傷害誘導普通白木香與奇楠種質沉香中倍半萜類成分及早期倍半萜合酶基因表達的差異分析△
2022-01-28 13:28:46呂菲菲楊云孫佩文馮劍劉培衛劉洋洋徐艷紅魏建和
中國現代中藥 2021年12期
關鍵詞:差異
呂菲菲,楊云,孫佩文,馮劍,劉培衛,劉洋洋,徐艷紅*,魏建和,*
1.中國醫學科學院 北京協和醫學院 藥用植物研究所 海南分所/海南省南藥資源保護與開發重點實驗室/國家中醫藥管理局沉香可持續利用重點研究室,海南 海口 570311;2.中國醫學科學院 北京協和醫學院 藥用植物研究所,北京 100193
沉香是 瑞香科(Thymelaeaceae)沉 香屬(AquilariaLam.)或擬沉香屬植物傷害誘導防御反應形成的含樹脂木材,是中國的傳統名貴中藥,廣泛應用于中成藥、藏藥和日本漢方藥中[1-2]。因具有獨特的芳香氣味,沉香也被廣泛用于熏香香料行業,是高級香水香料的定香劑[3]。奇楠被認為是品質優良的沉香,外表油潤光滑,香味醇厚,味微苦、麻、辣、清涼[4]。中國產的奇楠沉香以野生采挖為主[5],香農在野外采香的過程中發現某些白木香樹經輕微傷害后可以持續地形成奇楠沉香,但這種優良性狀很難通過有性繁殖而延續。近年來,香農將采挖的野生奇楠沉香樹木進行嫁接擴繁,獲得了可穩定遺傳的奇楠種質,通過鉆孔法傷害成功地生產了奇楠沉香,由奇楠種質生產的奇楠沉香已流通于市場。
倍半萜是沉香的主要成分之一,只有受傷害后才會在莖干、枝條和根內大量合成,其組成和含量決定了沉香的藥用價值和芳香氣味[6]。奇楠種質生產的奇楠沉香顏色重、油性大、香味獨特,明顯不同于普通白木香生產的沉香,可見2 種種質生產的沉香中倍半萜組分和含量有所不同,但倍半萜合成差異的分子調控機制尚缺乏研究。倍半萜合酶是白木香傷害誘導合成倍半萜的關鍵催化酶,其活性受基因表達調控[7]。健康白木香中倍半萜合酶幾乎不表達,外界傷害刺激后,才會被激活誘導表達[8],倍半萜合酶基因表達一定程度上決定了白木香受到傷害后倍半萜合成的種類和含量。基于此,本研究以一種生產上的奇楠種質和普通白木香種質為研究對象,分析2 種種質所結沉香倍半萜組分的差異,以及傷害早期顯著響應的7 個倍半萜合酶基因(AsTPS2、AsTPS13、AsTPS14、AsTPS17、AsTPS18、AsTPS20、AsTPS23)誘導表達模式,初步分析這2 種種質傷害誘導形成倍半萜差異的機制,也為沉香優良結香種質的鑒定和篩選、結香分子機制的解析提供依據。
1 材料
1.1 試藥
奇楠種質及其所結沉香取自廣東省茂名市電白區觀珠鎮,經中國醫學科學院藥用植物研究所海南分所采用DNA boarding 技術鑒定為白木香Aquilaria sinensis(Lour.)Gilg。普通白木香種質及其所結沉香取自海南省海口市中國醫學科學院藥用植物研究所海口研發中心。
