和LPS-VPA組的海馬CA1區神經元形態結構比LPS組的更為完整,排列也更整齊而致密、細胞核脫失少、染色較深。小劑量LiCl干預組和LPS-VPA組的海馬CA1區神經元數量與LPS組相比差異有統計學意義(P<0.05)。LPS-80 mg/kg LiCl組及LPS-WT組的海馬CA1區神經元形態結構的損壞比LPS組更為嚴重,神經元排列松散、散亂,細胞核脫失嚴重、著色較淺。海馬CA1區神經元數量與LPS-80 mg/kg LiCl組、LPS-WT組和LPS組之間差異有統計學意義(P<0.05),見圖4、5。
2.2.3不同干預對顱內感染大鼠模型海馬CA1區小膠質細胞的影響
LPS-小劑量LiCl組和LPS-VPA組的海馬CA1區小膠質細胞的激活比LPS明顯要少(P<0.05);而LPS-80 mg/kg LiCl組和LPS-WT組的海馬CA1區小膠質細胞的激活比LPS組顯著增多(P<0.05),見圖6、7。
1:LPS組;2~5:分別為LPS-10、20、40、80 mg/kg LiCl組;5:LPS-VPA組;6:LPS-WT組;a:P<0.05,與LPS組比較。

A:LPS組;B~E:分別為LPS-10、20、40、80 mg/kg LiCl組;F:LPS-WT組;G:LPS-VPA組。

A:LPS組;B~E:分別為LPS-10、20、40、80 mg/kg LiCl組;F:LPS-WT組;G:LPS-VPA組。
1:LPS組;2~5:分別為LPS-10、20、40、80 mg/kg LiCl組;6:LPS-VPA組;7:LPS-WT組;a:P<0.05,與LPS組比較。
2.2.4不同干預對顱內感染模型大鼠癲癇發作的影響
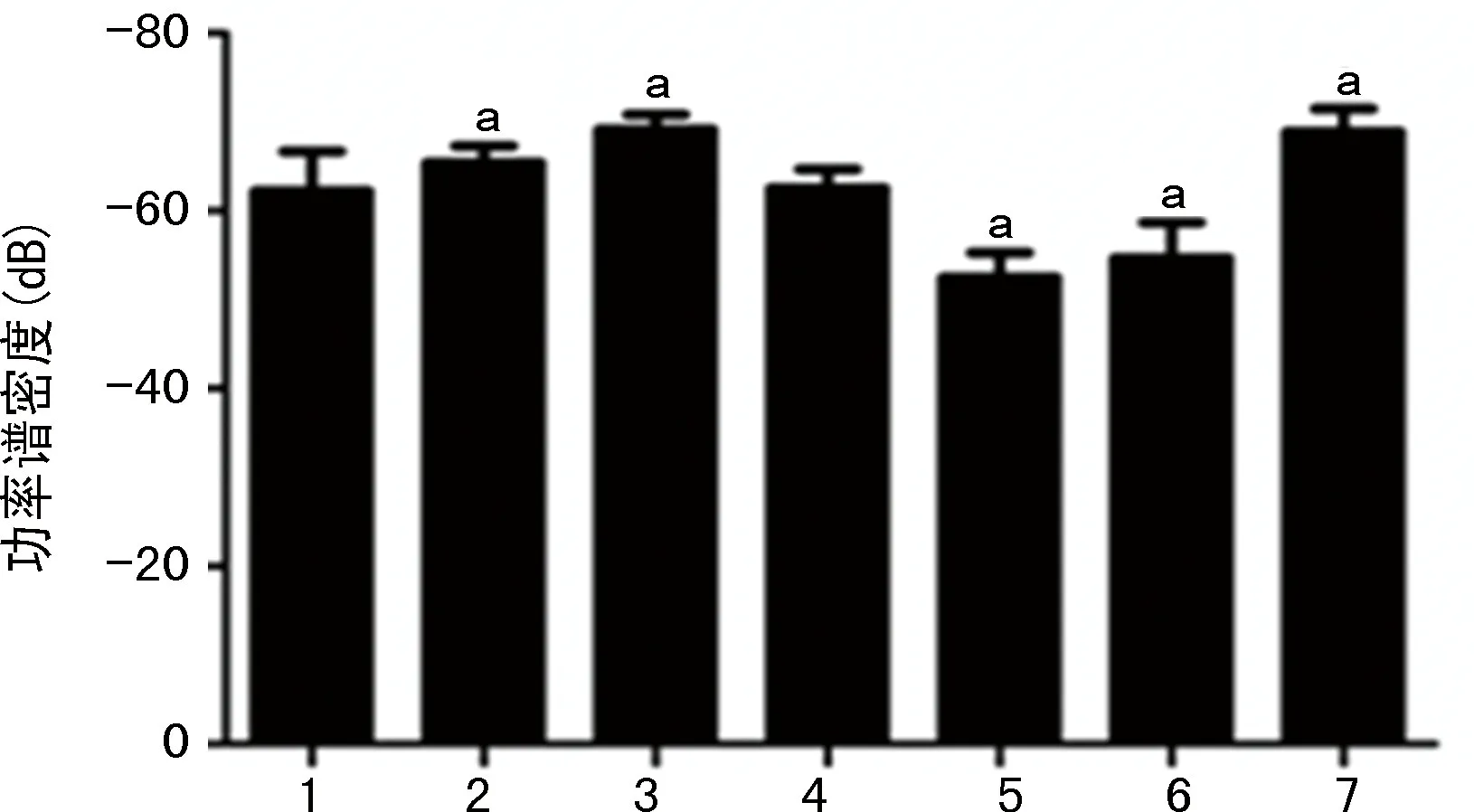
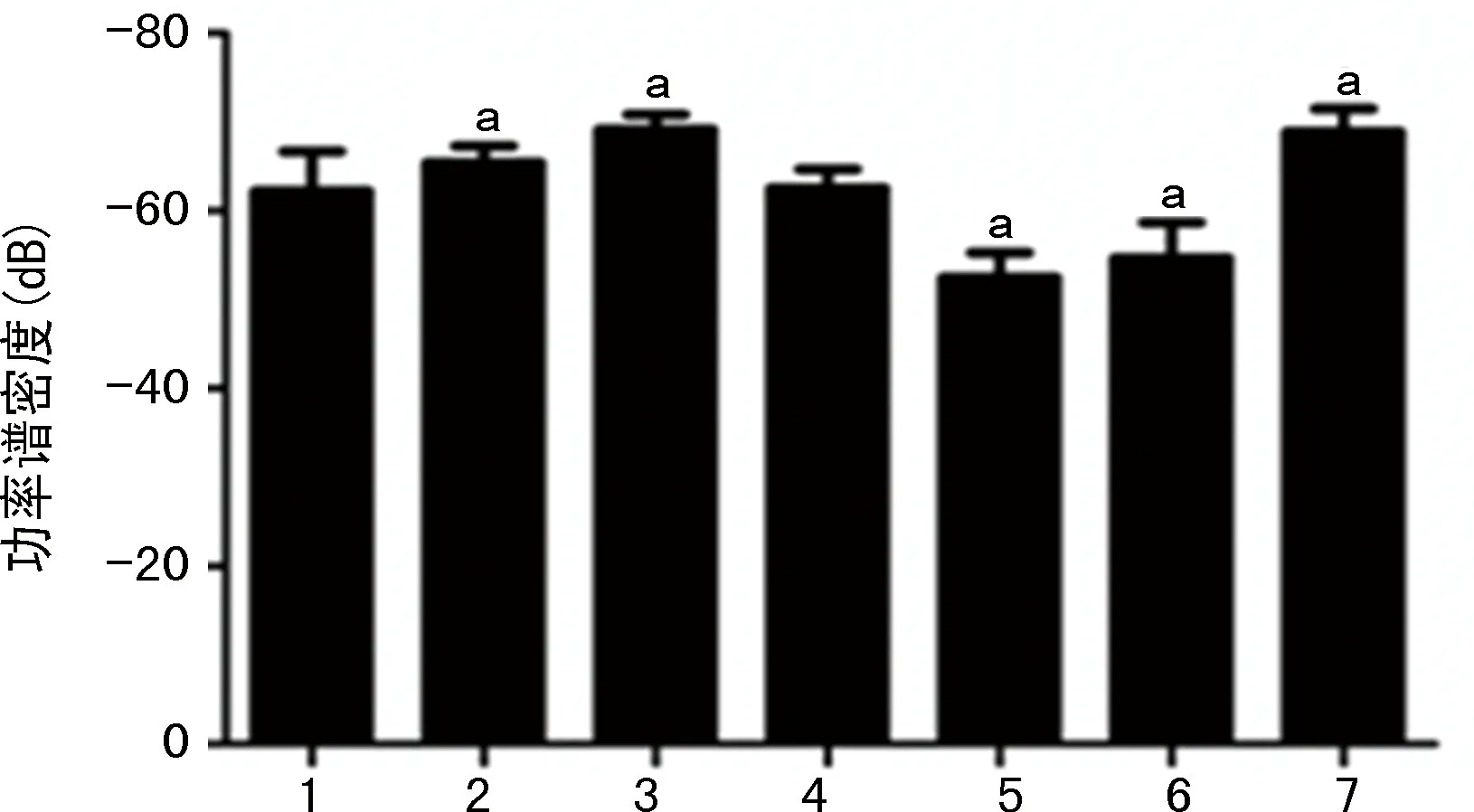
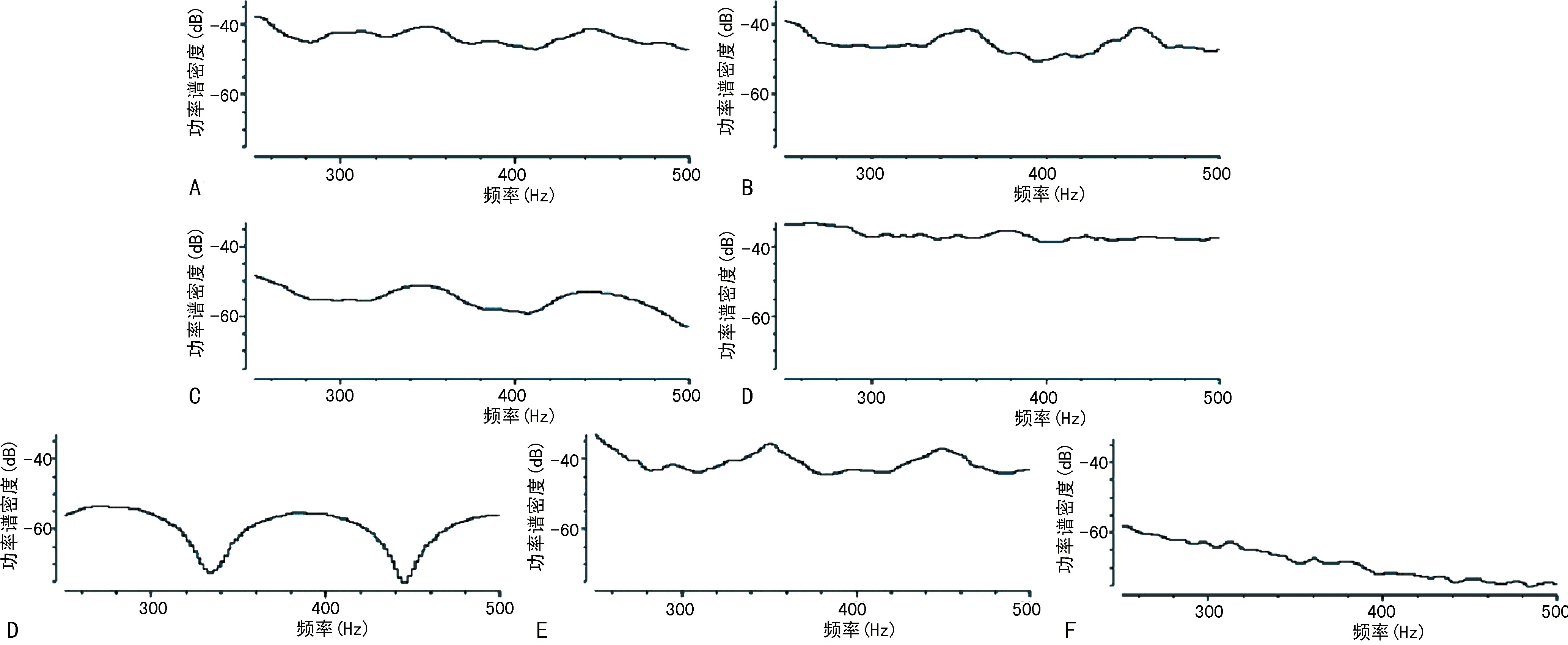
