動物源金黃色葡萄球菌生物被膜形成能力與分子分型關(guān)系研究
2022-03-04 06:09:38唐子云胡健欣陳進(jìn)陸毅興孔伶俐刁露張發(fā)福熊文廣曾振靈
中國農(nóng)業(yè)科學(xué) 2022年3期
唐子云,胡健欣,陳進(jìn),陸毅興,孔伶俐,刁露,張發(fā)福,熊文廣,曾振靈
動物源金黃色葡萄球菌生物被膜形成能力與分子分型關(guān)系研究
唐子云,胡健欣,陳進(jìn),陸毅興,孔伶俐,刁露,張發(fā)福,熊文廣,曾振靈*
華南農(nóng)業(yè)大學(xué)獸醫(yī)學(xué)院/廣東省獸藥研制與安全評價重點實驗室/嶺南現(xiàn)代農(nóng)業(yè)科學(xué)與技術(shù)廣東省實驗室/國家獸醫(yī)微生物耐藥性風(fēng)險評估實驗室,廣州 510642
【目的】調(diào)查產(chǎn)生生物被膜的金黃色葡萄球菌流行病學(xué)特征,探究生物被膜形成能力與分子型之間的相關(guān)性,為治療動物金黃色葡萄球菌感染提供理論依據(jù)。【方法】以廣東省獸藥研制與安全評價重點實驗室保存的98株金黃色葡萄球菌為研究對象,用結(jié)晶紫半定量法測定所有菌株生物被膜形成能力,將產(chǎn)膜菌株進(jìn)行22種常見抗菌藥的最小抑菌濃度測定,通過金黃色葡萄球菌常見3種(、MLST和PFGE)分型方式進(jìn)行分子分型,并分析產(chǎn)膜能力與分子型之間的相關(guān)性,最后通過全基因組測序技術(shù)分析生物被膜產(chǎn)生菌株中耐藥基因和毒力基因攜帶狀況。【結(jié)果】結(jié)晶紫半定量結(jié)果顯示,98株金黃色葡萄球菌中能產(chǎn)生生物被膜的菌株共計23株,占23.47%。包括牛乳源22株(占比25.29%,22/87),其中14株(占比60.87%,14/22)來自浙江奶牛養(yǎng)殖場,8株(占比39.13%,8/22)來自福建奶牛養(yǎng)殖場,以及豬源1株(占比9.10%,1/11),來自廣東屠宰場,說明牛乳源金黃色葡萄球菌產(chǎn)膜潛力高于豬源;將產(chǎn)膜能力劃分為強、中、弱3個等級,23株產(chǎn)膜菌株中,其中強產(chǎn)膜菌株2株(占比8.70%,2/23),中產(chǎn)膜菌株9株(占比39.13%,9/23),以及弱產(chǎn)膜菌株12株(占比52.17%,12/23)。藥敏試驗顯示,牛乳源產(chǎn)膜菌株對所有受試抗菌藥物敏感,而豬源產(chǎn)膜菌株對青霉素、阿莫西林、頭孢噻呋、頭孢西丁、恩諾沙星、環(huán)丙沙星、克林霉素、多西環(huán)素、紅霉素、氟苯尼考、復(fù)方新諾明、泰妙菌素、替米考星13種抗菌藥均表現(xiàn)出耐藥;分型結(jié)果顯示,98株金黃色葡萄球菌共獲得8種型,產(chǎn)膜的23株金黃色葡萄球菌占其中3種:1株t2922型來自廣東豬源,14株t2119來自浙江牛乳源,8株t189來自福建牛乳源;MLST分型結(jié)果顯示,98株菌共分為9種ST型,其中6種ST型不具有產(chǎn)生生物被膜的能力,分別為ST398、ST522、ST705、ST1651、ST479和ST151,僅3種ST型的菌株具有生物被膜產(chǎn)生能力,分別為ST9、ST7和ST188。結(jié)果表明,強產(chǎn)膜菌株分子型主要為ST7-t2119,中產(chǎn)膜菌株主要為ST7-t2119和ST188-t189,弱產(chǎn)膜菌株為ST9-t2922、ST7-t2119和ST188-t189;同時結(jié)合PCA分析不同ST型組間差異性發(fā)現(xiàn),弱產(chǎn)膜菌株的ST型能與中強產(chǎn)膜菌株很好的區(qū)分開來,只有特定ST型的金黃色葡萄球菌才具有產(chǎn)生生物被膜的能力。23株產(chǎn)膜菌株P(guān)FGE分型全部成功,結(jié)果顯示,廣東、福建、浙江三省產(chǎn)膜菌株共分為3種PFGE型,且PFGE型存在地域分布特征,同一地區(qū)的分離株可能存在克隆傳播,但克隆株之間生物被膜產(chǎn)生能力有明顯差異;全基因組測序結(jié)果顯示,產(chǎn)膜菌株中耐藥基因和毒力基因根據(jù)分子型不同攜帶情況呈現(xiàn)多樣性。【結(jié)論】不同來源的金黃色葡萄球菌有不同程度產(chǎn)生生物被膜的潛力,且均攜帶多種不同的產(chǎn)膜基因,牛乳源金黃色葡萄球菌產(chǎn)膜潛力遠(yuǎn)遠(yuǎn)高于豬源;菌株能否產(chǎn)膜與ST型存在強相關(guān)性,特定的ST型如ST9、ST7和ST188更容易產(chǎn)生生物被膜。
金黃色葡萄球菌;生物被膜;耐藥性;分子分型;毒力基因
0 引言