乙醚(西隴科學股份有限公司);液氮(純度>99.99%,海南佳騰化工氣體有限公司);EASY-spin plus RN38型植物RNA快速提取試劑盒(北京艾德萊生物科技有限公司);TransScript?First-Strand cDNA Synthesis SuperMix、TransStart?Top Green qPCR SuperMix、瓊脂糖均購于北京全式金生物技術有限公司。
1.2 儀器
XPE 105 型電子天平(梅特勒-托利多公司);7890A型氣相色譜-質譜儀(安捷倫公司);Nano Drop 2000C 超微量分光光度計(賽默飛世爾科技公司);LightCycler?96 型實時熒光定量聚合酶鏈式反應(qRT-PCR)儀(霍夫曼·羅氏公司);SB25-12DTDN型超聲波清洗機(寧波新芝生物科技股份有限公司);MiniVac Beta 型真空離心濃縮干燥機(Labogene 公司);ChemiDoc?XRS+型凝膠成像儀(Bio-Rad公司)。
2 方法
2.1 倍半萜提取與測定
沉香倍半萜提取參考文獻[9]方法,準確稱取結香2 年沉香0.5 g,液氮研磨成粉,向粉末中加入乙醚5 mL,冰浴超聲提取30 min,靜置5 min,收集上清液備用。向沉淀中再次加入乙醚5 mL,重復提取2 次,合并3 次收集的上清液,真空旋轉干燥,向干燥物中加入乙醚1 mL 溶解,經0.22 μm 濾膜濾過后,氣相色譜-質譜法(GC-MS)測定化學組分。
GC 分析條件:毛細管柱為HP-5MS 5% Phenyl Silox(30 m×250 μm,0.25 μm);載氣為氦氣;柱體積流量為1.0 mL·min-1,不分流;進樣口溫度為240 ℃;升溫程序:初始柱溫60 ℃,保持2 min,5 ℃·min-1升至250 ℃,保持10 min,共運行50 min。
質譜條件:電子轟擊離子源(EI);電子能為70 eV;離子源溫度為230 ℃;四級桿溫度為150 ℃;質量掃描范圍為m/z50~300。
2.2 莖干總RNA提取和cDNA合成
選取3 年生健康莖干,直徑為(1.2±0.2)cm,對莖干進行全斷干傷害處理,并分別在傷害后0、2、6、24 h 截取傷口處2 cm 莖干(傷害誘導結香早期0~24 h 是傷害信號傳遞、防御反應啟動、基因誘導表達相對最活躍階段),立即投入液氮中冷凍,-80 ℃保存備用。
全斷干傷害處理莖干經液氮研磨成粉,按照試劑盒說明書提取莖干總RNA,1.5%瓊脂糖凝膠電泳檢測RNA 完整性,測定RNA 濃度。取所提取的RNA 約500 ng,根據要求反轉錄合成cDNA第一條鏈,作為qRT-PCR模版。測定cDNA濃度。
2.3 qRT-PCR檢測基因表達
