膠州灣女姑口海域大型底棲動物群落特征與生態健康評價*
2022-02-09 13:03:42劉欣禹齊衍萍沙婧婧徐子鈞菅瀟揚杜小媛溫國義周瑞佳
廣西科學 2022年6期
劉欣禹,齊衍萍,沙婧婧,徐子鈞,菅瀟揚,杜小媛,溫國義,周瑞佳**
(1.國家海洋局北海環境監測中心,山東青島 266000;2.自然資源部渤海生態預警與保護修復重點實驗室,山東青島 266000;3.山東省海洋生態環境與防災減災重點實驗室,山東青島 266000)
大型底棲動物作為海洋生態結構的重要組成部分,通過攝食、掘穴等活動參與碳、氮、磷、硫等元素的循環,有助于加快底層生態系統的物質循環和能量傳遞[1,2]。由于活動能力弱、棲息地相對穩定和對海洋環境反應敏感等特點,所以大型底棲動物群落的物種組成通常會隨著非生物條件和人類干擾的變化而變化,這使得大型底棲動物經常被用作評價底棲生境健康狀況的生物指標[2,3],對海洋生態環境研究具有積極意義。
將底棲生物作為海洋生態環境監測和生態系統健康評價的生物指標已得到國內外科學家的廣泛認可[4]。多樣性指數、AZTI海洋生物指數(AMBI)、多變量AMBI (M-AMBI) 均能在一定程度上反映底棲動物受擾動狀態,可用于評價底棲環境狀態[5]。其中,AMBI和M-AMBI是Borja等[6]在海洋生物指數的基礎上創建的。在我國及歐洲沿海,這3種指數已被用于港灣、近岸海域以及河口區的底棲生態環境質量評價[7-10]。
膠州灣位于山東半島南部,屬于相對獨立的半封閉海灣,女姑口海域位于青島市城陽區,在膠州灣的東北部,具有典型的泥質底質。20世紀80年代開始,膠州灣東部淺海海域的養殖業迅速崛起,90年代周邊城區工業和旅游業等繁榮發展,近岸海域資源被快速開發利用[11]。膠州灣環海高速公路從女姑口上通過,附近灘涂上主要開展底棲型貝類如菲律賓蛤仔(Ruditapesphilippinarum)、縊蟶(Sinonovaculaconstricta)的養殖[12,13]。麻驁等[14]研究膠州灣養殖水域和自然水域的底棲狀況發現,菲律賓蛤仔養殖活動壓力已造成養殖區域及其鄰近自然海域的大型底棲群落結構發生改變。崔正國等[15]對膠州灣底播菲律賓蛤仔的質量安全風險進行了評價。其他研究主要基于膠州灣大型底棲動物進行海洋健康狀況評估[16],針對膠州灣女姑口附近海域調查相對較少。
