牛磺酸通過調(diào)控AKT/GSK-3β通路抑制結(jié)直腸癌細胞侵襲轉(zhuǎn)移
2022-01-27 03:29:42陳春香萬慧芳李淑英鐘小菊萬福生
南昌大學學報(理科版) 2021年6期
關(guān)鍵詞:實驗
陳春香,萬慧芳,李淑英,鐘小菊,萬福生,*
(1.南昌大學撫州醫(yī)學院生化與分子生物學教研室,江西 撫州 344000;2.南昌大學醫(yī)學實驗教學中心,江西 南昌 330031;3.南昌大學基礎(chǔ)醫(yī)學院生化與分子生物學教研室,江西 南昌 330006)
結(jié)直腸癌(Colorectal cancer,CRC)是人類最常見的胃腸道惡性腫瘤之一,發(fā)病率和死亡率高,目前是世界范圍內(nèi)癌癥相關(guān)死亡的第二大原因[1-2]。近年來由于飲食結(jié)構(gòu)和生活方式的改變,我國CRC的發(fā)病率保持了快速上升的趨勢,是我國第三大常見惡性腫瘤[3-4]。研究[5-7]表明,CRC高發(fā)病率和高死亡率的主要原因是其具有難以控制的持續(xù)增殖和侵襲轉(zhuǎn)移能力,最終縮短了患者的生存時間。糖原合酶激酶-3β(GSK-3β)是一種獨特的絲氨酸/蘇氨酸蛋白激酶,既是由NF-κb介導的癌細胞生存調(diào)節(jié)因子,又是多種細胞功能的復雜調(diào)節(jié)因子,涉及的疾病廣泛,包括神經(jīng)退行性變、炎癥、纖維化、非胰島素依賴性糖尿病及癌癥等[8-11]。
最近的研究認為GSK-3β可作為多種不同類型癌癥的一個潛在的治療靶點[12],在控制癌細胞侵襲和遷移方面發(fā)揮重要作用。GSK-3β是AKT/GSK-3β信號通路調(diào)控上皮-間質(zhì)轉(zhuǎn)化(EMT)的關(guān)鍵分子,與結(jié)直腸癌的侵襲轉(zhuǎn)移密切相關(guān)[13-14]。牛磺酸(Taurine,Tau)是細胞內(nèi)含量最豐富的游離氨基酸之一,具有較廣泛的生理功能及藥理作用。我們前期研究表明Tau能誘導結(jié)直腸癌細胞凋亡,抑制增殖[15],但Tau對結(jié)直腸癌侵襲轉(zhuǎn)移及AKT/GSK-3β/β-Catenin通路的影響如何尚不清楚。本文旨在觀察GSK-3β在Tau抑制SW480和HT29細胞侵襲轉(zhuǎn)移中的作用,探討Tau抑制結(jié)直腸癌侵襲轉(zhuǎn)移的作用靶點及分子機制。
1 材料和方法
1.1 細胞系和細胞培養(yǎng)
人結(jié)腸癌細胞系SW480、HT29購于中國科學院典型培養(yǎng)物保藏委員會細胞庫。SW480細胞常規(guī)培養(yǎng)于含10%胎牛血清的1640培養(yǎng)基上,HT29細胞培養(yǎng)于含10%胎牛血清的DMEM完全培養(yǎng)液中,保存于37 ℃,5% CO2培養(yǎng)箱中。
1.2 主要試劑和材料
牛磺酸(Sigma公司);胎牛血清、RPMI-1640培養(yǎng)基、胰蛋白酶、細胞裂解液及二甲亞風(DMSO)均購于Solarbio公司;抗AKT(cat.#40576)多抗購自SAB公司,p-AKT(cat.No.