桃酵母雙雜交cDNA文庫的構建及生長素響應因子4互作蛋白的篩選
2022-03-10 03:38:44郭振華要宏陽楊海清刁冬慧劉悅萍
北京農學院學報 2022年1期
郭振華,要宏陽,楊海清,王 青 ,刁冬慧,劉悅萍*
(1.北京農學院生物與資源環境學院/農業農村部華北都市農業重點實驗室,北京 102206;2.北京市平谷區人民政府果品辦公室,北京101200)
桃(PrunuspersicaL.)屬于薔薇科李屬植物,是世界第三大溫帶果樹,起源于中國[1],有著 4 000 多年的栽培歷史,在中國各省區廣泛栽培,資源十分豐富。北京市平谷區是中國最大的桃生產基地,桃園面積約7 000 hm2,2020年桃產量約18.33萬t。桃產值收入占農民收入比重大,桃產業的發展已成為平谷區果品產業提質升級的主動力,而桃的品質是影響產業發展的關鍵因素。桃的形狀、單果質量、果實大小和著色等外觀品質以及風味、香氣、糖酸比和貯運特性等內在品質決定桃的經濟價值[2],這些品質性狀的形成是由環境及遺傳等多種因素決定的,眾多代謝相關基因及調控因子參與桃品質形成過程,通過桃基因組的深測序已挖掘出一些控制果實品質性狀的基因[3]。
生長素是植物生長過程中的重要激素[4],不僅參與植物向性生長、組織器官形成等活動[5-6],還對果實的發育成熟起到重要的調控作用[7-9]。研究表明,內源生長素隨果實成熟含量逐漸降低,果實出現軟化、花青苷積累等成熟性狀[10-11]。施加較高濃度的外源生長素可以延遲果實成熟,因此生長素通常被認為是果實成熟過程的負調控信號[12-15]。在溶質性桃果實中,內源生長素含量隨果實成熟急劇上升。生長素通過介導其受體TIR1/AFB(Transport inhibitor response 1/Auxin signaling F-box)家族的F-box蛋白和Aux/IAA(Auxin/Indole-3-acetic acid)轉錄抑制因子結合, SCFTIR1/AFBE3泛素連接酶復合物將活化的泛素轉移到Aux/IAA上, Aux/IAA的多泛素化導致其通過26S蛋白酶體降解,將下游的轉錄因子ARF(Auxin Response Factor)去抑制化,從而激活生長素信號通路,使得植物最終表現出生理反應[8]。桃優質生態安全研究課題組在桃基因組中篩選出17個ARFs編碼基因,其中大部分ARF因子在桃的不同組織部位廣泛存在,個別ARF因子的表達部位具有特異性,如PpARF13和PpARF16在根與莖中沒有表達,PpARF5僅在果實中表達,PpARF4在桃果實成熟期的表達量呈現上升,外源生長素處理促進該基因在轉錄水平的表達[16]。