本研究基于2018年11月和2019年4月兩個航次膠州灣女姑口鄰近海域的大型底棲動物調查和環境要素調查數據,對膠州灣女姑口鄰近海域生物種類組成和生物群落結構進行研究,探討群落結構與環境因子的相互關系,并對該海域進行生態系統健康評價,以了解膠州灣女姑口近海底棲生物資源現狀及生態健康狀況,為膠州灣整體生態保護與管理提供數據支持。
1 材料與方法
1.1 樣品采集與處理
2018年11月、2019年4月在膠州灣女姑口近海設置12個站位進行采樣監測,站位布設見圖1。
大型底棲動物用0.1 m2抓斗式采泥器采集,每站采樣4次,樣品用網目為0.5 mm的過篩器分選,用5%福爾馬林溶液固定后,帶回實驗室進行定量定性分析,物種鑒定及名錄登記均通過WoRMS網站進行校對和復核。站位布設、生物密度及生物量分布圖由ArcGIS 10.6軟件繪制。
測定的環境參數包括海水水深(D)、水溫(WT)、鹽度(S)、pH值、溶解氧(DO)、化學需氧量(COD)、磷酸鹽(DIP)、無機氮(DIN)、懸浮物(SS)、表層沉積物底質類型及中值粒徑(MD)。
1.2 數據處理
1.2.1 群落結構及優勢種計算
使用Shannon-Wiener多樣性指數(H′)[17]、Pielous均勻度指數(J)[17]和Margalef物種豐富度指數(d)[17]分析生物群落結構。使用物種優勢度指數(Dominance index,Y)判定優勢種[18]。計算公式如下:
(1)
(2)
d=(S-1)/log2N,
(3)
Y=Pi×F,
(4)
式中,Pi=ni/N(ni是第i個物種的個體數,N是全部物種的個體數);S為種類數;F為該種的出現頻率,當Y>0.02時[19],即為該海域底棲動物優勢種。H′值等于0,說明無底棲生物,即嚴重污染;0-1為重污染;1-2為中度污染;2-3為輕度污染;大于3為清潔[17]。
1.2.2 環境因子相關分析
使用SPSS 18.0軟件,進行單因素方差分析(One-way ANOVA),分析各季節環境要素的差異性;采用Canoco 5.0軟件對大型底棲動物優勢種生物密度和環境因子進行典范對應分析(CCA),物種矩陣經lg(x+1)轉換[20],探究環境因子對大型底棲動物優勢種生物密度變化的影響。
1.2.3 底棲生境質量評價