根據本課題組白木香全基因組注釋的倍半萜合酶基因序列,采用Primer 5.0 軟件設計特異性引物(表1),由廣州艾基生物有限公司合成,所有引物經PCR擴增后,產物由廣州艾基生物有限公司測序驗證。
qRT-PCR 檢測基因表達量:20 μL PCR 體系包含Mix enzyme 10 μL,10 μmol·L-1的上下游引物各1 μL,反轉錄所得cDNA 1 μL 和雙蒸水(ddH2O)7 μL。qRT-PCR 擴增程序:95 ℃預變性10 min,95 ℃變性10 s,按不同基因引物退火溫度退火15 s(表1),72 ℃延伸20 s,共40 個循環,72 ℃延伸10 min。實驗設定3 個重復。以三磷酸甘油醛脫氫酶(GAPDH)為內參[10],將qRT-PCR 所得的樣品Ct值利用2-ΔΔCt方法轉化為基因相對表達量值。使用SPSS 1.1.0 數據統計軟件對檢測結果進行單樣本t檢驗顯著性分析,結果以(-x±s)表示,P<0.05 為差異有統計學意義。

表1 沉香倍半萜合酶基因引物和擴增片段
3 結果與分析
3.1 倍半萜類成分差異
GC-MS 測定后,將測得的化合物與NIST 數據庫比對,對匹配度>75%的21個倍半萜及其衍生物進行對比分析,發現普通白木香所結的沉香中有17個,奇楠沉香中有16 個,在奇楠沉香中檢測到的香橙烯、檸檬烯、α-馬欖烯、石竹烯4個倍半萜類化合物在普通白木香結的沉香中沒有檢測到,而普通白木香所結沉香中存在的蛇麻烯、(-)-馬兜鈴烯、別香橙烯、長葉醛、香橙烯氧化物1 也沒有在奇楠沉香中檢測到,可見2 種種質所結沉香倍半萜組分差異較大。對兩者共有成分相對含量進行對比分析,發現倍半萜及其衍生物在兩者中的相對含量也有所不同,如普通白木香所結沉香中α-檀香醇的相對含量是奇楠沉香的6 倍左右,而奇楠沉香中α-愈創木烯相對含量是普通白木香所結沉香的2 倍有余(表2),綜上,奇楠沉香和普通白木香所結的沉香中倍半萜組分和含量差異明顯。
表2 普通白木香所結沉香和奇楠沉香中倍半萜及其衍生物對比(, n=3)

表2 普通白木香所結沉香和奇楠沉香中倍半萜及其衍生物對比(, n=3)
注:根據Duncan′s多區間統計分析,上標不同字母代表各組間P<0.05。
3.2 倍半萜合酶基因檢測
提取莖干的總RNA,經瓊脂糖電泳檢測明顯有18S 和28S 2 條帶,條帶清晰,無拖尾,28S/18S 約為1.5(圖1),說明提取的莖干總RNA 質量較好,經反轉后可用于后續qRT-PCR檢測。
圖1 2種白木香莖干總RNA凝膠電泳