酵母雙雜交系統能在生物體內測定蛋白的相互作用,具有較高的靈敏性,被廣泛應用到蛋白互作的研究中。假陽性較高是這項技術的明顯缺點[17],目前通過改造Y2H Gold菌株可適當降低假陽性,另外采用 4 個報告基因(AUR1-C、ADE2、HIS3和MEL1)和新型穩定的抗生素AbAi,可以有效殺死非抗性克隆,從而有效降低背景克隆的生長[18]。目前酵母雙雜交技術已成為檢測已知蛋白之間相互作用的重要試驗手段。隨著科學研究的發展,更重要的是要發現與已知蛋白相互作用的未知蛋白,因此需建立高質量的酵母雙雜交cDNA文庫(以下簡稱cDNA文庫)用于未知蛋白的篩選。有報告通過cDNA文庫,在草莓果實發育成熟研究中獲得目標因子,FvMAP4K1是擬南芥AtSIK1的同源基因,屬于絲氨酸/蘇氨酸蛋白激酶,其通過細胞增殖和細胞膨大調控器官大小,長莢果變小,種子數量減少,推測其可能參與草莓的果實生長發育,通過cDNA文庫篩選,獲得4個可能與果實發育相關的蛋白因子[19]。在桃果實發育研究領域,PrupeSEP1在調節果實成熟軟化過程中起重要作用,通過文庫篩選,獲得與SEP1互作的蛋白因子AKR2(Aldo-Keto Reductase,醛酮還原酶)[20]。
為研究生長素響應因子對桃果實成熟的調控機制,該試驗把硬溶質桃果實成熟前后4個時期的果皮材料等量混合,利用Gateway技術構建cDNA文庫,對生長素響應因子PpARF4進行文庫篩選,共獲得26個互作蛋白。
1 材料與方法
1.1 材 料
以硬溶質桃‘突圍’為試材, 采摘于北京市平谷區中胡家務村果園,取盛花期后 75 d(第2次快速生長期,S3)、盛花期后85 d(成熟前期,S4-1)、盛花期后93 d(成熟后期,S4-2)和盛花期后101 d(完全成熟期,S4-3)的桃果實。采摘時選取長勢一致,大小相似,沒有明顯病蟲害和機械損傷的果實樣品,用蘸有清水脫脂棉將未離體的果實輕輕擦拭2~3次,去除桃外果皮的桃毛。待水漬自然風干后,摘取果實,迅速分離外果皮和中果皮,將二者切成小塊用錫紙包裹凍于液氮中。
1.2 方 法
1.2.1 桃果實總RNA的提取及雙鏈cDNA的合成 收集4個時期桃的中果皮和外果皮樣品,混合后液氮研磨成粉末,利用Total RNA提取總RNA,利用Oligotex mRNA Midi Kit對mRNA進行分離純化。使用CloneMiner逆轉錄合成雙鏈cDNA。將 cDNA 產物-20 ℃臨時保存,于-80 ℃長期保存,用于cDNA文庫的構建。