AMBI和M-AMBI采用AMBI V5.0軟件計算,先除去非大型底棲無脊椎動物的種類,其余大型底棲動物的生態分組根據文獻[10,21]規定執行。AMBI值為0.0-7.0,數值越低表示生態健康狀況越好;M-AMBI值為0.00-1.00,數值越高表示生態健康狀況越好。AMBI和M-AMBI的評價標準[10,22]詳見表1。

表1 AMBI和M-AMBI擾動等級、生態環境質量狀況標準
2 結果與分析
2.1 群落結構
2.1.1 種類組成與優勢種
調查共發現82種底棲生物,包括扁形動物、刺胞動物、紐形動物、環節動物、軟體動物、節肢動物、棘皮動物、半索動物和脊索動物9個門類,其中環節動物多毛類29種、節肢動物甲殼類29種、軟體動物18種,以上3個門類種數占總種數的92.68%,是女姑口鄰近海域主要的大型底棲動物類群,其他各門類均1種。春季(4月)采集到的大型底棲動物種類較多,為67種,其中環節動物27種,軟體動物15種,節肢動物21種,扁形動物、刺胞動物、紐形動物、棘皮動物各1種。秋季(11月)采集到大型底棲動物46種,其中環節動物21種,軟體動物7種,節肢動物15種,紐形動物、尾索動物、脊索動物各1種。
優勢種主要為環節動物多毛類和軟體動物,兩個季節均表現出明顯優勢的種為寡鰓齒吻沙蠶(Nephtysoligobranchia)、寡節甘吻沙蠶(Glycindegurjanovae)和菲律賓蛤仔。其他優勢種均為環節動物多毛類及小型貝類,膠州灣女姑口鄰近海域大型底棲動物有小型化趨勢。節肢動物甲殼類在本調查中共發現29種,其中秋季15種,其密度占秋季總密度的12.03%,密度占比最高的種為長尾亮鉤蝦(Photislongicaudata)和極地蚤鉤蝦(Pontocratesaltamarinus),兩種密度占比均為2.95%,站位出現率均為41.67%;春季21種,其密度占春季總密度的13.78%,密度占比最高的種類為東方長眼蝦(Ogyridesorientalis)(密度占比2.23%,站位出現率33.33%)。雖然甲殼類是種類組成的主要類群,但是密度占比未超過14%,站位出現率未超過45%,優勢度較低,并沒有出現甲殼類優勢種。調查海域大型底棲動物優勢種及優勢度指數見表2。
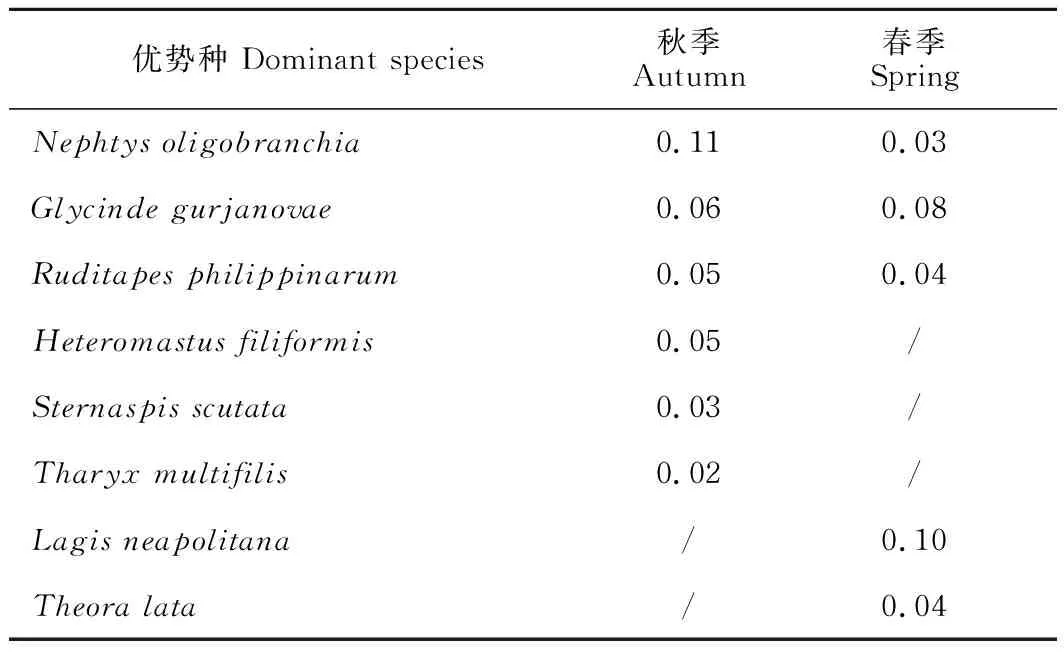
表2 秋季和春季調查海域大型底棲動物優勢種及優勢度指數
2.1.2 生物密度與生物量分布