【研究意義】金黃色葡萄球菌()作為一種自然界廣泛存在的革蘭氏陽性菌,是導(dǎo)致條件致病的人畜共患病原菌,通常定植在動物表皮和黏膜上,能引起多種組織炎癥和傷口惡化[1-2]。同時,金黃色葡萄球菌往往攜帶大量的毒力基因,如各類腸毒素基因,定植在熟肉制品、水產(chǎn)品、糧食制品中,誤食后對人體腸道產(chǎn)生破壞,導(dǎo)致惡心、嘔吐等中毒癥狀[3-5]。在人醫(yī)臨床上,金黃色葡萄球菌在引起血液感染的病原菌中常居首位[6]。在畜牧養(yǎng)殖業(yè)上,金黃色葡萄球菌不僅是引起奶牛乳房炎的主要病原菌之一,且能導(dǎo)致豬的滲出性皮炎,犬膿皮病等,該病給畜牧養(yǎng)殖業(yè)帶來巨大威脅[7-9]。【前人研究進(jìn)展】細(xì)菌生物被膜或稱細(xì)菌生物膜(bacterial biofilm,BF),是細(xì)菌黏附于特定載體表面,分泌大量多糖蛋白[10],在多糖黏附素的作用下逐漸堆積并產(chǎn)生完整的保護(hù)性外膜,成熟的外膜產(chǎn)生一段時間后,可將被包裹的菌體釋放并尋找新的黏附載體繼續(xù)形成新的包裹。定植于奶牛乳房表面的金黃色葡萄球菌一旦形成生物被膜,將成為一個巨大且源源不斷的細(xì)菌聚集輸出體,生物被膜的屏障作用導(dǎo)致藥物無法到達(dá)靶作用位點從而失去藥效,而生物被膜處于動態(tài)的溶解釋放與再生狀態(tài)[11-14],導(dǎo)致細(xì)菌源源不斷地被釋放到組織中,形成隱性感染和持續(xù)感染。而盲目改變抗菌藥種類和提高用藥劑量,只會導(dǎo)致細(xì)菌耐藥現(xiàn)狀的加劇,同時給養(yǎng)殖業(yè)造成極大威脅。【本研究切入點】本研究通過對實驗室分離保存的98株金黃色葡萄球菌進(jìn)行生物被膜形成能力檢測,對產(chǎn)膜菌株進(jìn)行最小抑菌濃度測定、分子分型以及全基因組測序,探討生物被膜形成與分子型之間的關(guān)系。【擬解決的關(guān)鍵問題】了解產(chǎn)膜的金黃色葡萄球菌耐藥情況和分子特點,分析生物被膜形成與分子型之間的相關(guān)性,為獸醫(yī)臨床使用抗菌藥治療動物金黃色葡萄球菌感染提供有效指導(dǎo)。
1 材料與方法
1.1 試驗材料
1.1.1 主要試劑及培養(yǎng)基 TaKaRa TaqTM、dNTP Mixture以及DL2000均購自寶生物工程(大連)有限公司;乙二胺四乙酸、Tris 堿、硼酸、氫氧化鈉、濃鹽酸、冰醋酸、甲醇、丙酮、無水乙醇、結(jié)晶紫等購自廣州化學(xué)試劑廠。葡萄糖、10×PBS、聚苯乙烯96孔板購于廣東凱閣生物技術(shù)有限公司。溶菌酶、溶葡萄球菌素、I酶、I酶均購自生工生物工程(上海)股份有限公司;細(xì)菌全基因組提取試劑盒購于廣州碧堯德生物科技有限公司。
金黃色葡萄球菌顯色培養(yǎng)基,高鹽甘露醇瓊脂培養(yǎng)基,7.5% NaCl 肉湯,水解酪蛋白瓊脂培養(yǎng)基(Mueller-HintonAgar),水解酪蛋白肉湯(MH Broth)購自廣東環(huán)凱微生物科技有限公司。LB 瓊脂培養(yǎng)基(Luria-Bertani Agar),LB 肉湯(Luria-Bertani Broth)購于青島海博生物有限公司。
1.1.2主要儀器 恒溫震蕩箱,電熱恒溫干燥箱,電子分析天平,-20℃冰箱,-80℃超低溫冰箱高壓滅菌鍋,微量移液器,Milli-Q 超純水儀,恒溫金屬浴電動移液槍,超凈工作臺,電熱恒溫培養(yǎng)箱,臺式高速離心機(jī),紫外分光光度計、PCR擴(kuò)增儀、脈沖場凝膠電泳儀、電泳凝膠成像系統(tǒng)、DNA濃度測定儀等。
1.1.3 抗菌藥物 阿莫西林、苯唑西林、青霉素、頭孢噻呋、頭孢西丁、慶大霉素、阿米卡星、多西環(huán)素、替加環(huán)素、氟苯尼考、紅霉素、替米考星、利福平、萬古霉素、克林霉素、泰妙菌素、環(huán)丙沙星、恩諾沙星、磷霉素、利奈唑胺、泰利唑胺、復(fù)方新諾明共22種,參照美國臨床和實驗室標(biāo)準(zhǔn)化委員會(CLSI)標(biāo)準(zhǔn)進(jìn)行抗菌藥物的配制[15]。
1.1.4 菌株來源 以廣東省獸藥研制與安全評價重點實驗室2019年間分離保存的98株動物源金黃色葡萄球菌為研究對象,菌株信息見表1,金黃色葡萄球菌標(biāo)準(zhǔn)菌株ATCC29213和耐甲氧西林金黃色葡萄球菌(MRSA)ATCC43300來自于該實驗室保存。

表1 本研究菌株信息
1.2 試驗方法
所有試驗于2019—2020年在華南農(nóng)業(yè)大學(xué)廣東省獸藥研制與安全評價重點實驗室,以及國家獸醫(yī)微生物耐藥性風(fēng)險評估實驗室開展。
1.2.1 結(jié)晶紫半定量 將受試菌株和ATCC43300劃線于高鹽甘露醇平板,37℃培養(yǎng)48 h。挑單菌落于高壓滅菌過的BHI肉湯試管(含0.25%葡萄糖)中,于37℃、180 r/min搖床振蕩過夜;在96孔板第1—11列加入199 μL BHI肉湯后,1—10每列加1 μL菌液小心混勻,第11列加入等量ATCC43300菌液作陽性對照,第12列加入200 μL BHI肉湯作為陰性對照;每株菌設(shè)8個平行,靜置培養(yǎng)24 h以便膜在底部形成;小心移除培養(yǎng)基后每列用200 μL 1×PBS吹洗,每列吹洗3次以除去浮游細(xì)菌,丟棄最后一次洗滌液并在超凈臺中吹風(fēng)晾干20 min,每列加入150 μL甲醇固定5 min,倒棄液體并加入100 μL結(jié)晶紫染色20 min;用緩水沖洗至陰性對照孔水流呈無色,加入洗脫液洗脫20 min后酶標(biāo)儀測OD595,重復(fù)3次,并計算平均值。根據(jù)所測OD值大小將生物被膜產(chǎn)生能力劃分為強中弱三個等級,規(guī)定強OD595>3,中2<OD595≤3,弱1<OD595≤2,統(tǒng)計分析結(jié)果并篩選得到產(chǎn)膜菌株。