1.2.2 cDNA文庫的構建及擴增 參照Clone Miner說明書合成cDNA第1鏈和 cDNA第2鏈,用104 μL的DEPC水反復吹打30~40次,溶解cDNA,獲得雙鏈cDNA,將cDNA與三框attB1重組接頭連接(3種接頭分別各連接1份)。對cDNA進行分級分離和收集,將雙鏈cDNA與pDONR 222載體通過BP反應進行連接,轉入大腸桿菌DH10B感受態細胞中,獲得初級文庫菌液。取轉化后大腸桿菌原液10 μL,稀釋100倍,吸取50 μL 稀釋液涂布LB培養基(含Kan+)進行總庫容量鑒定。隨機挑取24個單克隆進行菌落PCR鑒定,對其插入片段長度和重組率進行鑒定。庫容量(CFU/mL)=(培養基上的克隆數/50 μL)×稀釋倍數(1×103μL)。總庫容量(CFU)=庫容量(CFU/mL)×文庫菌液總體積(mL)。吸取1 mL次級文庫菌液,加入100 mL含有Amp+的肉湯培養液中,置于30 ℃搖床, 220 r/min振蕩培養過夜;將100 mL擴增的次級文庫菌液,利用質粒抽提試劑盒進行抽提,予以備用。
1.2.3 cDNA文庫質量與滴度檢測 按照Microtube配制反應液,隨機挑取24個克隆進行菌落PCR鑒定,在Thermal Cycler PCR儀上進行PCR反應,1% Agarose膠電泳鑒定。
1.2.4 pGADT7-PpARF4誘導表達載體的構建及自激活檢測 通過PCR反應擴增PpARF4基因,根據該基因的功能區設計引物:上游引物PpARF4-F是cgccatatgATGGAAATTGATCTGAACC,下游引物PpARF4-R是gcgtcgacTTAGACCCTGATTACTGTTGG。利用無縫克隆連接載體pGADT7,經酶切鑒定,測序,成功構建PpARF4-pGADT7。將目的基因與pGADT7載體連接后轉化到大腸桿菌后獲得PpARF4-pGADT7重組質粒。將PpARF4-pGADT7重組質粒轉入Y2H gold酵母感受態細胞中,在SD/-Leu-Ade-His培養基中檢測PpARF4轉錄因子的自激活能力。
1.2.5 以PpARF4為誘餌篩選cDNA文庫 采用Mating法將含有PpARF4-pGADT7重組質粒的Y2H gold酵母細胞與cDNA文庫共培養,形成結合子。在含有X-α-gal的SD/-Leu -Ura-Ade-His培養基中篩選酵母菌株。然后將酵母菌株進行菌落PCR試驗,選取其中的陽性克隆進行測序。
1.2.6 序列分析 在NCBI中通過Blast的方法獲得基因的全長序列,最后以GDR數據庫為依據對基因功能進行注釋。
2 結果與分析
2.1 果實總RNA的提取及ds cDNA的合成
為建立桃果實成熟前后的cDNA文庫,收集‘突圍’桃的中果皮和外果皮樣品,用液氮研磨成粉末,利用Total RNA試劑盒提取總RNA,利用Oligotex mRNA Midi Kit對mRNA進行分離純化,使用CloneMiner逆轉錄合成雙鏈cDNA,用于后續的建庫(圖1)。

