根際微生物組中細菌趨化系統的生態功能
2022-01-16 05:35:42常晶晶田春杰
生態學報 2021年24期
孫 雨,常晶晶,田春杰
中國科學院東北地理與農業生態研究所黑土區農業生態重點實驗室, 長春 130102
根際作為土壤與植物間物質交換的活躍界面,其物理、化學以及生物學特性與周圍土壤都存在著顯著差異[1]。在根際這一特殊的生態環境中,與植物根系相關的微生物種類和數量巨大[2]。根際微生物在植物的生長發育以及抵抗逆境脅迫方面都扮演著重要的角色[3],包括促進植物對營養元素的獲取及利用[4-5]、調控植物的生長和環境適應性[6-7]等。根際微生物組與非根際土壤微生物組在物種多樣性和豐富度方面均存在顯著差異[8],其中植物根系的調控作用是決定根際微生物組群落結構的重要因素之一[9]。因此,深入揭示植物對根際微生物組的選擇機制不僅可以在理論方面增進對植物-微生物互作關系的認識,還能在實際應用中指導根際微生物組的改良與重組,對未來農業生產具有重大意義。
植物調控根際微生物組的重要策略之一是通過根系分泌化合物,選擇性招募特定根際微生物[10-11]。在這一過程中,土壤微生物通過何種機制來感應根系分泌物,響應植物的招募?細菌的趨化反應給出了一種答案。土壤中存在一類趨化細菌,能夠利用自身獨特的趨化系統感應其生存環境中存在的化學物質濃度梯度,通過一系列信號轉導過程調控細菌的運動方向,使細菌趨近吸引物而趨離排斥物[12-13]。趨化細菌在沒有化合物濃度梯度的環境中,其運動方向是隨機的,向前直線運動數秒后停下“翻筋斗”,然后再以不同的方向繼續直線運動,以“直線運動-轉變方向-直線運動”這一過程循環往復[14];當周圍環境中存在吸引物(碳源等營養物)的濃度梯度時,趨化細菌轉變方向的頻率降低,能夠延長向高濃度區直線運動的時間,總體表現為細菌趨近吸引物[15]。
細菌的趨化反應是其對碳源、能源競爭的一種表現,體現了細菌的主動適應性。Wuichet和Zhulin[16]基于全基因組數據,對近400株細菌的趨化系統進行了分析,推測絕大部分的細菌可能都具有趨化性。但是,目前針對細菌趨化系統的研究主要集中在分子層面對其信號轉導和作用機理的解析,缺少從生態學的角度揭示其在植物根際微生物群落結構和功能中的表現和作用。本文以根際微生物組中的細菌趨化系統為核心,針對國內外最新研究進展,從以下三個方面進行綜述,提出了研究根際細菌趨化系統生態功能的具體方向,旨在闡明根際細菌趨化系統的生態功能,增進對根際微生物組的募集組裝過程的認知,以及為根際微生物組的重組構建提供理論依據。
1 細菌趨化系統的組成和信號轉導機制
細菌的趨化反應是在長期進化過程中形成的“趨利避害”的本能,研究人員以大腸桿菌(Escherichiacoli)[17]、芽孢桿菌(Bacillusspp.)[18]、假單胞菌(Pseudomonasspp.)[19]等為對象,對細菌的趨化反應進行了系統深入的研究。細菌的趨化系統是原核生物中最為復雜的信號轉導系統之一,雖然不同細菌中趨化系統的組成有一定差異,但是其信號轉導及調控機制在所有細菌中是相對保守的[15]。