調查海域大型底棲動物平均生物密度為665.83 ind./m2,平均生物量為358.65 g/m2。調查海域2018年秋季平均生物密度為485.00 ind./m2,平均生物量為235.80 g/m2;2019年春季平均生物密度為846.67 ind./m2,平均生物量為481.60 g/m2。由圖2可知,秋季生物密度高值區主要位于紅島灘涂附近海域,生物量高值區位于紅島外側海域;春季生物密度及生物量的高值區主要集中在墨水河口附近海域,墨水河口2號站春季調查發現大量菲律賓蛤仔,因此生物量生物密度高。在季節變化方面,春季生物密度、生物量較高,而且種類組成分析發現,春季軟體動物和節肢動物種類數量明顯增多。
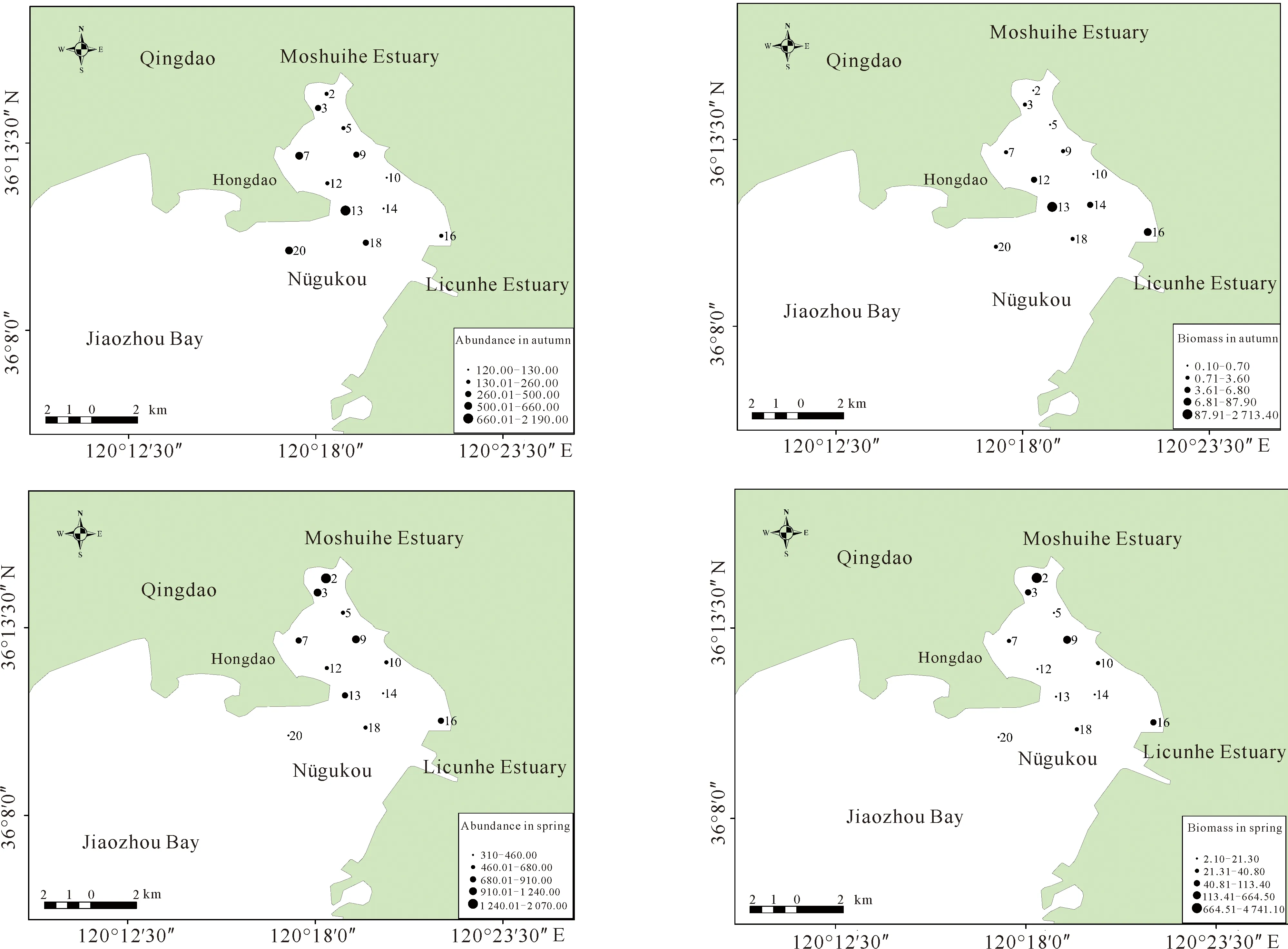
圖2 秋季和春季調查海域大型底棲動物生物密度、生物量分布
2.1.3 群落多樣性特征
由表3可知,調查海域大型底棲動物春季Shannon-Wiener多樣性指數(H′)平均值高于秋季,兩季節的平均多樣性指數為2.77,說明調查海域大型底棲動物受到輕度污染;Pielous均勻度指數(J)略低于秋季,Margalef豐富度指數(d)高于秋季。
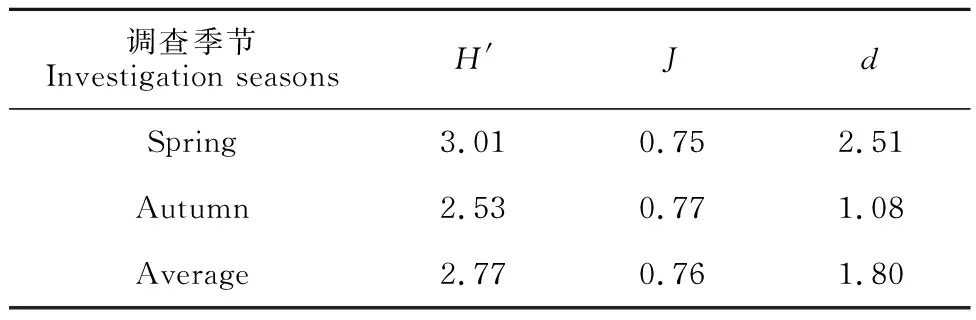
表3 秋季和春季調查海域大型底棲動物群落多樣性指數
2.2 大型底棲動物與環境因子的相關性分析
調查海域水深為2.0-7.5 m,平均水深為4.17 m,呈現梯度遞增趨勢;12個調查站位的底質為黏土質粉砂;水溫(WT)、鹽度(S)、溶解氧(DO)、化學需氧量(COD)、磷酸鹽(DIP)、懸浮物(SS)季節間呈極顯著差異(P<0.01);無機氮(DIN)季節間呈顯著差異(P<0.05);季節間pH值與中值粒徑(MD)無顯著差異。
采用《海水水質標準》[23]對調查海域水質進行評價。調查海域秋季和春季海水中pH值、溶解氧均滿足一類海水水質標準要求,化學需氧量、磷酸鹽、無機氮均超一類海水水質標準要求。其中,秋季和春季海水化學需氧量全部符合二類水質標準;秋季調查發現磷酸鹽和無機氮都超四類水質標準,超標率均為75.0%;春季調查發現磷酸鹽和無機氮都超四類水質標準,超標率分別為20.0%和50.0%。
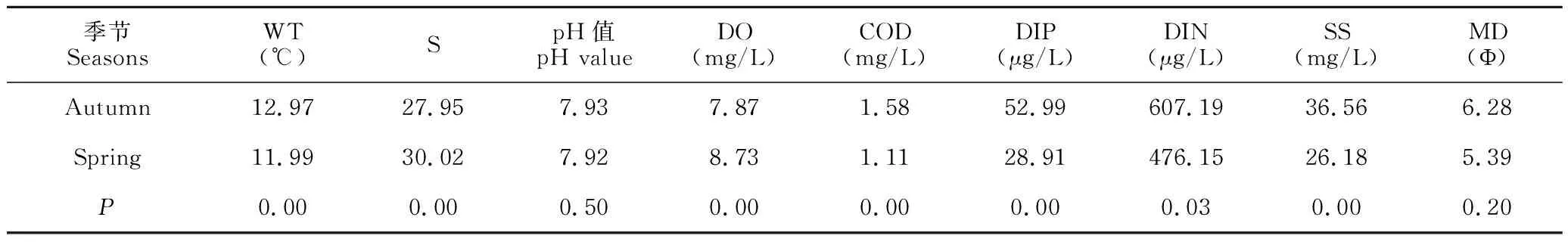
表4 秋季與春季調查海域各環境因子平均值