#3787)多抗購自中國上海細胞信息技術(shù)有限公司,PTEN單抗(cat.Ab32199),E-cadherin單抗(Cat.Ab1416),N-cadherin單抗(Cat.Ab18203),Vimentin單抗(Cat.Ab92547);MMP-2(cat.Ab6302),MMP-7(cat.Ab37150),MMP-9多抗(cat.Ab38898),β-Catenin多抗(cat.Ab92547),GSK-3β單抗(cat.Ab32391),p-GSK-3β-Ser9(Cat.ab75814)多抗及β-actin多抗(cat.20536-1-AP)均購自Abcam公司;辣根過氧化物酶(HRP)標記山羊抗兔IgG購于北京中杉金橋生物技術(shù)有限公司。CCK8試劑(上海東仁化學科技有限公司)。康寧基底膜基質(zhì)(356234)購自康寧公司(中國上海)。Transwell小室(8 μmoL)購自Corning公司,JetPRIME轉(zhuǎn)染試劑購自上海英捷生物有限公司。質(zhì)粒p-EGFP-GSK-3β及空載體p-EGFP-N1購買于上海諾百生物科技有限公司。
1.3 Transwell分析細胞遷移和侵襲實驗
Transwell小室細胞遷移實驗:收集對數(shù)生長期的SW480和HT29細胞,實驗分為正常對照組(Control)、4個Tau組(40,80,160,200 mmoL)。將生長良好的細胞用不含血清的培養(yǎng)基處理12 h,然后用含0.25% EDTA的胰蛋白酶消化,在不含血清的上室中以1×105/孔的密度放置。Transwell裝置下腔填充600μl RPMI 1640或DMEM,DMEM中添加10%胎牛血清,含不同濃度的Tau。培養(yǎng)48 h后,用棉拭子從上孔取出無創(chuàng)細胞,4%多聚甲醛固定20 min,1%結(jié)晶紫烘干20 min,洗3次。小室干燥后在倒置顯微鏡下觀察并使用圖像采集系統(tǒng)選取上、中、下、左、右5個視野,于100倍光鏡下拍照,計數(shù)其平均穿膜細胞數(shù)。
細胞侵襲實驗:將Matrigel膠分別與不含血清的H-DMEM和RPMI-1640基礎(chǔ)培養(yǎng)基按1:8的比例混合均勻。將Matrigel膠和培養(yǎng)基混合好后,取該混合物60 μL鋪于已預冷的小室中,將小室置24孔板中,在37℃過夜凝固成膠狀。然后接種,接種于小室的細胞數(shù)變?yōu)?×105個/孔,其余步驟與Transwell小室遷移實驗相同,計算平均細胞數(shù),即侵襲細胞數(shù)。實驗重復3次。
1.4 細胞轉(zhuǎn)染和Tau處理
收集對數(shù)生長期的SW480和HT29細胞,實驗分為以下5組:①正常對照組(Control):細胞于37℃、5%CO2正常培養(yǎng)48 h;②Tau(80 mmol·L-1):加入終濃度為80 mmol·L-1的Tau,正常培養(yǎng);③空載體對照組(p-EGFP-N1):轉(zhuǎn)染NC-shRNA后正常培養(yǎng);④p-EGFP-GSK-3β轉(zhuǎn)染組:轉(zhuǎn)染p-EGFP-GSK-3β,6 h后換液,正常培養(yǎng)48 h;⑤p-EGFP-GSK-3β轉(zhuǎn)染+Tau(80 mmol·L-1)聯(lián)合處理組:轉(zhuǎn)染p-EGFP-GSK-3β,6 h后換液,加入終濃度為80 mmol·L-1的Tau,培養(yǎng)48 h。轉(zhuǎn)染過程按jetPRIME轉(zhuǎn)染試劑說明書進行,基本步驟如下:將2 μg DNA溶于200 μL的jetPRIME緩沖液中,漩渦震蕩混勻。往上述混合液中加入4 μL jetPRIME轉(zhuǎn)染試劑,漩渦混勻10 s,簡單地上下顛倒。室溫孵育10 min后,每孔加入200 μl的轉(zhuǎn)染復合物到含有完全培養(yǎng)基的細胞中,且分布均勻,并將其放回培養(yǎng)箱中培養(yǎng)48 h,然后進行其他實驗。實驗重復3次。