1.2.2 最小抑菌濃度測定 依照美國臨床和實驗室標(biāo)準(zhǔn)化委員會(CLSI)標(biāo)準(zhǔn)推薦,采用瓊脂稀釋法對23株產(chǎn)膜金黃色葡萄球菌進(jìn)行最小抑菌濃度的測定,并將ATCC29213作為每組質(zhì)控菌株。根據(jù)CLSI-2019 和歐洲共同體藥敏試驗委員會(the European committee on antimicrobial susceptibility testing,EUCAST)標(biāo)準(zhǔn)進(jìn)行結(jié)果判斷[15]。
1.2.3 分子分型 采用水煮法粗提細(xì)菌基因組,挑取甘露醇平板劃線的單菌落于LB平板上培養(yǎng)生長12 h后刮取少量細(xì)菌于裝有滅菌1×TE的1.5 mL EP管中,充分混勻。于103.5℃金屬浴中加熱10 min后迅速轉(zhuǎn)移至-20℃冰箱中急凍30 min以上,再轉(zhuǎn)移至金屬浴加熱10 min后13 000×離心3 min迅速轉(zhuǎn)移至新的EP管中,并做好編號,長期保存于-20℃冰箱備用。
分型。委托北京擎科生物科技有限公司進(jìn)行引物合成,引物序列見表2。PCR擴(kuò)增步驟:配制25 μL反應(yīng)體系:10×rTaq buffer 2.5 μL,dNTP mixture 2.0 μL,上、下游引物各0.5 μL,rTaq酶0.125 μL,模板1 μL,加超純水至25 μL充分混勻;反應(yīng)程序按照{(diào)94℃×5min+[94℃×30s+Tm×30s+72℃×50s]×30+72℃×10 min}進(jìn)行擴(kuò)增后用凝膠電泳成像系統(tǒng)拍照保存。將陽性條帶的擴(kuò)增產(chǎn)物送至北京擎科生物科技有限公司進(jìn)行一代測序,測序結(jié)果剪切后提交至金黃色葡萄球菌分型網(wǎng)站(http://spa.ridom.de/ submission.shtml)比對得到結(jié)果。

表2 spa分型引物合成表
MLST分型。參考金黃色葡萄球菌MLST分型網(wǎng)站(http://saureus.mlst.net)提供的7對管家基因引物序列進(jìn)行引物合成(表3),PCR擴(kuò)增參數(shù):{94℃×5min+[94℃×60s+55×60s+72℃×60s]×30+72℃×10min}將擴(kuò)增產(chǎn)物送至北京擎科生物科技有限公司進(jìn)行一代測序,測序結(jié)果提交至MLST數(shù)據(jù)庫(http://saureus.mlst.net)比對結(jié)果。

表3 MLST分型引物合成表
PFGE分型。參照劉保光等[18]的方法將產(chǎn)膜菌株進(jìn)行PFGE分型,并將分型結(jié)果輸入到BioNumerics 6.6 軟件進(jìn)行親緣關(guān)系分析。
1.2.4 全基因組測序 按照全基因組試劑盒提供的方法改良提取金黃色葡萄球菌細(xì)菌基因組:用一次性接種環(huán)刮取一平環(huán)在LB平板上培養(yǎng)12 h的菌苔,在1 mL的1×TE中充分渦旋,加入220 μL Buffer STE、30 μL Lysozyme、5 μL RNAase A,再加入1.5 μL Lysostaphin(20 mg·mL-1),渦旋混勻后37℃水浴消化50 min;加入250 μL Buffer DL和10 μL Proteinse K輕輕顛倒混勻,70℃金屬浴50 min;加入250 μL無水乙醇充分渦旋后13 000×離心3 min,吸取上清液至裝有HiPure gDNA Micro Column的收集管中,13 000×離心3 min倒棄流出液,把柱子裝回收集管后加入500 μL GW1,10 000×離心1 min倒棄流出液,再把柱子裝回收集管后加入650 μL GW2,10 000×離心1 min倒棄流出液,把柱子裝回收集管10 000×離心3 min;把柱子裝在新的1.5 mL EP管中,在膜中央加入40 μL預(yù)熱至65℃的ddH2O放置3 min,10 000×離心1 min;丟棄結(jié)合柱,保存于-20℃。將提取的細(xì)菌基因組委托北京諾禾致源科技股份有限公司進(jìn)行建庫,并在Illumina HiSeq 2000平臺上以測序策略PE 150 bp測序。將測序結(jié)果clean reads導(dǎo)入CLC Genomics Workbench 10.1,用De Novo Assembly 進(jìn)行序列拼接,k-mer為默認(rèn)值63,將大于500 bp的contigs保留,進(jìn)行后續(xù)分析。在網(wǎng)絡(luò)數(shù)據(jù)庫https://cge.cbs.dtu.dk/services/中上傳組裝后的全基因組序列,對測序菌株中的耐藥基因和毒力基因進(jìn)行查找分析。
1.2.5 統(tǒng)計學(xué)分析 利用主成分分析法(principal component analysis,PCA)分析不同ST型組間差異性。使用Graphpad Prism 8和R 包中pheatmap對統(tǒng)計結(jié)果進(jìn)行可視化。
2 結(jié)果
2.1 生物被膜產(chǎn)生能力