注:M是marker;1是桃中果皮;2是桃外果皮。Note:M is marker; 1 is mesocarp of peach fruit; 2 is exocarp of peach fruit.圖1 桃果實總RNA的提取Fig.1 RNA extraction from peach fruit
2.2 cDNA文庫質量和滴度檢測
首先進行BP重組,將桃的cDNA與pDONR222共轉化大腸桿菌,培養后獲得初級文庫菌液。將初級文庫菌液稀釋100倍,然后取50 uL涂于LB固體培養基,培養后第2天得到初級文庫總庫容量1.04×106CFU(圖2A),隨機選取24個克隆進行菌落PCR檢測,電泳結果如圖2B,片段大小差異較為明顯,主要分布在1 000~2 000 bp,重組率為96%,覆蓋率較高,達到初級文庫要求。
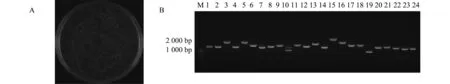
注:A是初級文庫總庫容量鑒定;B是初級文庫插入片段PCR鑒定;M是marker;1-24是挑選的克隆。Note:A is the capacity of primary library; B is PCR identification of inserted fragments in the primary library; M is Marker; 1-24 is selected colonies.圖2 初級文庫總庫容量及插入片段PCR鑒定Fig.2 Identification of the capacity and PCR of inserted fragments in the primary library
抽提初級文庫質粒,與PGADT7-DEST載體進行LR重組,轉化大腸桿菌DH10B,培養后得到次級文庫菌液,將菌液稀釋100倍,取50 uL涂于LB固體培養基,獲得總庫容量1.6×107CFU(圖3 A)。隨機選取24個克隆進行PCR檢測,主要集中在1 000~2 000 bp,重組率為100%,電泳結果如圖3 B,所獲結果滿足次級文庫要求,可進行后續cDNA文庫篩選試驗。
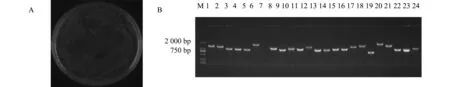