1.5 CRC細胞劃痕愈合實驗
收集對數(shù)生長期的SW480和HT29細胞,實驗分組與1.4細胞轉(zhuǎn)染和Tau處理相同。用Marker筆在6孔板的外底部橫著畫4條線,然后收集對數(shù)生長期的細胞接種于6孔板中,5×105個/孔,貼壁生長12 h后用10 μL移液器的槍頭在6孔板內(nèi)底部垂直Marker筆的畫線而豎直劃痕4條;用2 mL PBS洗去懸浮細胞,加入2 mL/孔新鮮培養(yǎng)液并在光學顯微鏡下拍照(40倍),以此時為0 h;接著按實驗分組,分別加入不同濃度Tau培養(yǎng)處理細胞12 h、24h,在固定時間點,同一條件下拍下劃痕照片并記錄細胞遷移情況。最后,用Image Pro Plus 6.0軟件統(tǒng)計平均值,計算細胞遷移率。細胞遷移率(%)=(0h劃痕距離-24h劃痕距離)/0 h劃痕距離×100%。實驗重復3次。
1.6 Western blot檢測細胞中蛋白水平
收集經(jīng)Tau處理48 h的細胞,在冰上用細胞裂解液進行裂解,12 000 r·min-1離心(4℃)10 min,取上清液。采用Pierce BCA ProteinAssayKit試劑盒檢測蛋白濃度。按20 μg蛋白上樣量將蛋白樣品與上樣緩沖液(5×)按4:1的體積比例混合,沸水中煮5 min,冷卻后上樣。經(jīng)10%SDS-PAGE并半干電泳轉(zhuǎn)至PVDF膜上,5%脫脂奶粉封閉2 h后,分別加入PTEN、AKT、p-AKT、GSK-3β、p-GSK-3β、E-cadherin、Vimentin、β-catenin及MMP2/7等抗體,并4 ℃孵育過夜。次日,在室溫下加入二抗孵育2h,ECL化學發(fā)光法顯色,X線膠片曝光成像,掃描儀掃描蛋白條帶。實驗重復3次。
1.7 統(tǒng)計處理
所有結(jié)果被描述為均數(shù)加減標準偏差(SD)。采用SPSS 19.0軟件進行雙尾配對t檢驗,以檢測各組間測量變量的顯著性差異,P<0.05為有統(tǒng)計學意義。
2 結(jié)果
2.1 Tau對人結(jié)直腸癌細胞侵襲和遷移的抑制作用
通過Transwell小室實驗檢測了SW480和HT29細胞的侵襲和遷移情況(見圖1)。結(jié)果顯示,對照組SW480細胞的侵襲能力明顯大于HT29細胞,Tau對SW480和HT29細胞侵襲和遷移的抑制作用均隨著藥物濃度的增加而增強(P<0.05)。經(jīng)Tau處理48 h后SW480細胞的侵襲能力也明顯比HT29細胞強;

注:a為Tau作用48 h的細胞侵襲結(jié)果(n=3),b為Tau作用48 h的細胞遷移結(jié)果(n=3),c為細胞侵襲統(tǒng)計結(jié)果圖,d為細胞遷移統(tǒng)計結(jié)果圖。
2.2 Tau對人結(jié)直腸癌細胞PTEN/AKT/GSK-3β通路的影響