根據(jù)結(jié)晶紫半定量結(jié)果,98株金黃色葡萄球菌中篩選出能產(chǎn)生生物被膜的菌株共計23株,包括牛乳源22株(產(chǎn)膜率22/87=25.29%),14株(14/22=60.87%)來自浙江奶牛養(yǎng)殖場,8株(8/22=39.13%)來自福建奶牛養(yǎng)殖場,和豬源1株(產(chǎn)膜率1/11=9.10%),來自廣東屠宰場,說明牛乳源金黃色葡萄球菌產(chǎn)膜潛力高于豬源;23株產(chǎn)膜菌株中,強產(chǎn)膜菌株2株(占比2/23=8.70%),中產(chǎn)膜菌株9株(占比9/23=39.13%),弱產(chǎn)膜菌株12株(占比12/23=52.17%)。
2.2 最小抑菌濃度測定
23株產(chǎn)生物被膜的菌株中,來源不同的金黃色葡萄球菌對22種抗菌藥物的敏感程度出現(xiàn)明顯差異,豬源較牛乳源耐藥情況極嚴(yán)重。牛乳源中分離得到的金黃色葡萄球菌對所有藥物均表現(xiàn)敏感,耐藥值較低;但值得注意的是,1株豬源金黃色葡萄球菌對青霉素、阿莫西林、頭孢噻呋、頭孢西丁、恩諾沙星、環(huán)丙沙星、克林霉素、多西環(huán)素、紅霉素、氟苯尼考、復(fù)方新諾明、泰妙菌素、替米考星均表現(xiàn)出耐藥(表4),耐藥值遠(yuǎn)高于其折點,為耐藥嚴(yán)重的多重耐藥菌株。
2.3 產(chǎn)膜菌株的分子特征
分型結(jié)果顯示,所有菌株共分為8種型,其中產(chǎn)膜的23株金黃色葡萄球菌共3種型(表5):1株t2922型來自廣東豬源,14株t2119來自浙江牛乳源,8株t189來自福建牛乳源。
MLST分型結(jié)果顯示(圖1),98株菌共分為9種ST型,其中6種ST型的菌株不具有產(chǎn)生生物被膜的能力,分別為:ST398、ST522、ST705、ST1651、ST479和ST151,而3種ST型的菌株具有生物被膜產(chǎn)生能力,分別為ST9、ST7和ST188;通過PCA進(jìn)行聚類分析:以PCA1和PCA2分別為橫縱坐標(biāo)展示散點圖的分類狀況。如圖2所示,主成分PCA1 和PCA2 分別占據(jù)所有成分中63.62%和17.74%,表明在這兩個主要成分可以很好的對菌株進(jìn)行組間聚類,弱產(chǎn)膜菌株ST7型能聚在一起,且與中強產(chǎn)膜菌株很好的區(qū)分開來,推測只有特定ST型的金黃色葡萄球菌才具有產(chǎn)生生物被膜的能力。

圖1 BF產(chǎn)生能力與ST型之間的關(guān)系

表4 產(chǎn)膜菌株分子分型和耐藥表型
PFGE分型結(jié)果顯示(圖3),產(chǎn)膜菌株共出現(xiàn)3種PFGE型,其中,廣東、福建、浙江各分為1種,顯示PFGE型可能存在地域分布特征。
綜上,分子分型結(jié)果顯示,廣東產(chǎn)膜菌株分子型為ST9-t2922-PFGEII(n=1),浙江產(chǎn)膜菌株分子型為ST7-t2119-PFGEI(n=14),福建產(chǎn)膜菌株分子型為ST188-t189-PFGEIII(n=8)。
2.4 全基因組測序
對23株生物被膜產(chǎn)生菌株進(jìn)行WGS測序,將數(shù)據(jù)提交到網(wǎng)絡(luò)數(shù)據(jù)庫進(jìn)行耐藥基因和毒力基因的查找,如圖4顯示,8株福建牛乳源的ST188-t189均攜帶包括、、、、、、及殺白細(xì)胞介素相關(guān)毒力因子、在內(nèi)的9種不同的毒力基因,14株浙江牛乳源的ST7-t2119還攜帶金黃色葡萄球菌激酶毒素因子、補體抑制劑毒素因子和腸毒素在內(nèi)的12種不同類型的毒力基因;值得注意的是,1株廣東豬源ST9-t2922主要攜帶氨基糖苷類、-內(nèi)酰胺類、大環(huán)類脂類、酰胺醇類、四環(huán)素等耐藥基因和包括、、、及6種腸毒素因子在內(nèi)的共10種毒力基因,該菌還同時攜帶甲氧西林類耐藥基因,分型顯示其攜帶XII型基因盒,即該菌株為能產(chǎn)生物被膜的耐甲氧西林金黃色葡萄球菌(MRSA)。所有菌株均攜帶調(diào)控多糖黏附素(PIA)合成的功能基因和調(diào)控表面蛋白合成的黏附基因、、等。

圖2 生物被膜產(chǎn)生菌株中ST型的分布
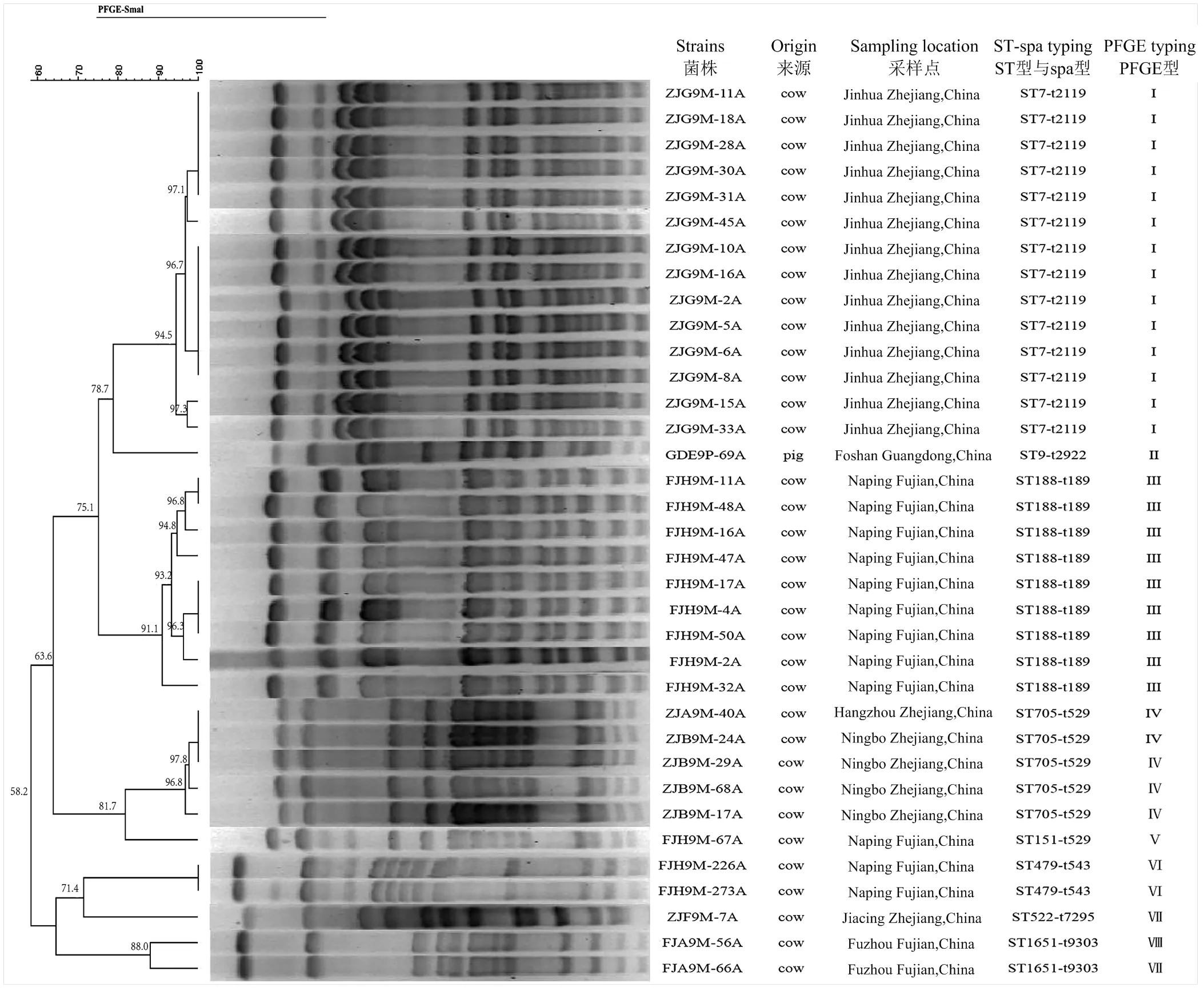
cow:奶牛;pig:豬。Jinhua Zhejiang,China:中國浙江省金華市;Hangzhou Zhejiang,China:中國浙江省杭州市;Ningbo Zhejiang,China:中國浙江省寧波市;Jiaxing Zhejiang, China:中國浙江省嘉興市;Nanping Fujian,China:中國福建省南平市;Fuzhou Fujian,China:中國福建省福州市;Foshan Guangzhou,China:中國廣東省佛山市

表5 金黃色葡萄球菌spa分型特征
3 討論
3.1 牛乳源金黃色葡萄球菌耐藥特點和分子特點