根據白木香全基因組(PRJNA524272)及前期白木香倍半萜合酶基因誘導表達模式[11]分析發現,傷害誘導結香早期顯著響應的倍半萜合酶基因主要為AsTPS2、AsTPS13、AsTPS14、AsTPS17、AsTPS18、AsTPS20、AsTPS23。經PCR 擴增,擴增條帶大小與設計引物得到的片段大小基本一致(圖2)。將PCR產物測序,所得結果進行BLAST 比對,所有序列比對結果均符合全基因組注釋結果,即所有擴增產物為倍半萜合酶的DNA序列。
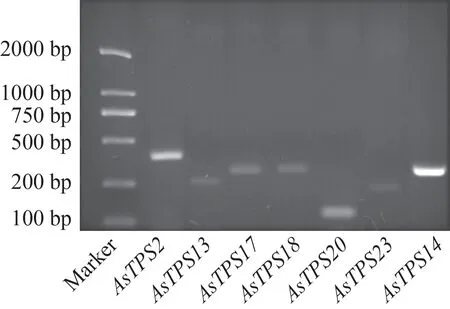
圖2 7個倍半萜合酶基因在傷害處理白木香莖干中PCR擴增條帶
3.3 普通白木香倍半萜合酶基因傷害早期誘導表達
對已驗證序列的7個傷害早期誘導表達倍半萜合酶基因進行了qRT-PCR 檢測,發現普通白木香AsTPS2、AsTPS13、AsTPS14、AsTPS17、AsTPS18、AsTPS20、AsTPS23基因均響應于傷害誘導,傷害24 h 內基因表達量均先上升,2 h 達到最大值后下降,誘導表達最大相對倍數均在幾十倍到幾百倍,誘導表達強度明顯(圖3),說 明AsTPS2、AsTPS13、AsTPS14、AsTPS17、AsTPS18、AsTPS20、AsTPS23均強烈響應于普通白木香的早期傷害誘導,且誘導表達模式基本一致,呈先上升后下降的態勢。
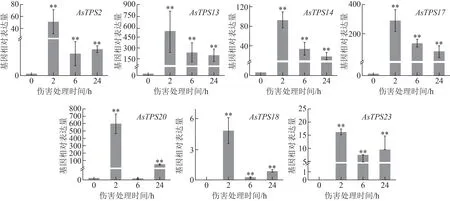
圖3 不同傷害處理時間對普通白木香種質中7個倍半萜合酶基因誘導表達水平的影響(, n=3)
3.4 奇楠種質倍半萜合酶基因傷害早期誘導表達
經qRT-PCR 檢測,發現除AsTPS18基因外,奇楠 種質中的AsTPS2、AsTPS13、AsTPS14、AsTPS17、AsTPS20、AsTPS23基因也都響應于傷害刺激,但各基因響應傷害誘導的表達模式不同,AsTPS2、AsTPS13、AsTPS14、AsTPS17和AsTPS23基因在傷害誘導24 h 內表達逐漸增強,基因表達量呈遞增態勢,AsTPS20則呈現先上升后下降的表達趨勢,傷害誘導6 h 表達量最高,這6 個倍半萜合酶基因傷害誘導24 h 內最大表達相對倍數集中于十幾倍到幾十倍,與普通白木香相比誘導響應強度較低(圖4)。AsTPS18基因在傷害誘導24 h 內表達不明顯,6 h 達到最大表達量,僅為3.60,遠低于普通白木香中的誘導相對表達量,其在奇楠種質中可能不是早期響應傷害的倍半萜合酶基因。說明除AsTPS18外,其余6 個倍半萜合酶基因在奇楠種質中均在傷害誘導結香早期有所響應,但與普通白木香中的基因誘導表達模式不一致,誘導表達強度相對普通白木香較低。
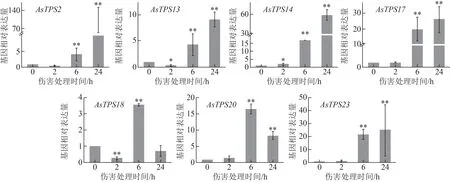
圖4 不同傷害處理時間對奇楠種質中7個倍半萜合酶基因表達水平的影響(, n=3)
3.5 健康奇楠種質與普通白木香倍半萜合酶基因表達分析
