邊緣革蜱Dm 86基因表達及免疫原性分析
2022-02-21 07:18:10李敏馬英呼爾查何文文史倩云阿力木江加帕爾蔣倩巴音查汗
新疆農業科學 2022年9期
李敏,馬英,呼爾查,何文文,史倩云,阿力木江·加帕爾,蔣倩,巴音查汗
(新疆農業大學動物醫學學院,烏魯木齊 830052)
0 引言
【研究意義】邊緣革蜱(Dermacenter marginatus,D.marginatus)為三宿主蜱[1]。我國主要分布于新疆,是攜帶多種病原的地方優勢種媒介蜱,可攜帶并傳播巴貝斯蟲、泰勒蟲、立克次體、布魯氏菌、和萊姆等多種蜱傳病原體[2]。近年來,我國新疆昭蘇、和碩、博樂、哈巴河等邊緣革蜱孳生數量逐漸增加[3]。降低蜱及蜱傳疾病風險的主要方法有化學殺螨劑的應用[4]。疫苗接種可使宿主增強免疫力,針對性消滅不同蜱種,污染率低。挖掘抗蜱疫苗候選基因及其保護性抗原分子的研究極其重要,是一個有效防蜱控蜱的研究方向,具有良好的應用前景。【前人研究進展】Boophilus microplus86(Bm86)抗原是被證明免疫后有效的商品化疫苗。Willadsen等[5-6]從半飽血微小牛蜱雌蜱腸道中分離出一種膜結合糖蛋白,用微克級的抗原免疫牛所產生的抗體即對牛有抗蜱保護作用,當微小牛蜱吸食了用Bm86抗原免疫的牛的血液時,血液中的抗體即與蜱腸道表面的相應糖蛋白結合,使蜱腸道受到嚴重損傷,當牛血液滲入蜱的血淋巴后,能更進一步引起蜱的大規模死亡及繁殖和生存能力的下降等,并已證明Bm86為一種“隱藏抗原”。國外針對其優勢蜱微小牛蜱的Bm86的基因工程疫苗已在多國注冊并進行田間試驗中,控蜱效果在50%~100%[7],降低了蜱傳疾病的發病率。【本研究切入點】在Bm86其它同系物研究中,璃眼蜱的Hd86抗原可能在若蜱蛻皮為成蜱階段起作用,長角血蜱HI86蛋白的宿主動物能可有效降低飽血雌蜱的體重,在篦子硬蜱Ir86抗原的免疫實驗中,并未觀察到顯著吸血和產卵階段變化[8-10]。關于新疆優勢硬蜱的保護性抗原的研究未見報道。邊緣革蜱Dm86能否作為保護性抗原發揮作用還需要相關的研究。【擬解決的關鍵問題】擬克隆Dm86基因CDs區核酸片段,經采用生物學分析,篩選親水性較高、抗原性強和抗原表位連續的核酸片段,構建重組質粒,表達蛋白后制備其多克隆抗體驗證其免疫原性,分析蜱蟲不同發育階段的Dm86基因相對表達量,為抗邊緣革蜱疫苗保護性抗原的研究奠定基礎。
1 材料與方法
1.1 材料