比較各干預組和LPS組的基礎腦電信號、癇性首次發作和SE潛伏期、SE期的局部場電位變化。(1)不同干預對海馬局部場電位ripples振蕩功率的影響:LPS-小劑量LiCl組和LPS-VPA組的海馬CA1區的ripples振蕩功率與LPS組相比,明顯降低(P<0.05)。LPS-80 mg/kg LiCl組和LPS-WT組的海馬CA1區的ripples振蕩功率與LPS組相比,顯著升高(P<0.05),見圖8。(2)不同干預對顱內感染模型大鼠SE潛伏期的影響:LPS-小劑量LiCl組和LPS-VPA組的SE潛伏期與LPS組相比明顯延長(P<0.05),LPS-80 mg/kg LiCl組和LPS-WT組的SE潛伏期與LPS組相比明顯縮短(P<0.05)。LPS-小劑量LiCl組和LPS-VPA組的癲癇發作頻譜能量與LPS組相比明顯降低(P<0.05),LPS-80 mg/kg LiCl組和LPS-WT組的癲癇發作頻譜能量與LPS對照組相比顯著升高(P<0.05),見圖9、10。(3)不同干預對顱內感染模型大鼠SE期FRs的功率譜密度均值的影響:LPS-小劑量LiCl組和LPS-VPA組的海馬區SE期FRs的功率譜密度均值與LPS組相比明顯降低(P<0.05),LPS-80 mg/kg LiCl干預組和LPS-WT組的海馬區SE期FRs的功率譜密度均值與LPS組相比明顯升高(P<0.05),見圖11、12。
2.2.5不同干預對顱內感染模型大鼠海馬組織中GSK-3β和p-GSK-3β的影響
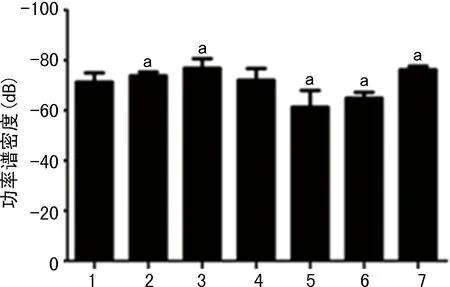
Western blot分析顯示,LPS-小劑量LiCl組和LPS-VPA組海馬組織中GSK-3β/p-GSK-3β的比值與LPS組相比明顯變小(P<0.05)。LPS-80 mg/kg LiCl組和LPS-WT組海馬組織中GSK-3β/p-GSK-3β的比值與LPS組相比明顯增大(P<0.05),見圖13。
1:LPS組;2~5:分別為LPS-10、20、40、80 mg/kg LiCl組;6:LPS- VPA組;7:LPS-WT組;a:P<0.05,與LPS組比較。

A:LPS組;B~E:分別為LPS-10、20、40、80 mg/kg LiCl組;F:LPS-WT組;G:LPS-VPA組。深紅色箭頭指示了腹腔注射毛果蕓香堿的起始時間,黑色箭頭指示了模型大鼠進入SE的起始時間。癲癇發作頻譜圖的橫坐標代表時間(s),縱坐標代表癲癇發作頻率(Hz),左側彩色條帶為頻譜能量示例,頻譜能量較高的以紅色表示,頻譜能量較低的則以藍色表示。