金黃色葡萄球菌不僅能引起多種動物疾病,且具有產(chǎn)生物被膜的生物學(xué)特性。生物被膜的存在更容易造成患病動物持續(xù)感染和治療失敗,其復(fù)雜的動態(tài)調(diào)控機(jī)制使得生物被膜的消除成為臨床重大難題[18-20],為解決這一問題而盲目更換抗菌藥種類和加大用藥劑量只會導(dǎo)致耐藥現(xiàn)狀加劇[21]。本研究發(fā)現(xiàn),98株金黃色葡萄球菌中牛乳源的產(chǎn)膜率明顯高于豬源,而生物被膜的存在更有利于細(xì)菌不斷繁殖釋放從而造成持續(xù)感染,這可能是牛更容易患乳房炎的原因;對產(chǎn)膜菌株的藥物敏感性檢測結(jié)果顯示,14株浙江牛乳源和8株福建牛乳源金黃色葡萄球菌對22種抗菌藥物敏感,與吳帆等[22]2017年從寧夏牛乳源中分離得到的金黃色葡萄球菌耐藥率相似,較郭慧琴等[23]2019年在杭州奶牛養(yǎng)殖廠分離得到的金黃色葡萄球菌耐藥率低,而1株廣東豬源MRSA則表現(xiàn)為多重耐藥。推測造成耐藥表型差異的主要原因是地域差異和各廠區(qū)養(yǎng)殖管理模式不同,一般規(guī)模化養(yǎng)殖場中消洗嚴(yán)格并存在相對成熟的飼養(yǎng)管理模式,能大大降低疾病發(fā)生幾率,且奶牛養(yǎng)殖場中使用抗菌藥頻率較低,用藥種類單一,治療多針對患病奶牛[24],這可能是牛乳源金黃色葡萄球菌耐藥普遍較低的原因。
3.2 生物被膜產(chǎn)生與分子型之間的相關(guān)性
分子分型結(jié)果表明,產(chǎn)膜優(yōu)勢ST型為ST9、ST7和ST188,推測只有特定ST型的金黃色葡萄球菌才具有產(chǎn)生生物被膜的能力。PCA相關(guān)性分析證明菌株生物被膜產(chǎn)生強弱很可能與ST型有關(guān),與CHEN[25]等已報道寵物源金黃色葡萄球菌產(chǎn)膜強弱可能與ST型有關(guān)相似。但值得注意的是,來源相同的同種ST型的菌株中,生物被膜產(chǎn)生強弱卻是各不相同的,其中原因值得深入探究。金黃色葡萄球菌是造成奶牛感染臨床型乳房炎的最主要病原菌,而特定ST型的菌株,因生物被膜的產(chǎn)生更容易聚集粘附于宿主,逐漸發(fā)展成為優(yōu)勢菌株并長期定植,造成無臨床癥狀的隱形乳房炎,其病理特征往往無法通過肉眼觀察,進(jìn)而發(fā)展成為慢性乳房炎導(dǎo)致乳房萎縮和硬結(jié)[26-27],極大降低產(chǎn)奶效率[28-29];全基因組測序結(jié)果表明,8株福建牛乳源的ST188-t189-PFGEIII均攜帶9種不同的毒力基因,14株浙江牛乳源的ST7-t2119-PFGEI分別同時攜帶12種不同類型的毒力基因,值得注意的是,1株廣東豬源ST9-t2922-PFGEII的中產(chǎn)膜菌株,攜帶10種耐藥基因,其中包括介導(dǎo)苯唑西林耐藥的甲氧西林類耐藥基因,但前人研究表明,我國動物源ST9-MRSA主要型為t899[29-31],而本試驗中分離得到的ST9-MRSA-SCCXII-t2922型在我國報道極少,同時該菌株還攜帶有10種不同的毒力因子,為攜帶多重耐藥基因和毒力基因且能產(chǎn)生生物被膜的MRSA。多種耐藥基因和毒力基因共存于MRSA菌株使得臨床治療極為棘手,且其產(chǎn)膜能力更是為該菌株發(fā)展為優(yōu)勢菌株并造成克隆傳播提供了條件,該類菌株的存在和傳播將給公共衛(wèi)生安全帶來極大威脅。
本研究以獸醫(yī)臨床分離得到的金黃色葡萄球菌為研究對象,對菌株生物被膜產(chǎn)生能力進(jìn)行檢測,并分析產(chǎn)膜陽性菌株的耐藥特點和分子特點,探究了生物被膜產(chǎn)生與分子型之間的相關(guān)性,為研究生物被膜的消除和治療動物金黃色葡萄球菌感染提供參考。
4 結(jié)論
對生物被膜產(chǎn)生能力進(jìn)行探究發(fā)現(xiàn),牛乳源和豬源金黃色葡萄球菌都有產(chǎn)生生物被膜的潛力,且牛乳源分離株較豬源分離株生物被膜產(chǎn)生能力更強,這將導(dǎo)致患病動物的隱性感染和持續(xù)感染;產(chǎn)膜菌株的藥物敏感性結(jié)果顯示,牛乳源分離株對受試22種常見抗菌藥物敏感,而豬源產(chǎn)膜株表現(xiàn)為多重耐藥;WGS分析耐藥基因與毒力基因發(fā)現(xiàn),所有菌株均攜帶產(chǎn)膜相關(guān)的功能基因和黏附類毒力基因,此外,廣東豬源MRSA產(chǎn)膜株還攜帶10種耐藥基因和介導(dǎo)苯唑西林耐藥的甲氧西林類耐藥基因,該類具有產(chǎn)膜能力的多重耐藥菌株給臨床治療帶來更大困難;菌株能否產(chǎn)膜與ST型存在強相關(guān)性,產(chǎn)膜優(yōu)勢ST型僅為ST9、ST7和ST188型;金黃色葡萄球菌生物被膜的產(chǎn)生存在著復(fù)雜的調(diào)控機(jī)制,找尋其關(guān)鍵原因和解決辦法仍值得我們不斷思考和探究。
[1] NEAMAH A J, AYYEZ H N, KLAIF S F, KHUDHAIR Y I, HUSSAIN M H. Molecular and phylogenetic study ofisolated from human and cattle of Al-Qadisiyah Governorate, Iraq. Veterinary World, 2019, 12(9): 1378-1382. doi:10.14202/ vetworld.2019.1378-1382
[2] 蘇洋, 蒲萬霞, 陳智華, 鄧海平. 牛源金黃色葡萄球菌的耐藥性及耐甲氧西林金黃色葡萄球菌的檢測. 中國農(nóng)業(yè)科學(xué), 2012, 45(17): 3602-3607.