為了探討Tau抑制CRC細胞生長、侵襲和遷移的分子機制,我們觀測了Tau對SW480和HT29細胞中PTEN/AKT/GSK-3β通路相關(guān)蛋白表達變化(見圖2)。結(jié)果顯示,當Tau作用細胞48 h后,對CRC細胞中PTEN,AKT,p-AKT,GSK-3β及p-Gsk-3β蛋白水平有不同程度的影響,與對照組比較,Tau有一定的濃度依賴性上調(diào)SW480和HT29細胞中PTEN,GSK-3β及p-GSK-3β蛋白水平(P<0.05),下調(diào)AKT,p-AKT Ser473蛋白水平;在SW480細胞中Tau對GSK-3β,AKT和p-AKT Ser473的作用要大于HT29細胞,而對PTEN和p-GSK-3β的作用要小于HT29細胞。結(jié)果提示Tau可提高CRC細胞內(nèi)PTEN、GSK-3β表達及p-GSK-3β水平,下調(diào)AKT,p-AKT Ser473蛋白水平。

注:A為Tau作用48h后SW480細胞結(jié)果(n=3),B為Tau作用48hHT29細胞結(jié)果(n=3);
2.3 質(zhì)粒轉(zhuǎn)染效果
結(jié)果如圖3所示,SW480和HT29細胞經(jīng)轉(zhuǎn)染48h后,p-EGFP-N1和p-EGFP-GSK-3β組細胞在熒光顯微鏡下可觀察到綠色熒光,而Control組未見熒光,說明這兩組有熒光蛋白導入;NC-shRNA組也顯綠色熒光,說明有RNA干擾片段導入SW480和HT29細胞中。

注:圖左邊有綠色熒光蛋白是在熒光顯微鏡下的結(jié)果(顯綠色熒光);右邊呈黃色的是普通光鏡下結(jié)果。
2.4 GSK-3β過表達對結(jié)直腸癌細胞侵襲和遷移的影響
為探討GSK-3β在Tau抑制CRC細胞侵襲和轉(zhuǎn)移中的作用,我們觀察了GSK-3β過表達在Tau在抑制SW480和HT29細胞侵襲和轉(zhuǎn)移中的影響。Transwell小室實驗結(jié)果(見圖4)顯示,與Control或p-EGFP-NC組比較,Tau組的細胞侵襲和遷移的數(shù)量呈顯著性減少(P<0.01);與p-EGFP-NC組相比,p-EGFP-GSK-3β組的細胞侵襲和遷移數(shù)量有顯著性減少(P<0.01);與p-EGFP-GSK-3β或Tau組比較,p-EGFP-GSK-3β+Tau組的侵襲和遷移細胞數(shù)量有顯著性降低(P<0.01)。劃痕愈合實驗結(jié)果(圖5)與Transwell小室實驗結(jié)果基本一致,細胞遷移的能力被GSK-3β顯著抑制,但pEGFP-GSK-3β組的劃痕愈合速度比Tau組慢;與pEGFP-GSK-3β組比較,pEGFP-GSK-3β+Tau組的愈合速度顯著降低(P<0.01)。結(jié)果提示GSK-3β能增強Tau對CRC細胞侵襲和遷移的抑制作用。

注:A為SW480細胞的結(jié)果,B為HT29細胞的結(jié)果;*P<0.05,**P<0.01 vs control;#P<0.05,##P<0.01 vs p-EGFP-NC;&P<0.05,&&P<0.01 vs p-EGFP-GSK-3β or Tau 80 mmoL group.Tau的濃度是80 mmoL·L-1,作用時間48 h;n=3

注:A為SW480細胞劃痕愈合實驗結(jié)果(處理48 h),B為HT29細胞劃痕愈合實驗結(jié)果(處理48 h),C為SW480細胞統(tǒng)計結(jié)果圖,D為HT29細胞統(tǒng)計結(jié)果圖.*P<0.05,**P<0.01 vs control;#P<0.05,##P<0.01 vs p-EGFP-NC;&P<0.05,&&P<0.01 vs p-EGFP-GSK-3β or Tau 80 mmoL group。