1.1.1 樣品及試驗動物
所用邊緣革蜱F3代由新疆農業大學寄生蟲實驗室飼養,供血動物為3月齡新西蘭大白兔1只,體重2.1 kg,購自新疆醫科大學實驗動物中心。
1.1.2 質粒、菌種及試劑
所使用的主要試劑材料中pMD19-T載體、T4連接酶、BL21感受態細胞、Blue Plus Protein Marker、DNA Marker購自北京全式金生物技術有限公司;RNAisoPlus、大腸桿菌DH5a感受態細胞、Fast-King cDNA第一鏈合成試劑盒、2×TaqPCR Mix、普通DNA產物純化回收試劑盒、質粒小提試劑盒購自新疆寶信源柏生物技術有限公司;谷胱甘肽瓊脂糖樹脂純化試劑盒購自上海翊圣生物科技有限公司;pGEX-4T-1為質粒由實驗室保存。
1.2 引物設計與合成
1.2.1 引物設計與合成
參考實驗室所有的邊緣革蜱轉錄組數據,參照GenBank中變異革蜱Dr86的序列(GB144601.1),使用Primer Premier 5.0軟件分別設計特異性引物。表1
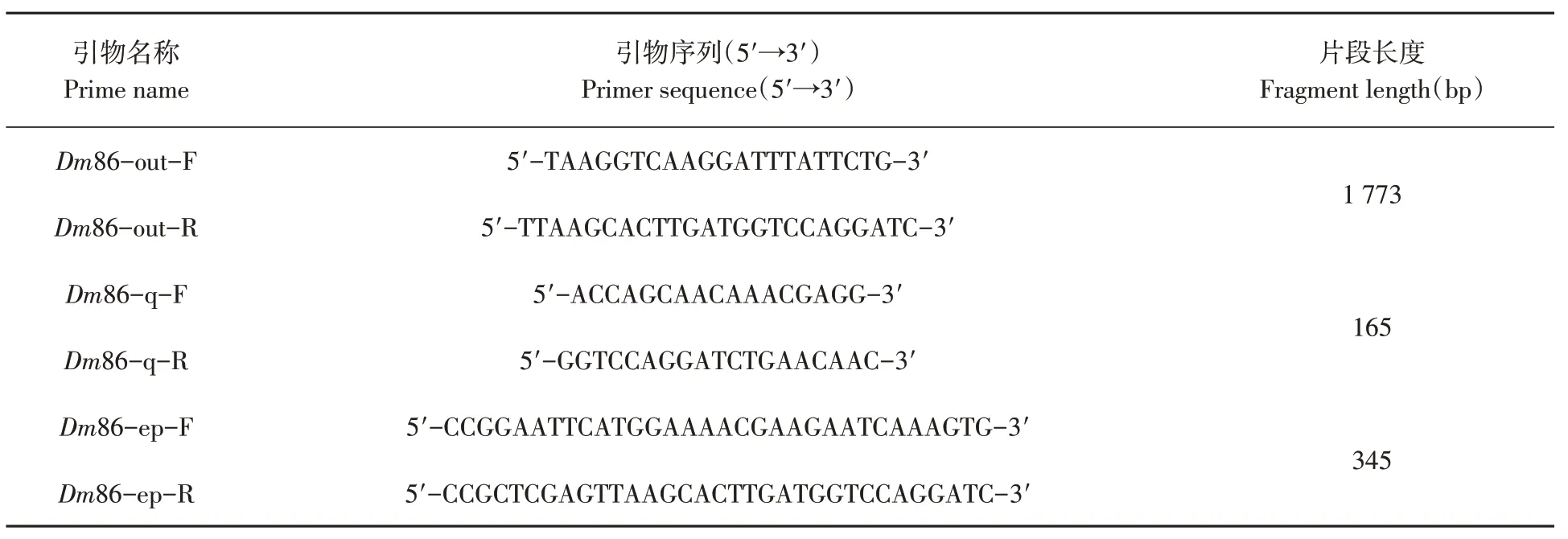
表1 Dm86基因PCR引物Table 1 Dm86 gene PCR primer
1.2.2 邊緣革蜱不同發育階段蟲體收集
以實驗室的F3代邊緣革蜱為材料,通過耳袋法將40只(雌雄各20只)接種于新西蘭大白兔的耳朵,每天打開耳套,觀察吸血情況。飽血雌蜱落體后,放置于培養皿中,溫度控制在25℃,濕度控制在85%待其產卵。卵蛻皮成為幼蜱后,接種兔耳朵繼續吸血。待幼蜱吸飽血后,蛻皮成為若蜱,接種于兔耳朵繼續吸血。待若蜱吸飽血后蛻皮后,即為成蜱。在開始吸血后第24、48、72和96 h、半飽血和飽血期和生活史的每個階段收集小部分蟲體于1.5 mL EP管。
1.2.3 邊緣革蜱總RNA的提取及第一鏈
cDNA合成以F4代邊緣革蜱各發育(卵、饑餓幼蜱、飽血幼蜱、饑餓若蜱、飽血若蜱、成蜱)及吸血(第24、48、72和96 h、半飽血、飽血)階段蟲體為組織材料,液氮研磨后Trizol法提取總RNA,按照FastKing cDNA第一鏈合成試劑盒說明書將總RNA反轉錄成cDNA。
1.2.4 Dm86基因片段的克隆