1:LPS組;2~5:分別為LPS-10、20、40、80 mg/kg LiCl組;6:LPS-VPA組;7:LPS-WT組;a:P<0.05,與LPS組比較。
1:LPS組;2~5:分別為LPS-10、20、40、80 mg/kg LiCl組;6:LPS- VPA組;7:LPS-WT組;a:P<0.05,與LPS組比較。
A:LPS組;B~E:分別為LPS-10、20、40、80 mg/kg LiCl干預組;F:LPS-WT組;G:LPS-VPA組。

1:LPS組;2~5:分別為LPS-10、20、40、80 mg/kg LiCl組;6:LPS-VPA組;7:LPS-WT組;a:P<0.05,與LPS組比較。
3 討 論
本實驗采用不同劑量的LiCl干預顱內感染大鼠,電生理研究結果海馬局部場電位(LFPs)顯示小劑量LiCl干預后延長了毛果蕓香堿誘發顱內感染大鼠癲癇發作的潛伏期,降低了SE期的快速高頻振蕩(FRs)功率譜密度均值,大劑量LiCl干預后則結果相反,由此得出小劑量LiCl降低了顱內感染后癲癇發作的易感性及減輕癲癇發作程度,LiCl對大鼠顱內感染后癲癇發作差異性影響的機制目前尚不十分清楚。盡管有證據表明感染所致的血腦屏障通透性增加、膠質細胞活化增殖、炎性介質大量釋放、神經元興奮性改變及變性壞死等一系列病理生理改變,均可能參與了顱內感染后癇性發作和癲癇形成的過程[11-13]。本實驗也得出小劑量LiCl干預后減少了顱內感染大鼠海馬小膠質細胞的活化,降低了海馬促炎細胞因子IL-1β、TNF-α水平,增加了抗炎細胞因子IL-10水平,減輕了海馬神經元的變性壞死。而大劑量LiCl干預后則結果相反。
本研究首先確定大鼠側腦室注射LPS 24 h后,大鼠活動明顯減少、食物攝入量降低、飲水量增加、體溫上升明顯,且海馬促炎細胞因子顯著增高,表明成功誘導大鼠顱內感染,并發現大鼠感染后腦內海馬GSK-3β水平有升高及p-GSK-3β水平降低變化,因此推測大鼠顱內炎癥嚴重程度可能與GSK-3β的活性有關。