SU Y, PU W X, CHEN Z H, DENG H P. Antimicrobial resistance analysis and detection of methicillin-resistant(MRSA) amongstrains isolated from bovine mastitis. Scientia Agricultura Sinica, 2012, 45(17): 3602-3607. (in Chinese)
[3] LI H, ANDERSEN P S, STEGGER M, SIEBER R N, INGMER H, STAUBRAND N, DALSGAARD A, LEISNER J J. Antimicrobial resistance and virulence gene profiles of methicillin-resistant and- susceptiblefrom food products in Denmark. Frontiers in Microbiology, 2019, 10: 2681. doi:10.3389/fmicb.2019. 02681.
[4] 王璇, 王娉, 葛毅強, 陳穎. 食品中金黃色葡萄球菌致病性研究進(jìn)展. 中國人獸共患病學(xué)報, 2017, 33(6): 553-558. doi:10.3969/j.issn. 1002-2694. 2017.06.016.
WANG X, WANG P, GE Y Q, CHEN Y. Review on pathogenicity ofin food. Chinese Journal of Zoonoses, 2017, 33(6): 553-558. doi:10.3969/j.issn.1002-2694.2017.06.016. (in Chinese)
[5] FANG R D, CUI J C, CUI T T, GUO H Y, ONO H K, PARK C H, OKAMURA M, NAKANE A, HU D L. Staphylococcal enterotoxin C is an important virulence factor for mastitis. Toxins, 2019, 11(3): 141. doi:10.3390/toxins11030141.
[6] JAVDAN S, NARIMANI T, ABADI M S S, GHOLIPOUR A. Agr typing ofspecies isolated from clinical samples in training hospitals of Isfahan and Shahrekord. BMC Research Notes, 2019, 12(1): 363. doi:10.1186/s13104-019-4396-8.
[7] SONESSON A, PRZYBYSZEWSKA K, ERIKSSON S, M?RGELIN M, KJELLSTR?M S, DAVIES J, POTEMPA J, SCHMIDTCHEN A. Identification of bacterial biofilm and thederived protease, staphopain, on the skin surface of patients with atopic dermatitis. Scientific Reports, 2017, 7: 8689. doi:10.1038/ s41598-017-08046-2.
[8] OTTO M.biofilms. Microbiology Spectrum, 2018, 6(4). doi:10.1128/microbiolspec. GPP3-0023-2018.
[9] BASANISI M G, BELLA G L, NOBILI G, FRANCONIERI I, SALANDRA G L. Genotyping of methicillin-resistant(MRSA) isolated from milk and dairy products in South Italy. Food Microbiology, 2017, 62: 141-146. doi:10.1016/j.fm.2016.10.020.
[10] LI X L, ZHOU Y, ZHAN X L, HUANG W C, WANG X. Breast milk is a potential reservoir for livestock-associatedand community-associatedin Shanghai, China. Frontiers in Microbiology, 2018, 8: 2639. doi:10.3389/fmicb.2017. 02639.
[11] MA D Z, MANDELL J B, DONEGAN N P, CHEUNG A L, MA W Y, ROTHENBERGER S, SHANKS R M Q, RICHARDSON A R, URISH K L. The toxin-antitoxin MazEF drivesbiofilm formation, antibiotic tolerance, and chronic infection. mBio, 2019, 10(6): e01658-19. doi:10.1128/mBio.01658-19.
[12] OTTO M. Staphylococcal biofilms. Microbiology Spectrum, 2018, 6(4): 207-228. doi:10.1128/microbiolspec.gpp3-0023-2018.
[13] KIM M K, ZHAO A S, WANG A, BROWN Z Z, MUIR T W, STONE H A, BASSLER B L. Surface-attached molecules controlquorum sensing and biofilm development. Nature Microbiology, 2017, 2: 17080. doi:10.1038/nmicrobiol.2017. 80.
[14] MOORMEIER D E, BAYLES K W.biofilm: a complex developmental organism. Molecular Microbiology, 2017, 104(3): 365-376. doi:10.1111/mmi.13634.
[15] 房詩薇, 黃玲利, 謝書宇, 王玉蓮, 瞿瑋, 程古月, 劉振利, 袁宗輝, 郝海紅. 獸用抗菌藥耐藥判定標(biāo)準(zhǔn)的研究進(jìn)展. 中國抗生素雜志, 2019, 44(6): 667-673. doi:10.13461/j.cnki.cja.006669.