細菌趨化系統由以下一系列不同功能的蛋白組成:趨化受體蛋白Mcps(Methyl-accepting Chemotaxis Proteins)、支架蛋白CheW/CheV、組氨酸激酶CheA、反應調節蛋白CheY、甲基酯酶CheB、甲基轉移酶CheR、磷酸酶CheC/CheX/CheZ、以及脫酰胺酶CheD[16]。趨化系統的信號轉導及調控機制如下(圖1):Mcps負責結合胞外趨化信號,并通過支架蛋白CheW/V與CheA相連;Mcps結合信號物質后調控CheA的組氨酸激酶活性,進而將信號向下游的CheY傳遞;被CheA磷酸化的CheY(CheY-P)通過鞭毛蛋白FliM與鞭毛-馬達復合體相互作用,調控細菌的運動方向。共同構成“適應系統”的CheB和CheR互相競爭,調控Mcps的甲基化狀態從而調節其對信號物質的結合親和力,不斷“重置”細菌趨化系統的靈敏度。磷酸酶CheC/CheX/CheZ能夠將CheY-P去磷酸化,使CheY持續接受來自CheA的磷酸基團。脫酰胺酶CheD則能夠調節CheC的磷酸酶活性,并作用于Mcps。這一套精密運作的趨化系統使細菌能夠靈敏感應胞外信號物質的濃度梯度,適應周圍環境的變化。
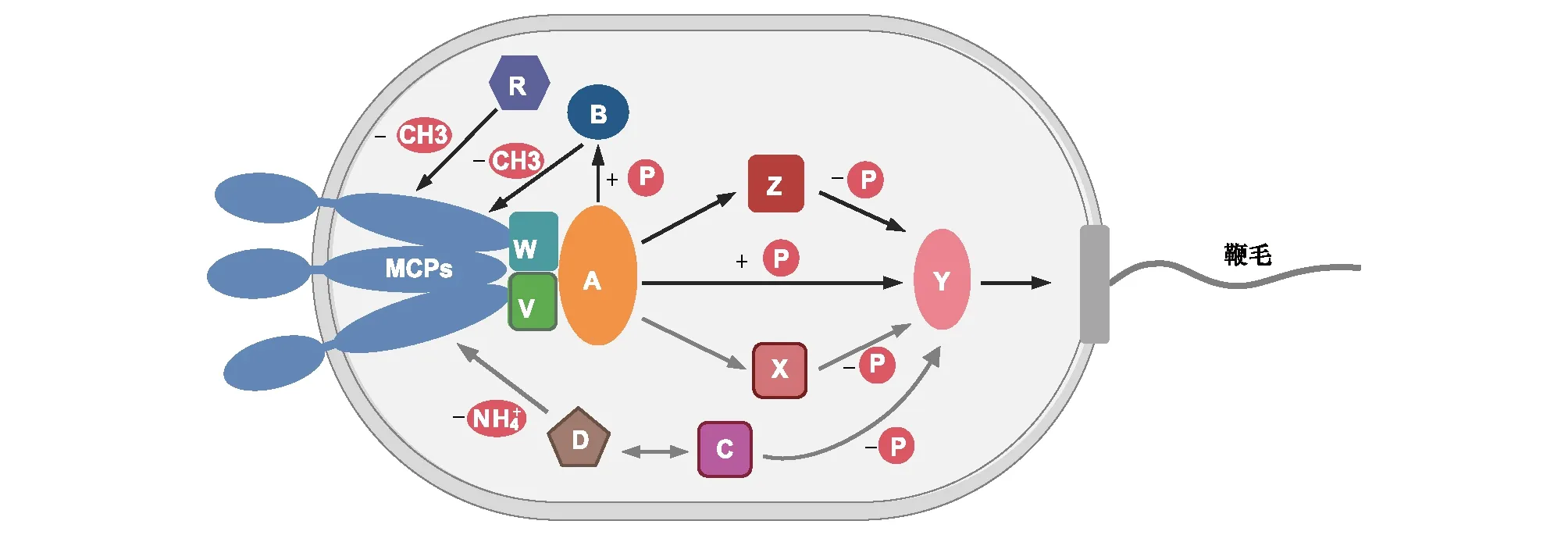
圖1 細菌趨化系統信號轉導模式圖Fig.1 A summary model illustrates the organization of bacterial chemotaxis systemsMcps: 趨化受體蛋白;V/W: 支架蛋白CheV 和CheW;A: 組氨酸激酶CheA;B: 甲基酯酶CheB;R: 甲基轉移酶CheR;D: 脫酰胺酶CheD;C/X/Z: 磷酸酶CheC/CheX/CheZ;Y: 反應調節蛋白CheY;P: 磷酸基團;CH3: 甲基; 銨根;本模式圖在大腸桿菌趨化系統的基礎上,集中了當前細菌趨化系統研究的模式菌(包括芽孢桿菌、假單胞菌等)中已鑒定的所有蛋白組分,帶有輪廓線的組分與灰色箭頭標注的反應不存在于大腸桿菌中
2 細菌趨化系統介導植物與微生物間相互作用
近年來,大量的研究證明了細菌趨化系統能夠介導植物與微生物相互作用。在根瘤菌和豆科植物共生體系的建立過程中,豆科植物根系分泌類黃酮等化合物,根瘤菌則能夠利用趨化系統感應這一類信號分子從而靠近植物根系表面,Cooper[20]因此總結:根瘤菌對豆科植物根系分泌的信號分子的趨化反應是二者建立共生關系的第一步。此外,在根際促生菌與植物的互作中細菌趨化系統也發揮重要作用。Allard-Massicotte等[21]發現,模式植物擬南芥的根系分泌物能夠誘導芽孢桿菌B.subtilis產生趨化反應進而吸引其達到根系,并且B.subtilis對擬南芥根系的成功定殖依賴于其完整的趨化系統;芽孢桿菌B.velezensis對作物玉米、黃瓜根系分泌物的趨化反應調控了其對宿主的根系定殖[22-23]。