注:A是次級文庫總庫容量鑒定;B是次級文庫插入片段PCR鑒定;M是Marker;1-24是挑選的克隆。Note:A is the capacity of secondary library; B is PCR identification of inserted fragments in the secondary library; M is marker; 1-24 is selected colonies.圖3 次級文庫總庫容量及插入片段PCR鑒定Fig.3 Identification of the capacity and PCR of inserted fragments in the secondary library
2.3 pGBKT7-PpARF4誘餌載體的構建與自激活檢測
通過PCR反應成功擴增PpARF4基因。對目的基因和pGBKADT7質粒進行連接,并轉化至大腸桿菌中,通過菌落PCR篩選,再經過雙酶切檢驗,將構建成功的pGBKT7-PpARF4重組質粒轉入AH109酵母菌株中,置于SD/-Trp固體培養基上培養,然后對單菌落重懸,采用GAL4系統檢測自激活,在添加X-gal試劑的SD/-Trp/-Ura-Ade/X-gal/AbAi缺陷型培養基中未長出藍色酵母菌株,證明PpARF4誘餌蛋白不存在自激活(圖4),可進行下一步試驗。
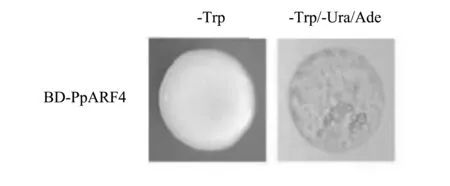
圖4 pGBKT7-PpARF4誘餌蛋白自激活檢測Fig.4 Self-activation assay of pGBKT7-PpARF4 bait protein
2.4 酵母雙雜交篩選PpARF4的互作蛋白
將構建好的pGBKT7- PpARF4 質粒和桃果實cDNA文庫質粒共轉化Y2H Gold 酵母感受態細胞,稀釋100倍涂布于SD/-Trp/-Leu培養基上,培養4 d長出菌落,將篩選得到的陽性克隆再次轉移到SD/-Trp/-Ura-Ade/X-gal/AbAi選擇性培養基上,得到48個陽性克隆(圖5),選取陽性克隆進行PCR鑒定,最終得到測序成功的陽性克隆共26個。
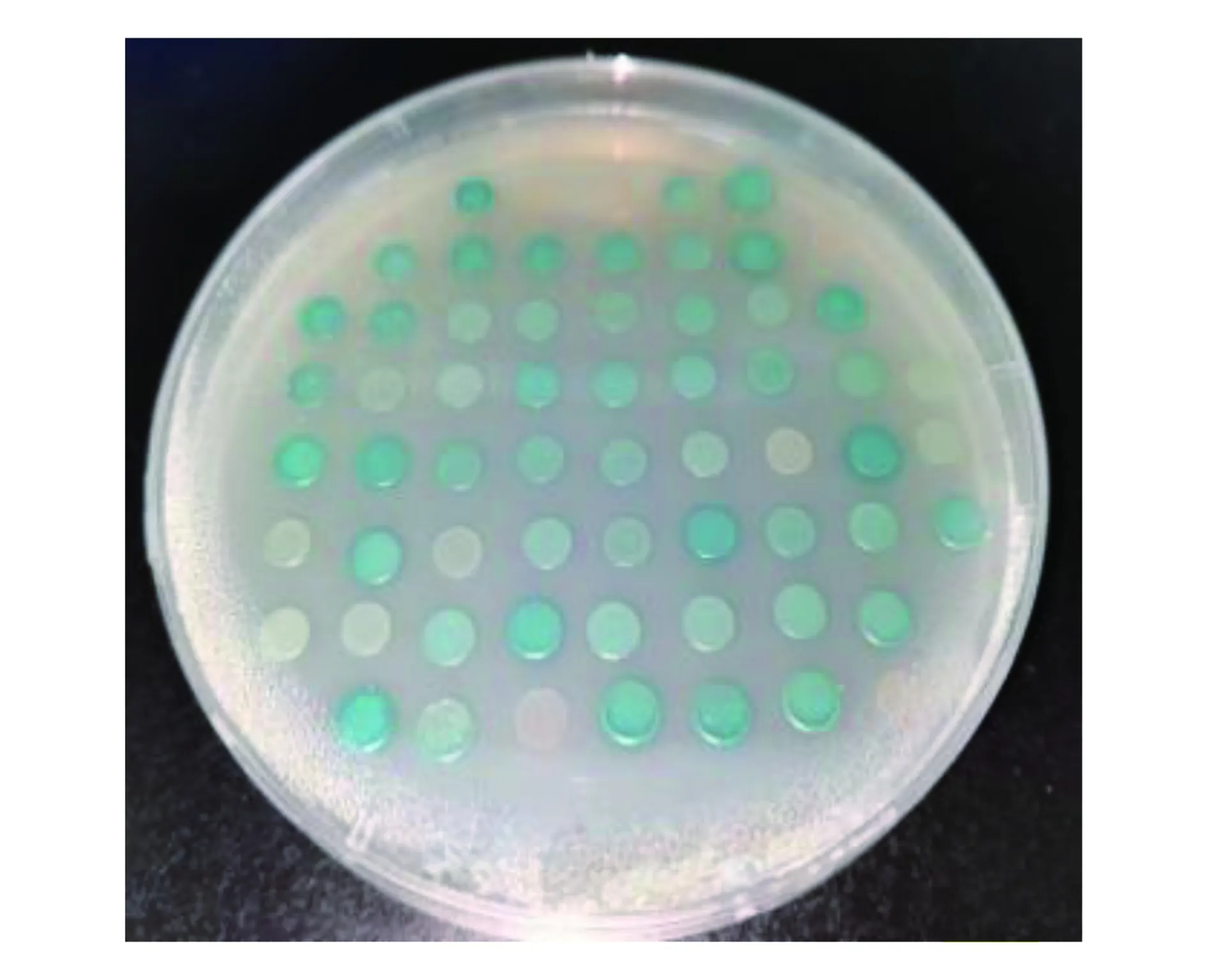
圖5 PpARF4蛋白cDNA文庫篩選獲得的克隆Fig.5 Colonies obtained by PpARF4 screening in cDNA library
2.5 互作蛋白的功能注釋
利用酵母質粒提取試劑盒,提取測序成功的陽性克隆的酵母質粒。將酵母質粒轉入大腸桿菌,過夜培養,送公司測序。測序后得到26個蛋白,在NCBI上的信息如表1所示,獲得的鑒定蛋白功能涉及植物激素信號轉導相關蛋白、脅迫應答因子、物質代謝和形態建成等因子。通過該試驗建立的cDNA文庫,篩選獲得的PpARF4互作蛋白,為進一步研究桃果實發育成熟機制提供了有價值的信息。
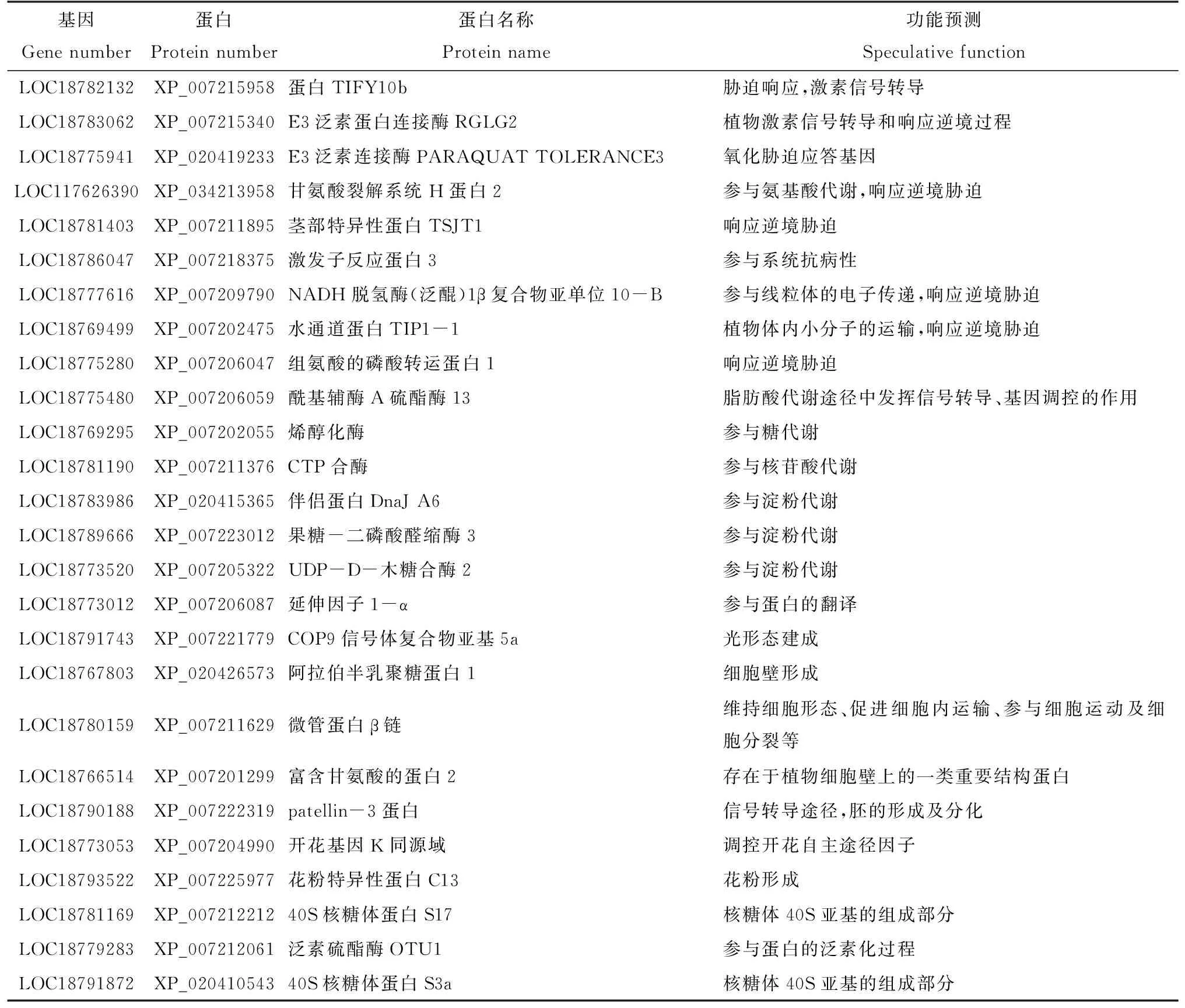