2.5 GSK-3β過表達對EMT、MMPs及AKT/GSK-3β/β-Catenin通路相關(guān)蛋白表達的影響
為探討GSK-3β對SW480細胞中EMT、MMPs及AKT/GSK-3β/β-Catenin通路相關(guān)蛋白表達的作用。我們的實驗結(jié)果(見圖6)發(fā)現(xiàn),與p-EGFP-NC組比較,p-EGFP-GSK-3β組的E-Cadherin、GSK-3β蛋白表達有顯著升高(P<0.01),而N-Cadherin,Vimentin,β-Catenin,MMP2,MMP7和AKT蛋白表達均有顯著降低(P<0.01或P<0.05);與Tau組或p-EGFP-GSK-3β組比較,p-EGFP-GSK-3β+Tau組的E-Cadherin,GSK-3β蛋白表達有顯著升高(P<0.01),而N-Cadherin、Vimentin、β-Catenin、MMP2、MMP7和AKT蛋白表達均有顯著降低(P<0.01)。結(jié)果表明GSK-3β蛋白能增強Tau抑制SW480細胞EMT過程,下調(diào)細胞內(nèi)MMP2、MMP7及β-Catenin含量。
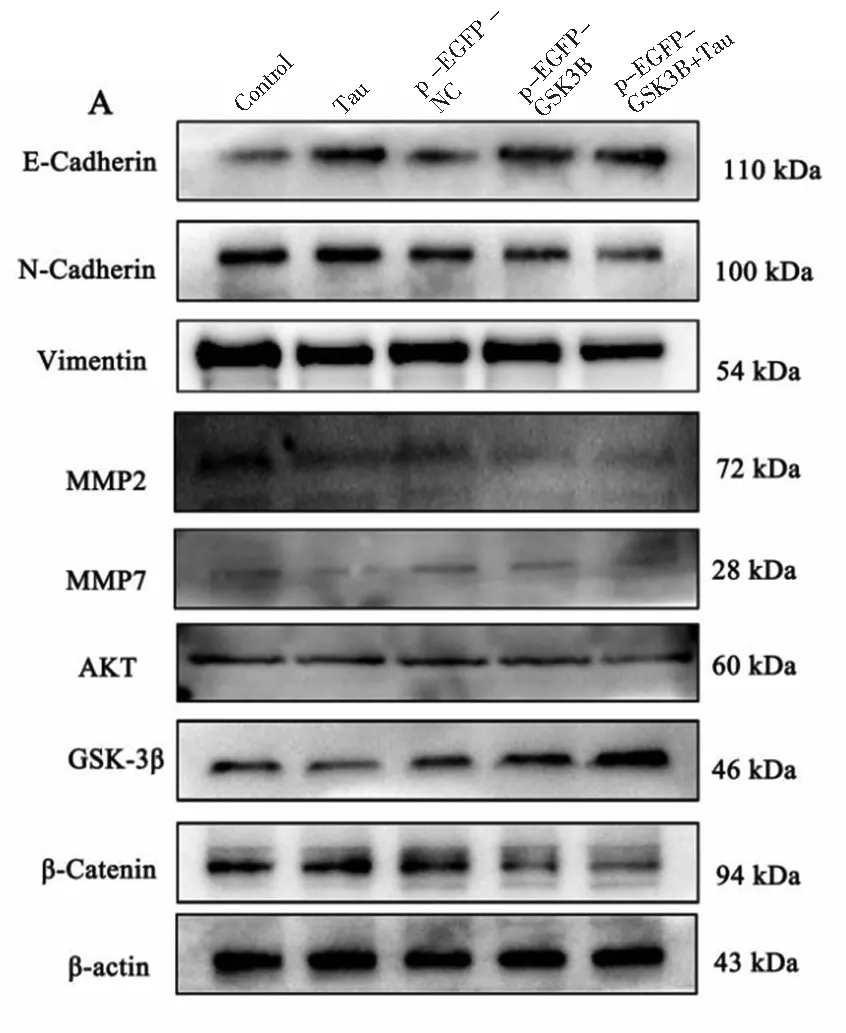
注:A為Western blotting結(jié)果,B,C和D為結(jié)果統(tǒng)計處理圖;*P<0.05,**P<0.01 vs control;##P<0.01
3 討論
牛磺酸(又名2-氨基乙磺酸)是一種不用于蛋白質(zhì)合成的含硫氨基酸,在哺乳動物中被稱為半必需氨基酸,主要由肝臟和腎臟產(chǎn)生,是哺乳動物組織中含量最多的游離氨基酸[16]。牛磺酸具有許多基本的生物學作用,它存在于包括視網(wǎng)膜、大腦、心臟和胎盤等不同的器官,對心血管功能、骨骼肌、視網(wǎng)膜及中樞神經(jīng)系統(tǒng)發(fā)育和功能至關(guān)重要[17-19]。在人體內(nèi)各種器官和組織成分中起著重要的生理和病理作用,包括滲透壓調(diào)節(jié)、膜穩(wěn)定、鈣水平的調(diào)節(jié)、能量代謝、基因表達、抗氧化、抗凋亡、抗炎及抗脂質(zhì)活性等[20-22]。牛磺酸的抗氧化作用已被發(fā)現(xiàn)可以影響細胞增殖、炎癥和膠原蛋白沉積。近年來不斷有研究表明[23-26]Tau具有較強的抗腫瘤作用,主要表現(xiàn)為誘導癌細胞凋亡、自噬及對化療藥物的增效減毒作用等,但牛磺酸抗腫瘤作用的機制尚未弄清。據(jù)報道牛磺酸與順鉑聯(lián)合應用可下調(diào)基質(zhì)金屬蛋白酶2(matrix metalloproteinase 2,MMP-2),抑制膠質(zhì)瘤細胞侵襲轉(zhuǎn)移,也可抑制前列腺癌EMT過程[23]。本文實驗結(jié)果顯示,牛磺酸不僅對結(jié)直腸癌侵襲遷移有抑制作用(圖1),且可提高CRC細胞內(nèi)PTEN、GSK-3β表達及p-GSK-3β水平,下調(diào)AKT,p-AKT Ser473蛋白水平(圖2)。結(jié)果表明牛磺酸可下調(diào)p-AKT Ser473蛋白水平,提高CRC細胞內(nèi)GSK-3β,而GSK-3β能增強牛磺酸對CRC細胞侵襲遷移的抑制作用(見圖4和5)。最近有研究[27]結(jié)果顯示,在使用偶氮氧基甲烷(AOM)/硫酸鈉(DSS)誘導的結(jié)腸癌小鼠模型上,牛磺酸能顯著提高凋亡標記物、cleaved caspase-9和腫瘤抑制蛋白PTEN的水平,證明牛磺酸在體內(nèi)有顯著降低致癌性的作用。類似研究[28-29]結(jié)果是使用二甲基苯[a]蒽(DMBA)誘導的大鼠乳腺癌發(fā)生作為乳腺癌模型。給藥牛磺酸可顯著降低DMBA誘導的大鼠乳腺癌發(fā)生率(從80%降至40%)。但牛磺酸在體內(nèi)是否有抑制癌細胞侵襲遷移的作用,目前罕見有關(guān)研究報道。