在LPS致大鼠顱內感染的基礎上,采用GSK-3β間接抑制劑VPA(30 mg/kg)和GSK-3β激動劑WT(0.6 μg/kg)分別干預顱內感染模型大鼠后,Western blot結果顯示VPA與小劑量LiCl干預后均使顱內感染大鼠海馬中p-GSK-3β/GSK-3β比值明顯變大(P<0.05),WT、與大劑量LiCl干預后均使顱內感染大鼠海馬p-GSK-3β/GSK-3β比值明顯變小(P<0.05),p-GSK-3β與GSK-3β 的比值大小體現GSK-3β活性的高低。因此VPA與小劑量LiCl干預后抑制了顱內感染模型大鼠海馬GSK-3β活性,WT、與大劑量LiCl干預后則增加了顱內感染模型大鼠海馬GSK-3β活性。研究證實抑制GSK-3β可有效減輕外周和中樞神經系統的炎癥[18],而激活GSK-3β可間接誘導IL-6、TNF-α的產生來促進CD11b表達而加重炎癥[19]。本實驗結果小劑量LiCl與VPA減少了海馬CA1小膠質細胞的激活,同時海馬促炎細胞因子IL-1β、TNF-α分泌減少,抗炎細胞因子IL-10分泌增加,而大劑量LiCl與WT干預后則得出相反的結果,YU等[20]發現LiCl通過促進GSK-3β的磷酸化而抑制GSK-3β的活性來實現抑制神經炎癥作用。本實驗表明LiCl對顱內炎癥影響的差異可能是通過調節GSK-3β的活性來實現的。
LiCl和VPA作為膜穩定劑,對多種中樞神經系統疾病具有抗炎及神經保護作用,LiCl通過調節GSK-3β的活性來直接影響顱內感染后癲癇的發作,還是通過調節GSK-3β的活性來調控顱內炎癥從而間接影響顱內感染后癲癇的發作,目前尚不清楚。但LiCl在其他神經系統疾病的研究中發現,LiCl主要通過抑制GSK-3β的活性來發揮神經保護。眾多研究發現通過調節GSK-3β途徑亦發揮神經保護作用[21-22]。本實驗和其他研究均發現LPS誘發顱內感染大鼠的海馬總GSK-3β水平增加[23]。LPS誘發顱內感染大鼠癲癇發作的敏感性顯著增加。推測GSK-3β可能與癲癇發作的敏感性有關。本實驗發現小劑量LiCl與VPA干預后明顯延長了顱內感染后SE潛伏期,而大劑量LiCl與WT干預后明顯縮短了顱內感染后SE潛伏期,這種結果可能與LiCl參與調節GSK-3β的活性有關。研究表明毛果蕓香堿誘導癲癇發作后海馬的GSK-3β的水平增加,因此GSK-3β與癲癇的發作可能直接相關[24]。 