以邊緣革蜱飽血雌蜱cDNA為模板,反應體系(50μL):2×TaqPCR Mix 25μL,Dm86-out-F/Dm86-out-R各2μL,cDNA模板2μL,ddH2O補足至50μL,反應條件:95℃,預變性5 min;95℃,變性30 s;64℃,退火30 s;72℃,延伸2min,共35個循環;72℃,延伸10min。擴增產物用1%瓊脂糖凝膠電泳檢測,目的基因使用DNA產物回收試劑盒回收,純化后的目的片段與載體質粒pMD19-T連接,轉化至DH5a感受態細胞中,扎取菌落擴大培養后進行菌液PCR驗證,并將有目的條帶的菌液送往上海生工生物工程有限公司測序。
1.2.5 Dm86基因生物學
在線軟件ExPASy-ProtParam(https://web.ex-pasy.org/compute_pi/)分析目的蛋白理化性質;經ProtScale(http://web.web.expasy.org/protscale)在線分析目的蛋白的親(疏)水性;經SignalP 3.0(http://www.cbs.dtu.dk/services/SignalP-3.0/)在線分析目的蛋白信號肽;經TMHMM(http://www.cbs.dtu.dk/services/TMHMM/)在線分析目的蛋白有無跨膜結構;NetNGlyc 1.0 Server(http://www.cbs.dtu.dk/cgi-bin/webface2.)N端糖基化位點分析。經IEDB(http://tools.iedb.org/bcell/)在線預測B細胞抗原表位。NPS@(https://npsa-prabi.ibcp.fr/cgi-bin/npsa)在線蛋白分析系統對目的蛋白預測二級結構,經Phyer2和EzMol2.1在線預測目的蛋白的三級結構。
1.2.6 實時熒光定量
PCR反應以不同發育、吸血階段的邊緣革蜱cDNA為模板,延伸因子1α基因(Elongation factor 1-alpha)為內參[11],Dm86-q-F/Dm86-q-R為引物,根據QuantiNova SYBR?Green PCR試劑盒說明書,應用生物系統7 500快速實時?系統(美國應用生物系統),反應條件為95℃,2 min;95℃,15s、60℃,35 s,共40個循環。檢測Dm86在邊緣革蜱的相對表達量。
1.2.7 截短目的基因克隆
以邊緣革蜱飽血雌蜱cDNA為模板,反應體系(50μL):2×TaqPCR Mix 25μL,Dm86-ep-F/Dm86-ep-R各2μL,cDNA模板2μL,ddH2O補足至50μL,反應條件:95℃,5 min;95℃,30 s、64℃,30 s、72℃,30 s共35個循環;72℃,8 min。擴增產物用1%瓊脂糖凝膠電泳檢測,目的基因使用DNA產物回收試劑盒進行回收。純化后的目的片段與載體質粒pMD19-T連接,轉化至大腸桿菌DH5ɑ感受態細胞中,扎取菌落擴大培養后驗證菌液PCR,并將有目的條帶菌液送往上海生工生物工程有限公司測序。