另有研究結(jié)果[30-31]表明,牛磺酸還可以作為一種化學保護劑對抗金屬離子Cr(VI)誘導的氧化損傷,并可能被用于緩解這種過渡金屬離子的毒性作用;牛磺酸也可以降低赭曲霉素A(OTA)的毒性作用,其機制是牛磺酸通過降低LC3-II蛋白表達和GFP-LC3點的熒光強度來降低OTA誘導的自噬,減少細胞凋亡,維持細胞穩(wěn)態(tài)。簡而言之,牛磺酸可緩解OTA誘導的凋亡,抑制pk15細胞自噬,這些作用也與牛磺酸的抗腫瘤作用有關(guān)。
GSK-3β屬于絲氨酸/蘇氨酸蛋白激酶,最初被描述為一種參與糖原代謝的關(guān)鍵酶。近年來研究發(fā)現(xiàn)GSK-3β是一種多功能蛋白,是磷脂酰肌醇-3激酶(PI3K)下游作用的主要介質(zhì),協(xié)調(diào)多種細胞內(nèi)信號,控制細胞對外源性刺激的反應,對細胞內(nèi)信號通路至關(guān)重要,參與多種細胞活動[8],如調(diào)節(jié)心肌細胞生長[9]、細胞遷移、葡萄糖代謝調(diào)節(jié)、炎癥反應及細胞凋亡[10]等,GSK-3β是AKT信號傳導途徑的重要下游靶標,AKT通過Ser473處被磷酸化激活和使GSK-3β失活來促進細胞侵襲轉(zhuǎn)移。PTEN則是AKT的上游信號分子,對AKT起著負調(diào)控作用。GSK-3β參與β-catenin的降解,是調(diào)節(jié)細胞內(nèi)β-catenin含量、分布和功能的重要因子[32];胞內(nèi)β-catenin水平的降低可能是GSK-3β-介導的在β-catenin Ser 33/37和Thr 41上的磷酸化所致,β-catenin的磷酸化形式由蛋白酶體降解的β-TrCP泛素E3連接酶泛素化。GSK-3β的失活會導致胞內(nèi)β-catenin的累積,促進腫瘤侵襲轉(zhuǎn)移[33]。本文實驗結(jié)果表明,牛磺酸通過調(diào)控AKT/GSK-3β通路,抑制CRC細胞EMT過程和下調(diào)MMP2、MMP7及β-Catenin表達,最終抑制CRC細胞侵襲遷移。
猜你喜歡
作文·小學低年級(2025年2期)2025-02-13 00:00:00
小雪花·小學生快樂作文(2024年11期)2024-12-31 00:00:00
作文·小學低年級(2024年2期)2024-04-29 00:00:00
作文·小學低年級(2023年3期)2023-04-29 00:00:00
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
小主人報(2022年4期)2022-08-09 08:52:06
中學生數(shù)理化·中考版(2022年11期)2022-02-16 07:01:20
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
發(fā)明與創(chuàng)新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