由圖3可知,兩個排序軸的特征值分別為0.58和0.17,種類與環境因子排序軸的相關系數為0.952和0.754,排序可以較好地反映底棲動物與環境因子間的關系。鹽度(S)與第一排序軸呈最大正相關(0.717),水深(CD)與第一排序軸呈第二正相關(0.534),水溫(WT)與第一排序軸呈最大負相關(-0.754)。說明影響膠州灣女姑口大型底棲動物分布的主要驅動因子有鹽度、水溫。
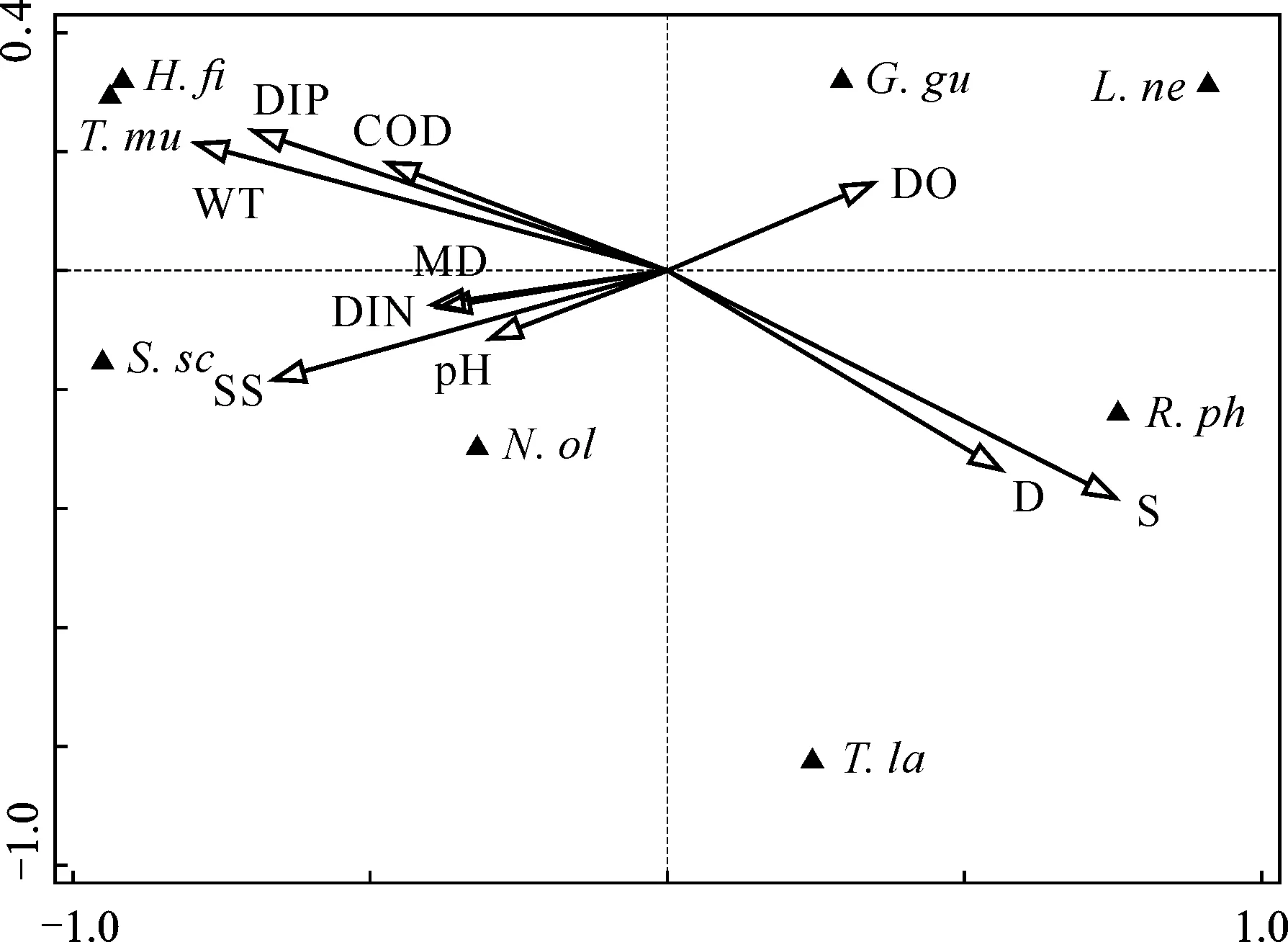
N.ol:Nephtys oligobranchia;G.gu:Glycinde gurjanovae;R.ph:Ruditapes philippinarum;H.fi:Heteromastus filiformis;S.sc:Sternaspis sculata;T.mu:Tharyx multifilis;L.ne:Lagis neapolitana;T.la:Theora lata
2.3 AMBI及M-AMBI結果
2018年秋季 12個調查站位AMBI為1.41-3.16,均為輕度擾動;M-AMBI為0.46-0.92,站位13生態環境質量處于中等級,其他站位為優或者良(表5)。2019年春季12個調查站位AMBI為1.58-3.70,10個調查站位為輕度擾動,2個站位(站位14、站位3)為中度擾動(表5)。站位14第二機會種(EG Ⅳ)占比為77.8%,站位3第二機會種(EG Ⅳ)占比為62.3%。M-AMBI為0.50-0.91,站位14底棲生態環境質量處于中等級,其他站位屬于良或者優(表5)。
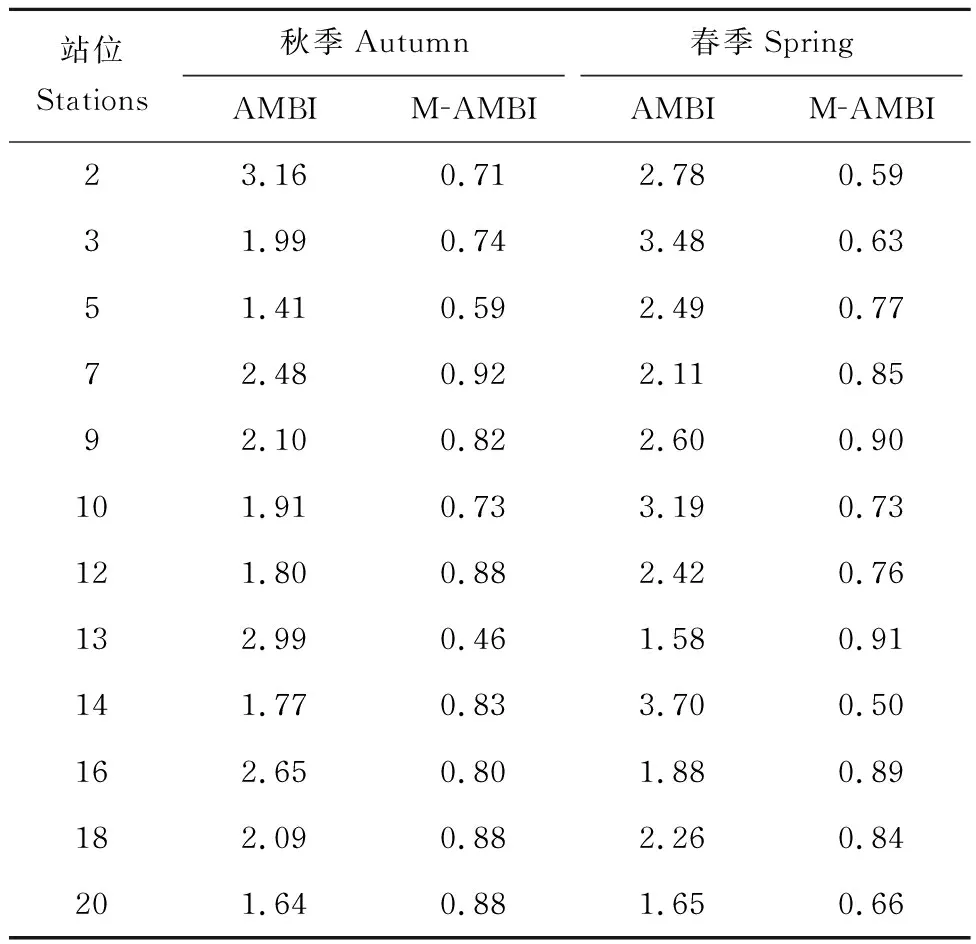
表5 秋季與春季調查海域AMBI及M-AMBI
3 討論
3.1 大型底棲動物與環境要素的關系
本研究結果表明,對膠州灣女姑口附近海域大型底棲動物分布特征造成影響的主要環境因子為鹽度和水溫。這與陳曉娟等[24]和畢洪生[25]對膠州灣大型底棲動物的研究結論相似,即水溫與鹽度影響膠州灣大型底棲動物的生物密度分布。崔雯瑤等[26]對膠州灣2018年春季數據進行分析,其調查時間與本研究調查時間相近,同樣發現水溫與鹽度會影響膠州灣大型底棲動物生物密度分布,而其他環境因子無顯著相關關系。膠州灣屬于暖溫帶水域,水深較淺,水溫變化劇烈,水溫影響底棲動物的生長和發育[27],從而影響生物的種類數量、生物量和分布范圍[28]。以往調查中發現,鹽度對底棲生物的影響主要集中在河口附近區域,該處因河流淡水的注入,尤其是在雨季,大量注入的淡水引起鹽度發生較大的變化,從而影響該處的群落組成和生物密度[29,30]。