1.2.8 重組質粒的構建
將重組克隆質粒pMD19-T-Dm86和表達載體pGEX-4T-1使用XhoI和EcoRI雙酶切,回收目的片段。通過T4DNA連接酶與載體質粒pGEX-4T-1連接,構建重組質粒Dm86-pGEX-4T-1并轉化至大腸桿菌BL21感受態細胞中,扎取菌落擴大培養后,提取質粒進行PCR和雙酶切驗證,將菌液送往上海生工生物工程有限公司測序驗證。
1.2.9 重組質粒Dm86-pGEX-4T-1誘導表達和純化
將Dm86-pGEX-4T-1菌液按1:100比例加入100 mL含有Amp+的LB液體培養基中,180 r/min,37℃,培養3 h,OD600達到0.6時,加入1 mmol/L IPTG進行誘導表達后,離心取上清和沉淀進行可溶性分析。按照說明書使用樹脂純化試劑盒純化Dm86融合蛋白,并通過SDS-PAGE鑒定。
1.2.10 制備多克隆抗體及Western blot分析
以純化的融合蛋白免疫新西蘭大白兔,獲得該融合蛋白的多克隆抗體,并檢測其抗體效價。以獲得的多克隆抗體作為一抗,用5%脫脂奶粉1∶100稀釋,孵育。羊抗兔IgG為二抗,用5%脫脂奶粉1∶10000稀釋,對重組蛋白進行Western blot分析。
1.3 數據處理
使用MEGA 6.0軟件,選擇T92+G+I模型,以軟蜱(Ornithodoros savignyi)為外類群,對Dm86基因構建ML(最大似然法)系統發育樹。SignalP 3.0在線分析,經NPS@在線蛋白分析系統(MLRC方法)對目的蛋白進行二級結構預測,在線預測經IEDB在線分析,單因素方差分析吸血期,收集蛋白并使用One Drop?OD-1000紫外-可見光全光譜分光光度計檢測,經SDS-PAGE驗證。
2 結果與分析
2.1 邊緣革蜱Dm86基因擴增PCR
研究表明,長片段的Dm86序列在1 773 bp出現明顯的擴增帶,與預期大小一致。圖1
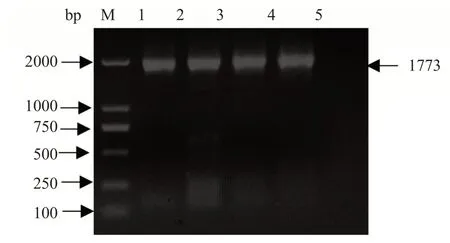
圖1 邊緣革蜱Dm86基因PCR擴增Fig.1 Result of PCR amplification of Dm86 gene of D.marginatus