此外,另外一類典型的根際促生菌假單胞菌如P.fluorescens、P.putida等的趨化系統在其對宿主植物玉米、番茄根系定殖中的重要作用也得到了驗證[24-25]。除了根瘤菌和根際促生菌等有益菌,一些植物病原菌也能夠利用自身的趨化系統感應植物根系分泌物,通過趨化反應靠近植物根系并起始侵染致病過程。Hida等[26]研究發現,番茄根系分泌物能夠誘導青枯雷爾氏菌Ralstoniapseudosolanacearum產生趨化反應,促進其向番茄根際的運動及對番茄根系的定殖。
上述的研究主要針對單一菌株與植物的互作,在群落水平上,也有研究證明了細菌趨化系統在介導植物與微生物互作中的重要作用。Kamutando等[27]對入侵樹種銀荊的研究發現,其根際土壤中細菌趨化系統相關基因的豐度顯著高于非根際土壤;Xu等[28]對全球范圍內六個大洲的柑橘根際微生物組的研究表明,細菌趨化系統相關基因在柑橘根際核心微生物組中顯著富集。但是,當前在群落水平上對細菌趨化系統的認識仍有局限,將趨化系統作為一個整體考慮是可以理解的,然而趨化系統作為一個復雜的信號轉導系統,不同組分的進化速率和變異程度可能并不完全同步,所以針對信號系統各組分的細致研究對深入揭示其生態功能至關重要。
3 植物對根際細菌趨化系統的選擇及其調控機制
研究表明,不同植物根際細菌趨化系統的分布存在一定差異,主要體現在豐度和多樣性方面。在趨化基因豐度方面,Mendes等[29]對大豆根際微生物組的研究發現,抗鐮刀菌根腐病大豆的根際趨化基因豐度顯著高于易染病大豆,并且趨化基因的豐度與根際細菌群落結構的復雜度呈正相關。在趨化基因多樣性方面, Buchan等[30]采用了“T-RFLP”(Terminal restriction fragment length polymorphism,末端限制性片段長度多態性)技術對小麥和豇豆根際細菌趨化系統中核心基因cheA的序列多樣性進行了分析,發現根際趨化細菌中cheA序列的多樣性低于非根際細菌,且在兩種植物根際間差異顯著。但是上述研究受當時技術所限,缺少對全部趨化基因的系統分析,尤其是趨化系統中受外界信號調控的組分,包括結合外界信號物質的趨化受體mcp,以及調控趨化受體結合物質靈敏度的cheB和cheR。由于趨化系統介導了植物與微生物間的互作,根際微生物組中趨化基因的豐度能夠一定程度上體現植物與微生物間互作關系的強弱,趨化基因的多樣性則體現了植物對細菌趨化系統的特異性選擇。當前,對根際趨化相關基因的豐度和多樣性,以及其體現的植物對趨化系統的選擇缺少認識,仍需進行深入探索。隨著高通量測序技術的發展,當前利用宏基因組分析結合傳統熒光定量PCR手段已能夠實現對特定信號通路中相關基因的豐度和多樣性進行分析。