3.2 膠州灣女姑口附近海域底棲生態環境健康狀況
膠州灣女姑口附近海域大型底棲動物秋季Shannon-Wiener多樣性指數為2.53,春季Shannon-Wiener多樣性指數為3.01。李新正等[31]2001年調查發現,膠州灣底棲動物的物種多樣性指數春季、秋季平均值分別為4.59和4.76;王金寶等[32]2005-2009年膠州灣調查,春季Shannon-Wiener多樣性指數為0.46-3.66,秋季Shannon-Wiener多樣性指數0.86-3.61;隋吉星等[33]2010年對膠州灣中部海域調查顯示,膠州灣大型底棲動物春季和秋季Shannon-Wiener多樣性指數較低,平均值分別為2.59和2.50;全秋梅等[11]2016-2017年調查膠州灣大型底棲動物春季和秋季Shannon-Wiener多樣性指數分別為2.61和2.34;丁敬坤等[16]2020年調查膠州灣發現,女姑口附近站位Shannon-Wiener多樣性指數為1.67;近20年調查顯示,膠州灣大型底棲動物的Shannon-Wiener多樣性指數逐漸降低,從側面說明該區域大型底棲動物受到輕度污染。
本次調查結果顯示,膠州灣女姑口附近海域12個站位AMBI秋季為1.41-3.16,春季為1.58-3.70,說明調查海域皆受到不同程度的干擾,所有站次均處于輕度擾動以上;M-AMBI秋季為0.46-0.92,春季為0.50-0.91,說明調查海域部分站位的生態環境健康狀況處于中等狀態;Shannon-Wiener多樣性指數(H′)、AMBI及M-AMBI評價結果基本一致。本調查結果與丁敬坤等[16]評價結果相似,即膠州灣海域底棲生態系統受到中等程度的干擾。
菲律賓蛤仔是膠州灣養殖的主要種類,在菲律賓蛤仔較多的站位,群落Shannon-Wiener多樣性指數和Pielous均勻度指數降低,小個體的多毛類,如那不勒斯膜帽蟲(L.neapolitana)、多絲獨毛蟲(T.multifilis)等,生物密度升高,此類生物多為機會種,因此導致AMBI變大、M-AMBI變小,王振鐘[34]調查也得出相似結論。養殖過程中的營養鹽排放、養殖生物的逃逸、疾病的傳播等給海洋生態系統帶來較大影響[35]。菲律賓蛤仔自身覓食及挖掘活動對養殖海域的環境造成一定的擾動,同時其捕獲過程對表層的沉積環境造成劇烈的物理擾動,對大型底棲動物的生活環境造成一定的破壞[16]。本次調查受中度擾動的站位位于菲律賓蛤仔養殖用海附近,該海域的大型底棲動物的健康狀況可能受到養殖活動干擾。