2.2 邊緣革蜱Dm86基因進化
研究表明,克隆菌液測序所得序列,與NCBI所公布的網文革蜱Dermacentor reticulatus86(Dr86)的相似度為89.29%。邊緣革蜱Dm86基因序列與Dr86基因聚為一支。圖2
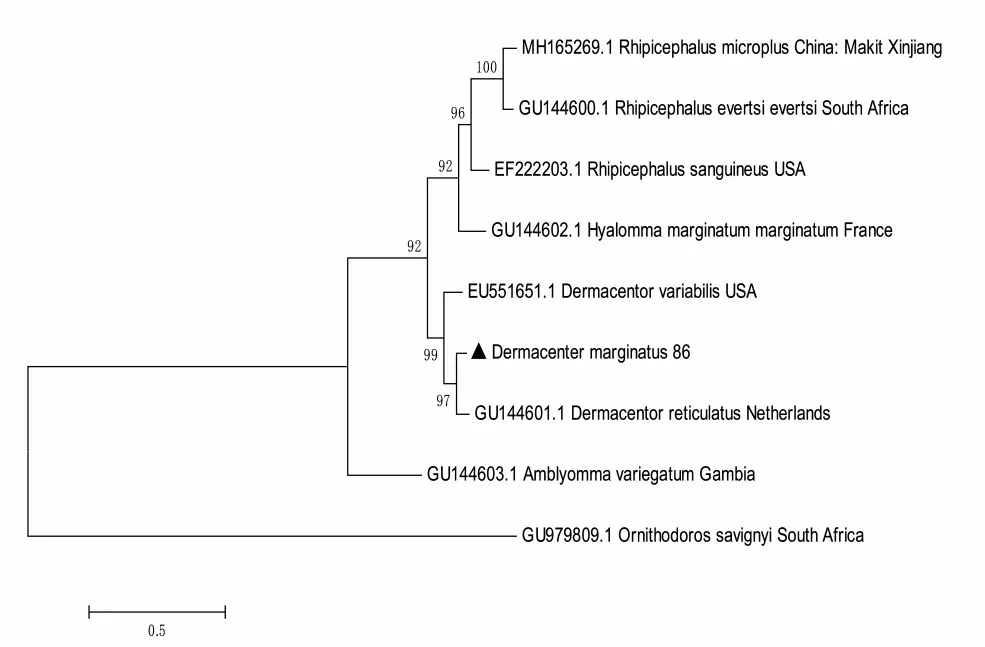
圖2 邊緣革蜱Dm86基因的系統發育Fig.2 Phylogenetic analysis of Dm86 gene of D.marginatus
2.3 生物信息學(圖3)
2.3.1 目的蛋白理化性質
研究表明,Dm86蛋白有591個氨基酸組成,分子量為64 857.81,理論等電點為6.78,酸性。在體外哺乳動物網狀細胞的半衰期為1.9 h,在酵母菌體內為20 h,在大腸桿菌體內為10 h。不穩定指數為51.77,是不穩定蛋白。脂肪系數為57.60。
2.3.2 親水性、信號肽、跨膜結構和N端糖基化位點
研究表明,目的蛋白大部分屬于親水性氨基酸,總平均親水性系數為-0.502<2,是親水性蛋白。存在信號肽。NetNGlyc 1.0 Server有5個N端糖基化位點。
2.3.3 二級結構、B細胞抗原表位和三級結構
研究表明,從高到低依次是無規則卷曲(65.48%)、延伸鏈(24.03%)和α螺旋(10.49%)。抗原性最強且抗原位點最多的區域在522~564。Phyre2 Job Status蛋白質三級結構,紅色部分抗原性最強且抗原表位連續的43個氨基酸。
2.4 邊緣革蜱生活史各階段Dm86基因熒光定量PCR檢測
研究表明,Dm86基因在邊緣革蜱不同階段均有表達。幼蜱階段的相對表達量較高,但無顯著差異;若蜱階段的相對表達量低,有顯著差異;成蜱階段的相對表達量較低,有顯著差異。以饑餓雌蜱為基準,饑餓雌蜱較緩慢吸血期和半飽血期的基因相對表達量高,但無顯著差異,較飽血階段的基因相對表達量低,存在顯著差異。圖4,圖5
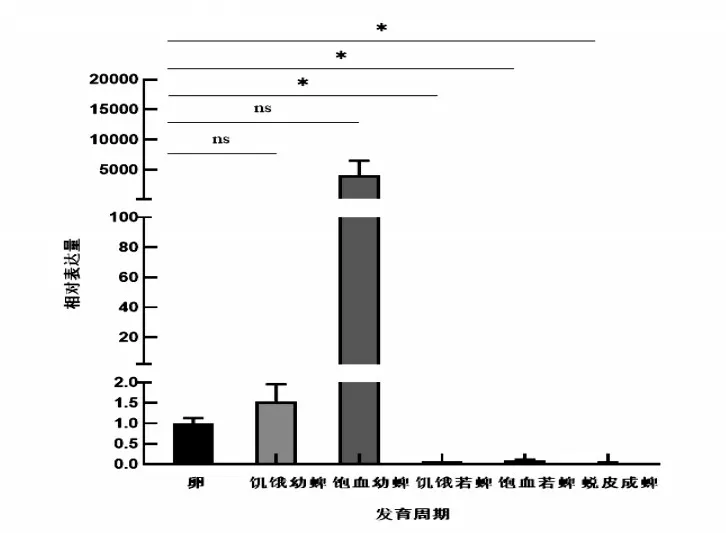
圖4 各發育周期相對表達量Fig.4 Relative expression levels of each developmental period
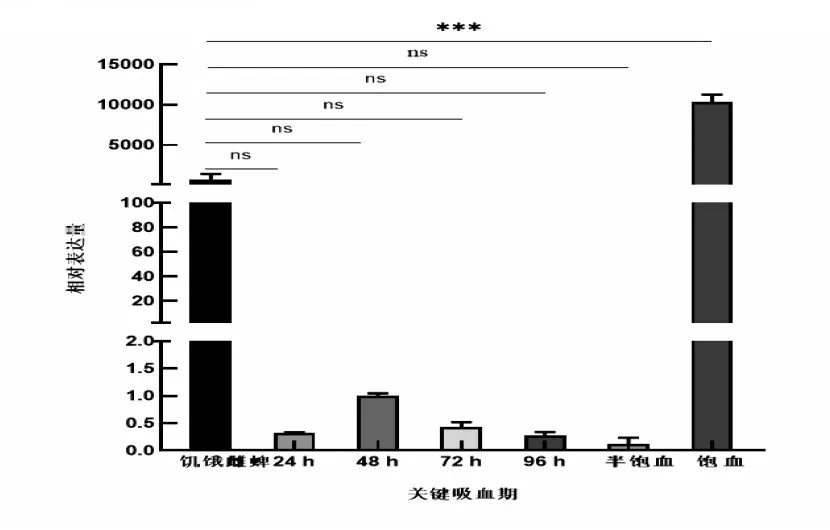
圖5 吸血期相對表達量Fig.5 Relative expression level in the bloodsucking stage