為了進一步探索2 種種質的差異,對比了奇楠種質和普通白木香健康莖干內倍半萜合酶基因表達情況(圖5),發現普通白木香種質中響應早期傷害的7 個倍半萜合酶基因在健康莖干中幾乎不表達,而奇楠種質中除AsTPS2與AsTPS20外,AsTPS13、AsTPS14、AsTPS17、AsTPS23和AsTPS18表達明顯,且高于健康白木香,說明健康的奇楠種質和普通白木香中倍半萜合酶基因的表達也存在明顯差異。
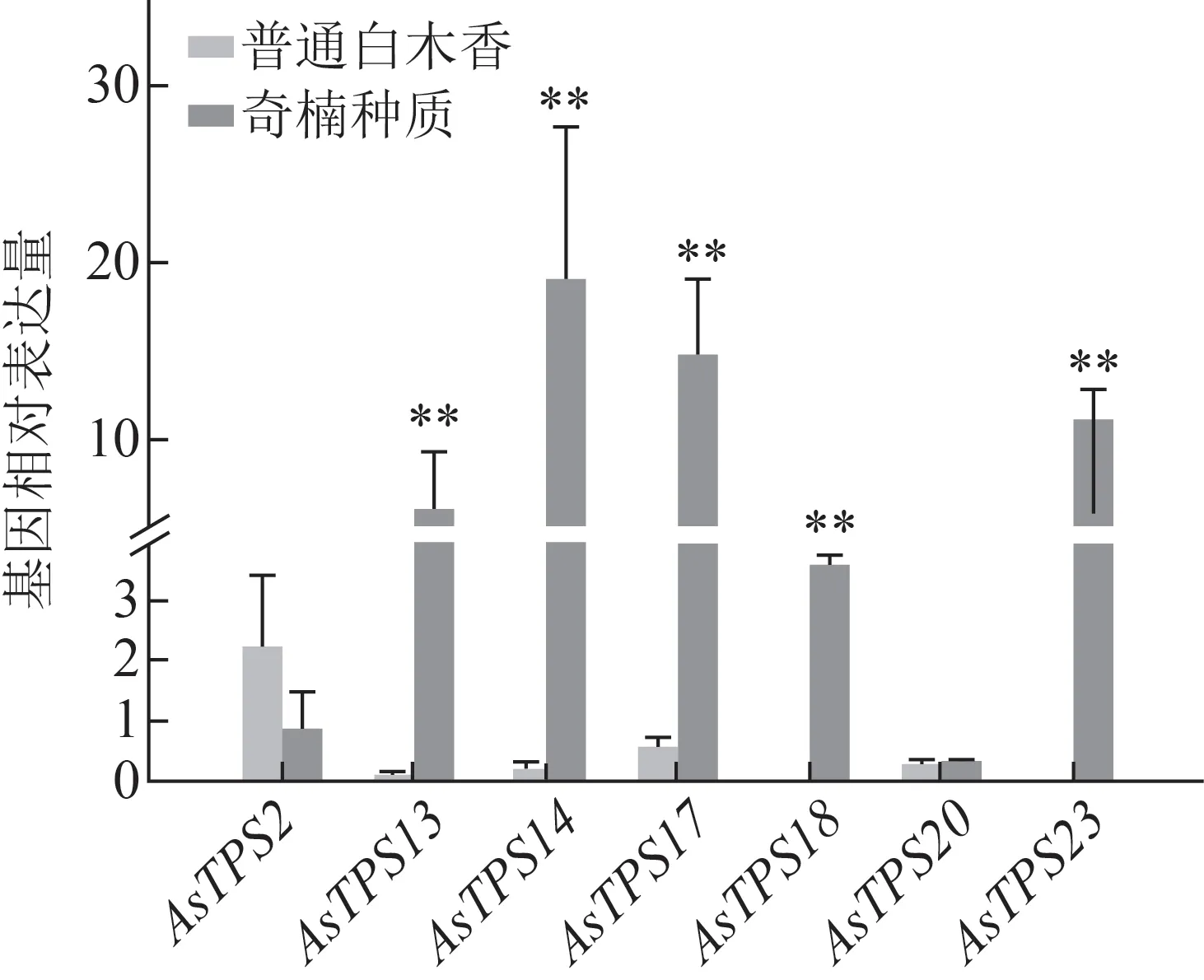
圖5 奇楠種質和普通白木香健康莖干內7個倍半萜合酶基因表達對比(, n=3)
4 討論
倍半萜是沉香的主要成分之一,其含量和組分在一定程度上決定了沉香的含油量和香味[4,12]。目前,奇楠種質生產的奇楠沉香含油量高,香味醇厚,品質明顯高于普通白木香種質所結沉香。為了探索2種種質結香差異及篩選優良種質,本研究選取一種市面上大量推廣嫁接的奇楠種質,發現其生產的奇楠沉香與普通白木香所結沉香的倍半萜組分有所不同,且共有組分含量差異也較大,說明在相同傷害下,奇楠種質和普通白木香種質誘導合成沉香倍半萜的分子調控機制存在差異,兩者倍半萜組分和含量的差異可能是奇楠沉香品質優于普通白木香種質的關鍵原因。該發現與大麻不同品種間萜類組分不同的研究結果相一致[13]。
倍半萜是沉香屬植物受傷害后形成的次生代謝產物,植物中多種因子協同調控其合成[14-17]。倍半萜合酶就是倍半萜合成的關鍵催化酶,可催化法呢烯焦磷酸(farnesyl pyrophosphate,FPP)形成不同的倍半萜骨架[7]。本研究發現普通白木香種質中傷害誘導結香早期顯著誘導高表達的AsTPS2、AsTPS13、AsTPS14、AsTPS17、AsTPS20、AsTPS23基因在奇楠種質中響應強度較低,而AsTPS18基因在奇楠種質中幾乎不響應于早期傷害誘導,可見倍半萜合酶基因誘導表達機制在奇楠種質和普通白木香種質中差異顯著,表明2 種種質傷害后啟動防御反應的機制有所不同,致使誘導合成倍半萜的組分和含量不同。此外,還發現AsTPS13、AsTPS14、AsTPS17、AsTPS23和AsTPS18基因在健康奇楠種質中表達量顯著高于健康白木香,說明奇楠種質中倍半萜合酶基因表達更為敏感,這可能是盛傳的奇楠種質不傷害也能產生奇楠沉香的原因。倍半萜合酶基因的差異表達機制是奇楠種質和普通白木香種質傷害誘導結香性能差異的關鍵調控機制,本研究首次從基因表達層面解析2 種種質結香性能差異,對其深入研究有助于完善沉香形成機制,差異表達倍半萜合酶的篩選和論證將為新的分子標記開發和沉香優良結香種質的鑒定提供參考。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