當群落受到中等程度的污染擾動時,個體較大的種的優勢度被削弱,生物量占優勢的大個體消失,因此在數量上占優勢的種是個體較小的種[35],膠州灣歷史調查中已發現存在大型底棲動物小型化現象[36-38],畢洪生等[36]曾在1991-1995年對膠州灣的底棲動物進行連續監測,發現20世紀90年代與80年代相比,小型底泥食性種類豐度上升,主要種類為絲異須蟲(H.filiformis)和方格獨鰓蟲(Tharyxtesselata);王洪法等[37]2009年在膠州灣調查發現優勢種為多毛類動物的不倒翁蟲(S.scutata)和軟體動物菲律賓蛤仔;全秋梅等[11]在膠州灣調查中發現優勢種以多毛類為主,如絲異須蟲、索沙蠶科(Lumbrineriiae)等。在渤海大型底棲動物調查中也發現小型化現象,萊州灣自20世紀90年代以來大型底棲動物群落結構已發生變化,總體呈現小型化趨勢,主要是小型多毛類動物和甲殼類動物取代大個體的棘皮動物和軟體動物[39]。本次膠州灣女姑口調查中,環節動物多毛類種類共29種,在秋季總密度中占比49.65%,在春季占比50.29%,優勢種為菲律賓蛤仔、寡鰓齒吻沙蠶和寡節甘吻沙蠶,其他優勢種均為環節動物多毛類及小型貝類,與歷史調查結果相近,膠州灣女姑口附近種類組成與優勢種呈小型化現象。
造成大型底棲動物小型化的原因主要有水體富營養化、過度捕撈及大型底拖網使用等[40,41]。由于近岸水域通常受到較嚴重的環境脅迫,近海污染、徑流攜帶大量的有機質,以及水流和波浪對海底沉積物和底棲生物群落的影響較大,導致近岸的群落主要由一些小型多毛類耐受種組成,這些種多為機會種[27]。本次調查區域位于膠州灣東北部,常年平均水深4 m,水動力交換較弱,墨水河、白沙河及李村河入海口在本調查區域附近,調查發現重金屬高值區主要位于東部李村河口[42,43]。青島市生態環境狀況公報顯示,2018年青島市膠州灣優良水質面積比例為73.7%,李村河、墨水河和大沽河入海口附近海域水質較差[44]。2019年膠州灣東北部海域水質較差,主要污染物為無機氮[45]。本次調查秋季磷酸鹽和無機氮都超四類水質標準,超標率均為75.0%;春季磷酸鹽和無機氮都超四類水質標準,超標率分別為20.0%和50.0%。