2.5 Dm86截短基因的擴增
研究表明,在345 bp出現表達性截短片段。圖6
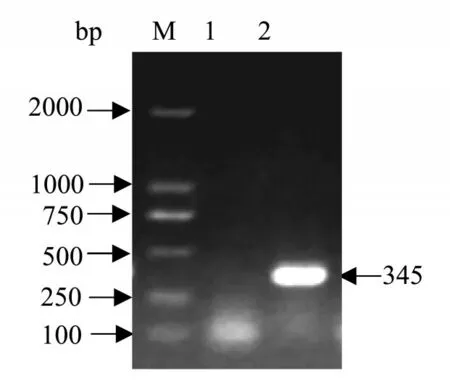
圖6 邊緣革蜱截短Dm86基因PCR擴增結果Fig.6 Result of PCR amplification of Dm86 gene of D.marginatus
2.6 重組質粒的雙酶切驗證
研究表明,重組質粒經過XhoI和EcoRI雙酶切,產物經1%瓊脂糖凝膠電泳,出現2條特異性條帶分別約為4 954和345 bp,與預期片段大小一致,送往上海生工生物工程股份有限公司進行測序。目的基因Dm86插入到表達載體pEGX-4T-1中。圖7
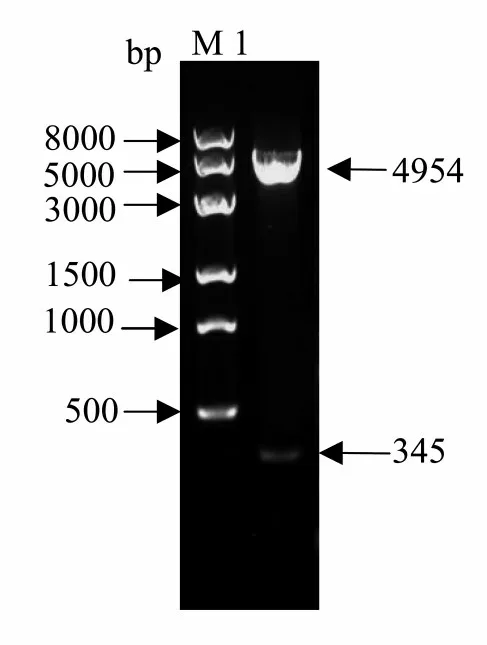
圖7 Dm86重組質粒酶切驗證Fig.7 Dm86 recombinant plasmid enzyme digestion validation
2.7 重組蛋白表達及純化
研究表明,純化濃縮蛋白,濃度為2.0 mg/mL。在39 kDa處出現條帶,與預期大小一致。圖8
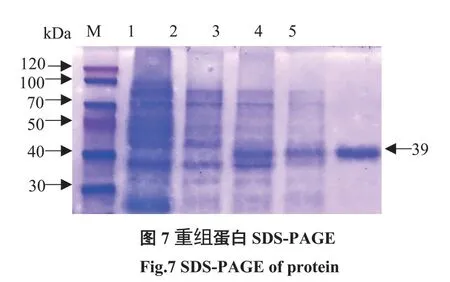
圖8 重組蛋白SDS-PAGEFig.8 SDS-PAGE of protein
2.8 多克隆抗體的制備和Western blotting分析
研究表明,多克隆抗體能夠識別重組蛋白,在39 kDa有明顯特異性條帶,陰性血清無特異性條帶。圖9
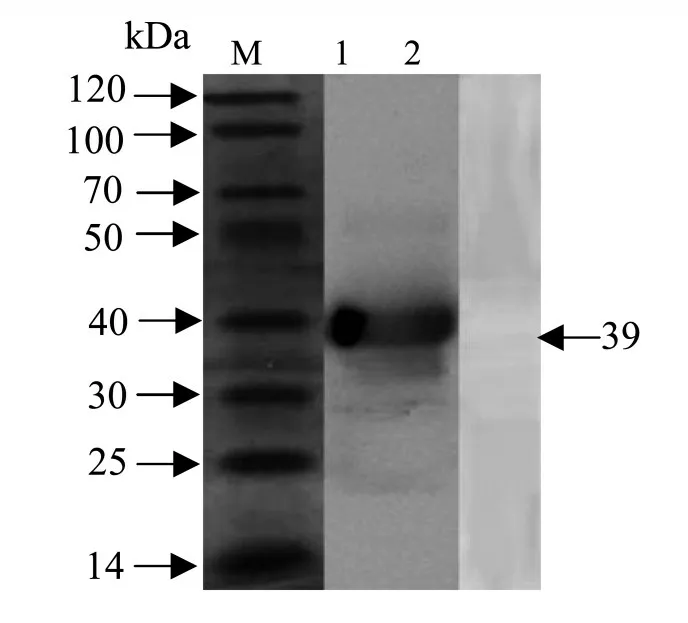
圖9 蛋白多克隆抗體的Westernblotting分析Fig.9 Western blotting analysis of polyclonal antibody against Dm86 protein
3 討論
3.1Bm86是迄今為止候選蜱抗原中最有效的基因,主要降低吸血蜱的數量、重量、產卵和蛻皮能力,減少蜱的數量和蜱媒病原體的流行[12]。通過實驗室所有的新疆優勢種邊緣革蜱轉錄組數據,參照GenBank中變異革蜱Dr86的序列(GB144601.1)克隆Dm86基因。克隆序列BLAST比對并構建系統發育樹顯示,Dm86與Dr86相似度為89.29%并聚為一支,證明邊緣革蜱體內存在Dm86基因。