本研究利用的GSK-3β抑制劑VPA干預,結果與小劑量LiCl干預相同,均降低了癇性發作的敏感性。而利用GSK-3β的激動劑,結果與小劑量LiCl干預結果相反,與大劑量LiCl干預結果相同,增加了癇性發作的敏感性。本實驗結果顯示LPS誘發的顱內感染大鼠海馬的GSK-3β水平增高,p-GSK-3β的水平降低。GREEN等[23]用LPS刺激體外培養的星形膠質細胞和小膠細胞發現細胞GSK-3β水平較未刺激組明顯增加,而p-GSK-3β水平較未刺激組明顯減少,因而證明了炎癥提高了GSK-3β的表達活性,同樣用LiCl和LPS同時干預發現LPS-LiCl組較LPS組GSK-3β的表達活性明顯降低,而p-GSK-3β的表達活性相反,表明GSK-3β與炎癥直接相關。然而在紅藻氨酸誘導癲癇模型的研究中敲出多巴胺D2受體(D2R)基因的小鼠,意外發現其海馬中 GSK-3β顯著活化,增加了紅藻氨酸對小鼠的興奮毒性,促進了海馬神經元的凋亡,增加了小鼠對紅藻氨酸誘發癲癇的敏感性;然而使GSK-3β磷酸化后,明顯減輕了紅藻氨酸對海馬神經元興奮毒性,減少了海馬細胞的凋亡和壞死[25]。提示GSK-3β通過影響海馬神經元興奮毒性及變性壞死導致癲癇發作。然而本研究發現小劑量LiCl和VPA干預后海馬局部場電位功率譜密度值ripples振蕩功率譜較LPS組明顯降低,且海馬神經元凋亡和壞死較少,而大劑量LiCl和WT干預則出現相反的結果。海馬局部場電位ripples振蕩功率譜可能與癲癇發作相關[26],其功率譜密度值的高低則代表作海馬神經元的興奮性。因此小劑量LiCl降低了顱內感染大鼠海馬神經元興奮性,而大劑量LiCl和WT則作用相反。似乎也驗證了通過調節GSK-3β活性而影響神經網絡的興奮性。雖然有報道紅藻氨酸誘發癲癇24 h后海馬中的GSK-3β的活化明顯[27]。但也有報道稱顳葉癲癇患者海馬組織中的p-GSK-3β明顯高于沒有癲癇的患者,但GSK-3β水平輕微增高[28]。GSK-3β僅是WNT信號通路的一部分,有研究發現癲癇后GSK-3β的活化改變可獨立于WNT途徑[25];也有學者認為GSK-3β是通過多因素多途徑共同作用于興奮性神經遞質如谷氨酸釋放影響癲癇發作[28]。