表1 篩選獲得的PpARF4互作蛋白及其功能預測Tab.1 The proteins interacting with PpARF4 by screening and its functional prediction
3 討 論
桃果實的發育成熟過程是由多因子調控的生理代謝過程,該試驗用桃果實成熟前后果皮樣品成功構建cDNA文庫,通過篩選,有利于獲得與果實成熟相關的目標蛋白因子。構建高質量的cDNA文庫是利用酵母雙雜交技術篩選互作蛋白的重要保證,該試驗構建的cDNA文庫有較高的重組率(100%),較長的插入片段(1 000~2 000 bp)和較大的總庫容量(1.6×107CFU),屬于高質量的cDNA文庫,為后續篩選與桃果實成熟相關的靶蛋白提供重要的試驗體系。
轉錄因子ARF是響應生長素信號的關鍵反應因子,在桃果實上存在17個PpARFs基因[21],在果實的成熟過程中PpARF4基因的表達量下調,是桃果實成熟過程中果皮花色苷積累的負調控因子[22],而PpARF4調控的靶基因及其互作的蛋白因子是探究PpARF4功能的關鍵。該研究利用構建好的cDNA文庫,成功篩選到誘餌蛋白PpARF4的26個潛在互作蛋白。其中響應逆境脅迫或植物激素信號轉導相關蛋白有9個,分別是蛋白TIFY10b,E3泛素蛋白連接酶RGLG2,E3泛素連接酶PARAQUAT TOLERANCE 3,甘氨酸裂解系統H蛋白2,莖部特異性蛋白TSJT1,激發子反應蛋白3,NADH脫氫酶(泛醌)1β復合物亞單位10-B,水通道蛋白TIP1-1和組氨酸的磷酸轉運蛋白1;與大分子代謝相關的蛋白7個,分別是酰基輔酶A硫酯酶13,烯醇化酶,CTP合酶,葉綠素體內的伴侶蛋白dnaJ A6,果糖-二磷酸醛縮酶3,UDP-D-木糖合酶2和延伸因子1-α;參與組織器官形成的蛋白有7個,包括patellin-3蛋白,開花基因K同源域,COP9信號體復合物亞基5a,阿拉伯半乳聚糖蛋白1,微管蛋白β鏈,富含甘氨酸的蛋白2和花粉特異性蛋白C13;參與蛋白合成和修飾的因子有3個,分別是40S核糖體蛋白S17、40S核糖體蛋白S3a和泛素硫酯酶OTU1。目前獲得的26個互作蛋白中沒有發現與桃果實花青苷代謝相關的因子,可能的原因是該試驗構建的誘餌蛋白載體時選取的是PpARF4的功能區,不是完整的編碼區序列,對于一些互作較弱的蛋白沒有檢測到。該試驗發現與花青苷形成的光信號因子COP9信號體復合物亞基5a,并獲得一些與糖代謝及細胞壁形成相關的蛋白,這為進一步探究PpARF4在桃果實品質形成中的可能作用提供了重要的信息。
4 結 論
建立了高質量的桃果實cDNA文庫,初步篩選出與生長素響應因子PpARF4互作的蛋白26個。