磷酸鹽和無機氮超標易導致水體富營養化,水體富營養化對大型底棲動物多樣性產生顯著的負面影響[11],富營養化加速大型底棲動物的種類更新頻率,導致群落組成變化以及群落功能多樣性降低[46]。本次調查優勢種多絲獨毛蟲、絲異須蟲、那不勒斯膜帽蟲在AMBI生態組中為第二級機會種(EG Ⅳ),機會種的高生物密度和高出現率導致部分站位生態環境質量評價較差。大型底棲動物群落結構發生變動,出現小型化,在一定程度上反映了近年調查海域受到污染和擾動[39]。
綜上所述,養殖和污染可能導致膠州灣女姑口附近海域大型底棲動物小型化,該海域的底棲生態環境健康狀況受到一定擾動。對此,建議加強對膠州灣東北部近岸海域環境管理,探索有效的綜合治理措施,使膠州灣海洋生態系統得以持續健康地發展。
4 結論
本次膠州灣女姑口鄰近海域調查共獲得大型底棲動物82種,以環節動物、軟體動物、甲殼動物為主,棘皮動物等其他動物占比很小。兩個調查季節中均表現出明顯優勢的種為寡鰓齒吻沙蠶、寡節甘吻沙蠶和菲律賓蛤仔,但是其他優勢種均為環節動物多毛類及小型貝類,說明膠州灣女姑口鄰近海域大型底棲動物有小型化趨勢,高經濟價值種在生物密度和生物量上也有一定程度的下降。典范對應分析的結果顯示,鹽度、水溫是影響大型底棲動物分布特征的主要因子。綜合大型底棲動物群落的Shannon-Wiener多樣性指數、AMBI及M-AMBI分析結果,膠州灣女姑口鄰近海域底棲生態系統受到輕度干擾,紅島灘涂附近海洋生態環境質量狀況處于一般水平,其他地區處于優良或高等水平,養殖和污染可能導致膠州灣女姑口附近海域大型底棲動物群落結構呈小型化趨勢,并導致該海域的底棲生態環境健康狀況受到擾動。膠州灣東北部女姑口海域是人類活動及工業生活密集區域,加強該海域的生態環境保護具有重要意義。
猜你喜歡
我愛學·笑話與口才(2025年3期)2025-02-24 00:00:00
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
趣味(語文)(2020年3期)2020-07-27 01:42:46
航空世界(2020年10期)2020-01-19 14:36:20
作文與考試·初中版(2017年12期)2017-04-19 20:26:27
中國衛生(2015年2期)2015-11-12 13:13:54