近年研究發現洛伐他汀通過抑制GSK-3β的活性,成功的減少了癲癇大鼠海馬齒狀回(DG)苔蘚纖維發芽(MFS)[24],DG中的MFS是反復自發性發作的發作頻率和顳葉癲癇的嚴重性高度相關的重要指標[29-31]。因此抑制GSK-3β的活性可以減輕癲癇的發作程度和腦損傷。已有研究報道FRs可作為癲癇發作的嚴重程度的標志[32-34],且與神經元變性丟失相關[35]。本實驗發現LPS-小劑量LiCl組及LPS-VPA組SE期FRs的功率譜密度均值較LPS組比較明顯降低(P<0.05),因此可以得出小劑量LiCl及VPA減輕了顱內感染模型大鼠的癲癇發作程度和神經元的變性丟失。因此GSK-3β并非直接影響LiCl對顱內感染后癲癇的發作。而是小劑量LiCl抑制顱內感染大鼠海馬組織中GSK-3β的表達,抑制炎癥因子,減輕組織病理損傷,延長腦炎后癲癇放電潛伏期,降低海馬神經元興奮性,提高癲癇發作閾值,降低腦炎后癲癇發作的敏感性,減輕癲癇發作程度;大劑量LiCl則結果相反。提示GSK-3β可能參與LiCl修飾顱內感染后癲癇發作的機制。
綜上所述,LiCl可能通過影響GSK-3β活性,從而影響膠質細胞的活化和炎性細胞因子的分泌,進一步影響神經元的變性,最后影響神經元的異常放電,達到影響顱內感染后癲癇的發作。因此調控GSK-3β的活性可望成為改善顱內感染后癲癇的發作治療靶點。
猜你喜歡
課堂內外·初中版(科學少年)(2023年10期)2023-12-10 00:43:06
全科護理(2022年10期)2022-12-26 21:19:15
作文周刊·小學二年級版(2022年20期)2022-05-05 01:33:06
中國民間療法(2021年5期)2021-06-09 09:21:04
國際放射醫學核醫學雜志(2021年10期)2021-02-28 08:41:58
創新作文(小學版)(2019年10期)2019-09-25 08:12:28
飲食科學(2017年5期)2017-05-20 17:11:53
小學生學習指導(低年級)(2017年5期)2017-05-04 04:14:38
作文與考試·小學高年級版(2015年17期)2015-05-30 10:48:04
西南軍醫(2015年4期)2015-01-23 01:19:30