3.2 對Dm86編碼的蛋白進行生物信息學預測,Dm86蛋白屬于酸性、不穩定的親水性蛋白,存在信號肽和5個N端糖基化位點,這與新疆微小牛蜱Bm86的部分理化特性相似[13]。抗原表位是抗原分子中決定抗原特異性的特殊化學集團,找到抗原表位就可能找到針對該抗原的抗體或疫苗[7]。Dm86蛋白無規則卷曲所占比例較高、親水性較強及多個抗原表位的預測結果,均提示該蛋白抗原性較強[14-16]。蛋白序列在522~564個氨基酸是無規則卷曲、抗原性最強且連續抗原表位最多的區域,為獲得免疫原性較強的蛋白,截取此區域在大腸桿菌中以重組蛋白的形式表達。
3.3 “隱蔽抗原”在自然感染條件下,不與宿主接觸引起免疫反應的一類抗原[17]。Willadsen等[5]研究表明,Bm86是位于中腸的“隱蔽抗原”,不同于其他種類的抗原,牛自然感染微小牛蜱時不會與Bm86接觸并產生抗體。試驗通過人工感染融合蛋白制備多克隆抗體,產生的抗體效價較高,并通過Western blotting驗證融合蛋白具有免疫原型,重組蛋白可以作為抗蜱疫苗的候選抗原。
3.4Dm86基因在邊緣革蜱各發育階段的相對表達量存在差異,與微小牛蜱Bm86表達情況不同[19]。微小扇頭蜱為典型的一宿主蜱,在宿主體表連續吸血,3周左右即可完成整個吸血期[20],而邊緣革蜱需要1~2年的時間完成生長,在幼蜱、若蜱和成蜱3個階段飽血后均會離開宿主,進行能量消耗和蛻皮生長,可能造成發育各周期的表達量不同。對于Bm86蛋白的功能和特定的細胞類型定位研究成果較少,邊緣革蜱的幼蜱、若蜱和成蜱階段吸血造成中腸腸腔擴大[21],飽血后的Dm86基因表達量較飽血前的高,可能與中腸細胞在吸血時期的生長和消化有關[22]。在饑餓雌蜱吸血期間,基因相對表達量下降后一直在一個相對較低的水平,可能中腸在吸收血液時消化細胞的不斷倍消耗和未分化細胞不斷取代等血液消化過程有關[21]。Dm86基因作為保護性抗原在各發育階段,均有可能發揮作用,但其真正的保護效果,還需要進一步的研究驗證熒光定量PCR的推測結果。
4 結論
PCR擴增獲得了1 773 bp目的片段,Dm86蛋白由591個氨基酸組成,分子量為64 857.81,理論等電點為6.78,酸性,具有不穩定性、親水性和多個B細胞抗原表位;Dm86基因在邊緣革蜱不同階段均有不同程度的表達,飽血蜱階段的表達量較高;重組蛋白大小為39kDa,以包涵體形式表達;抗體效價可達1:25 600,能夠識別重組蛋白。Dm86重組蛋白具有一定的反應原性和免疫原性。