FANG S W, HUANG L L, XIE S Y, WANG Y L, QU W,CHENG G Y, LIU Z L, YUAN Z H, HAO H H. Study progress on breakpoints of veterinary antibiotics. Chinese Journal of Antibiotics, 2019, 44(6): 667-673. doi:10.13461/j.cnki.cja.006669. (in Chinese)
[16] 張林吉, 張小榮, 曹永忠, 巢國祥, 任士飛, 吳艷濤. 金黃色葡萄球菌腸毒素基因與MLST及spa分子克隆相關(guān)性研究. 中國病原生物學(xué)雜志, 2018, 13(5): 449-456. doi:10.13350/j.cjpb.180501.
ZHANG L J, ZHANG X R, CAO Y Z, CHAO G X, REN S F, WU Y T. Diversity of enterotoxin genes ofand characterization ofclones. Journal of Pathogen Biology, 2018, 13(5): 449-456. doi:10.13350/j.cjpb.180501. (in Chinese)
[17] ENRIGHT M C, DAY N P, DAVIES C E, PEACOCK S J, SPRATT B G. Multilocus sequence typing for characterization of methicillin- resistant and methicillin-susceptible clones of. Journal of Clinical Microbiology, 2000, 38(3): 1008-1015. doi:10. 1128/JCM.38.3.1008-1015.2000.
[18] 劉保光, 蔡田, 李小申, 劉營營, 賀丹丹, 匡秀華, 高延玲, 胡功政. 牛奶源金黃色葡萄球菌血清型、毒力基因及PFGE分型. 食品科學(xué), 2019, 40(2): 281-286. doi:10.7506/spkx1002-6630-20180110-124.
LIU B G, CAI T, LI X S, LIU Y Y, HE D D, KUANG X H, GAO Y L, HU G Z. Serotyping, virulence genes and pulsed field gel electrophoresis (PFGE) typing ofisolated from fresh milk. Food Science, 2019, 40(2): 281-286. doi:10.7506/spkx1002-6630- 20180110-124. (in Chinese)
[19] LUTHER M K, PARENTE D M, CAFFREY A R, DAFFINEE K E, LOPES V V, MARTIN E T, LAPLANTE K L. Clinical and genetic risk factors for biofilm-forming. Antimicrobial Agents and Chemotherapy, 2018, 62(5): e02252-17. doi:10.1128/ AAC.02252-17.
[20] PAHARIK A E, HORSWILL A R. The staphylococcal biofilm: adhesins, regulation, and host response. Microbiology Spectrum, 2016, 4(2): 10.1128/microbiolspec.VMBF-22. doi:10.1128/microbiolspec. VMBF-0022-2015.
[21] LAKSHMI S A, BHASKAR J P, KRISHNAN V, SETHUPATHY S, PANDIPRIYA S, ARUNI W, PANDIAN S K. Inhibition of biofilm and biofilm-associated virulence factor production in methicillin- resistantby docosanol. Journal of Biotechnology, 2020, 317: 59-69. doi:10.1016/j.jbiotec.2020.04.014.
[22] 吳帆. 寧夏地區(qū)牛源金黃色葡萄球菌生物被膜相關(guān)基因檢測及耐藥性研究[D]. 銀川: 寧夏大學(xué), 2017.
WU F. Study on detection of bofilm genes and antimicrobial resistance ofisolated from cow in ningxia[D]. Yinchuan: Ningxia University, 2017. (in Chinese)
[23] 郭慧琴, 李田, 肖鵬, 余茂林, 姜中其. 牛源金黃色葡萄球菌的分離鑒定、耐藥性分析及木糖醇對生物被膜形成的干預(yù). 中國獸醫(yī)學(xué)報, 2019, 39(5): 889-893. doi:10.16303/j.cnki.1005-4545.2019.05.14.
GUO H Q, LI T, XIAO P, YU M L, JIANG Z Q. Isolation, identification and bacterial resistance of dairy cows'and intervention of biofilm formation by xylitol. Chinese Journal of Veterinary Science, 2019, 39(5): 889-893. doi:10.16303/ j.cnki.1005-4545.2019.05.14. (in Chinese)
[24] 李寶明, 戴命子, 李學(xué)釗. 奶牛乳房炎的診治. 中國牛業(yè)科學(xué), 2019, 45(6): 92-93. doi:10.3969/j.issn.1001-9111.2019.06.030.
LI B M, DAI M Z, LI X Z. Diagnosis and treatment of mastitis in dairy cows. China Cattle Science, 2019, 45(6): 92-93. doi:10.3969/ j.issn.1001-9111.2019.06.030. (in Chinese)
[25] CHEN L, TANG Z Y, CUI S Y, MA Z B, DENG H, KONG W L, YANG L W, LIN C, XIONG W G, ZENG Z L. Biofilm production ability, virulence and antimicrobial resistance genes infrom various veterinary hospitals. Pathogens (Basel, Switzerland), 2020, 9(4): 264. doi:10.3390/pathogens9040264.
[26] ANTóK F I, MAYRHOFER R, MARBACH H, MASENGESHO J C, KEINPRECHT H, NYIRIMBUGA V, FISCHER O, LEPUSCHITZ S, RUPPITSCH W, EHLING-SCHULZ M, FE?LER A T, SCHWARZ S, MONECKE S, EHRICHT R, GRUNERT T, SPERGSER J, LONCARIC I. Characterization of antibiotic and biocide resistance genes and virulence factors ofspecies associated with bovine mastitis in Rwanda. Antibiotics (Basel, Switzerland), 2019, 9(1): 1. doi:10.3390/antibiotics9010001.
[27] 吳文學(xué), 李秀波, 王瑞, 王朋朋, 田欣睿, 張海燕, 焦曉宇, 李金祥. 奶牛乳房炎的發(fā)病機(jī)制. 中國獸醫(yī)雜志, 2019, 55(10): 113-116.
WU W X, LI X B, WANG R, WANG P P, TIAN X R, ZHANG H Y, JIAO X Y, LI J X. Pathogenesis of mastitis in dairy cows. Chinese Journal of Veterinary Medicine, 2019, 55(10): 113-116. (in Chinese)
[28] LIMA M G B, BLAGITZ M G, SOUZA F N, SANCHEZ E M R, BATISTA C F, BERTAGNON H G, DINIZ S A, SILVA M X, LIBERA A M, DELLA M P. Profile of immunoglobulins, clinical and bacteriological cure after different treatment routes of clinical bovine mastitis.Arquivo Brasileiro de Medicina Veterinaria e Zootecnia2018,70(4):1141-1149.doi: 10.1590/1678-4162-9695.
[29] Prevalence of bovine mastitis in the Anaime Canyon, a Colombian dairy region, including etiology and antimicrobial resistance. Revista de Investigaciones Veterinarias del Perú, 2018, 29(1):226-293.doi.org/10.15381/rivep.v29i1.14084.
[30] CUI S H, LI J Y, HU C Q, JIN S H, LI F Q, GUO Y C, RAN L, MA Y. Isolation and characterization of methicillin-resistantfrom swine and workers in China. Journal of Antimicrobial Chemotherapy, 2009, 64(4): 680-683. doi:10.1093/jac/dkp275.
[31] 李德喜. 惡唑烷酮類耐藥基因cfr和optrA在豬源MRSA和CoNS中流行及傳播機(jī)制的研究[D]. 北京: 中國農(nóng)業(yè)大學(xué), 2016.
LI D X. The epidemiological study on the oxazolidinones resistance genescfr and OptrA and theirs transmission mechanism among MRSA and CoNS isolates from swine[D]. Beijing: China Agricultural University, 2016. (in Chinese)
Relationship Between Biofilm Formation and Molecular Typing offrom Animal Origin
TANG ZiYun, HU JianXin, CHEN Jin, LU YiXing, KONG LingLi, DIAO Lu, ZHANG FaFu, XIONG WenGuang, ZENG ZhenLing*
College of Veterinary Medicine, South China Agricultural University/Guangdong Provincial Key Laboratory of Veterinary Pharmaceutics Development and Safety Evaluation/Guangdong Laboratory for Lingnan Modern Agriculture/National Risk Assessment Laboratory for Antimicrobial Resistance of Animal Original Bacteria, Guangzhou 510642
【Objective】The aim of this study was to investigate the epidemiological characteristics of()in the biofilm producing strains and to explore the correlation between biofilm forming ability and molecular typing, so as to provide the theoretical basis for the treatment ofinfection.【Method】The biofilm producing ability of all strains ofwas determined by crystal violet semi-quantitative method. The minimum inhibitory concentrations of 22 common antibiotics were determined by the membrane producing strains. Molecular typing was conducted by three common typing methods of, includingtyping, MLST typing and PFGE typing, and the correlation between membrane production capacity and molecular typing was analyzed. Finally, whole genome sequencing technology was used to analyze the antibiotics resistance gene and virulence genes in the biofilm producing strains. 【Result】The semi-quantitative results of crystalline violet showed that a total of 23 strains (23.47%) of 98strains were able to produce biofilm, including 22 strains (25.29%, 22/87) from cow milk source, 14 strains (60.87%, 14/22) from Zhejiang dairy farms, 8 strains (39.13%, 8/22) from Fujian dairy farms, and 1 strain from pig source (9.10%, 1/11) from Guangdong slaughterhouse, indicating that the film-producing potential offrom cow's milk source was higher than that of pig source, 22 strains (95.65%) of which were from cow's milk source and 1 strain (4.35%) was from pig source. The film-producing ability was classified into strong, medium and weak, and among the 23 film-producing strains, 2 strains (8.70%, 2/23) were strong film-producing strains, 9 strains (39.13%, 9/23) were medium, and 12 strains (52.17%, 12/23) were weak. The results of the drug sensitivity test showed that the bovine milk-derived membrane- producing strains were sensitive to all the tested antibacterial drugs, while the pig-derived membrane-producing strains showed resistance to 13 antibacterial drugs, including penicillin, amoxicillin, ceftiofur, cefoxitin, enrofloxacin, ciprofloxacin, clindamycin, doxycycline, erythromycin, flupenthixol, cotrimoxazole, tiamulin, and tilmicosin. Thetyping results showed that 98 strains ofobtained 8types, and 23 strains of film-producingaccounted for 3 of them: 1 strain t2922 from porcine origin in Guangdong, 14 strains t2119 from Zhejiang cow milk source, and 8 strains t189 from Fujian cow milk source. MLST typing results showed that 98 strains were classified into 9 ST types, of which 6 ST types did not have the ability to produce biofilm, namely ST398, ST522, ST705, ST1651, ST479 and ST151, and only 3 strains of ST type had biofilm production ability, namely ST9, ST7 and ST188. It was found that the molecular types of strong film-producing strains were mainly ST7-t2119, the medium film-producing strains were mainly ST7-t2119 and ST188-t189, and the weak film-producing strains were ST9-t2922, ST7-t2119 and ST188-t189. The ST type of weak film-producing strains could be well distinguished from the medium and strong film-producing strains, and only the specific ST type ofhad the ability to produce biofilm. 23 film-producing strains PFGE typing all successful PFGE typing results showed that the results show that each strain of film-producing bacteria in Guangdong, Fujian and Zhejiang provinces were divided into 3 PFGE types, and there were geographical distribution characteristics of PFGE types; the strains isolating from the same region had clonal transmission, and the strains in the province were clonal to each other, but there were significant differences in biofilm production ability between clones.; the whole genome sequencing results showed that the drug resistance genes and virulence genes in the film-producing strains were diverse according to the molecular type. 【Conclusion】from different sources had different potential to produce biofilm and all carried different film-producing genes. The film-producing potential offrom bovine milk source was much higher than that of porcine source, and all carrid different film-producing genes. Whether strains could produce film or not may be strongly correlated with ST type, and the specific ST types, such as ST9, ST7 and ST188, were more likely to produce biofilm; however, at the same time, the strains with the same molecular type had different abilities to produce biofilm.
; biofilm; antibiotics resistance gene; molecular typing; virulence genes
2020-12-10;
2021-07-08
“十三五”國家重點研發(fā)計劃(2018YFD0500300)
唐子云,Tel:15827491246;E-mail:1598387644@qq.com。通信作者曾振靈,Tel:13609768425;E-mail:zlzeng@scau.edu.cn
(責(zé)任編輯 林鑒非)
猜你喜歡
天天愛科學(xué)(2022年9期)2022-09-15 01:12:54
保健醫(yī)苑(2022年5期)2022-06-10 07:46:38
現(xiàn)代臨床醫(yī)學(xué)(2022年3期)2022-06-06 07:59:40
天天愛科學(xué)(2022年4期)2022-05-23 12:41:48
當(dāng)代水產(chǎn)(2022年3期)2022-04-26 14:26:56
昆明醫(yī)科大學(xué)學(xué)報(2022年1期)2022-02-28 07:43:40
科學(xué)大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
科學(xué)大眾(2020年12期)2020-08-13 03:22:22
航空世界(2020年10期)2020-01-19 14:36:20