細菌趨化系統能夠感應植物的根系分泌物,已有的研究結果表明,多種不同類型的根系分泌物包括蘋果酸、草酸等有機酸[31-32],組氨酸、精氨酸等氨基酸[33-34],甘露糖、半乳糖等糖類[23, 35],以及酚酸[36]、苯并惡唑嗪酮[37]等次級代謝產物都能夠誘導根際趨化細菌產生趨化反應。Zhang等[38]通過HPLC鑒定了黃瓜和香蕉根系分泌物中的有機酸,并分別檢測了芽孢桿菌屬細菌B.amyloliquefaciens和B.subtilis對兩種植物根系分泌物的趨化反應,發現黃瓜根系分泌物中特有的檸檬酸能夠誘導B.amyloliquefaciens和Bacillussubtilis產生趨化反應,而香蕉根系分泌物中特有的延胡索酸只能誘導B.subtilis產生趨化反應。該研究表明,植物能夠通過分泌特定的化合物對趨化細菌進行特異性選擇,而這一選擇實現的前提則是細菌的趨化系統決定了其對何種化合物會產生趨化反應。從群落水平的角度來說,植物的根系分泌譜與根際細菌趨化系統的組成之間一定存在特定關系,這正是植物實現對細菌趨化系統特異性選擇的基礎。此外,之前的研究主要是在實驗室條件下利用特定的單一化合物或多種化合物進行細菌趨化反應的誘導,缺少針對根系分泌物與細菌趨化反應間的全面系統性研究,以及更深層次上與環境生態因子的復合影響效果。
植物根系代謝組學技術可以實現對植物根系分泌物中初級、次級代謝產物的定性及定量分析[39],已成為植物代謝研究的常用手段。比如,M?nchgesang等[40]通過根系代謝組學技術對19種不同基因型擬南芥的根系分泌物譜進行了解析,建立了擬南芥基因型與其根系分泌物譜之間的聯系;Zhalnina等[41]通過比較基因組學和代謝組學的聯合分析,發現特定細菌對根系分泌物中芳香類有機酸的代謝偏好性決定了其在燕麥根際的特異性富集。因此,未來可利用植物代謝組學技術深入探索植物對細菌趨化系統特異性選擇的調控機制。
4 趨化細菌類群在根際微生物群落結構調控中的功能
在自然生態系統中,微生物之間通過營養代謝、信號交流、生存空間的競爭與共享等進行競爭或者協作[42]。在根際微生物組群落結構的調控因素中,除宿主植物外,微生物間相互作用也是重要因素之一[43]。細菌趨化系統不僅能夠介導植物-微生物間相互作用,還能夠介導微生物間相互作用。Garbeva和de Boer[44]研究發現,假單胞菌屬細菌Pseudomonassp.和土地桿菌屬細菌Pedobactersp.在碳源限制的條件下共同生長時,表現出競爭性相互作用,且二者的趨化基因表達水平都顯著提高,證明了趨化系統在介導細菌種間競爭中的功能。Lu等[45]對通過種間協作提高產氫效率的梭菌屬細菌Clostridiumcellulovorans和紅假單胞菌屬細菌Rhodopseudomonaspalustris進行了轉錄組分析,發現C.cellulovorans分泌的揮發性脂肪酸可能作為R.palustris的趨化吸引物,導致其趨化基因表達水平上調,說明趨化系統同樣能夠介導細菌的種間協作。此外,Haq等[46]的研究表明,趨化系統還能夠介導伯克霍爾德菌屬細菌Burkholderiaterrae與離褶傘屬Lyophyllum和木霉屬Trichoderma真菌間的相互作用。以上研究主要針對單一菌株間的相互作用,在群落水平上,細菌趨化系統在調控微生物間相互作用進而影響群落結構方面同樣起到重要作用,但是其生態功能仍缺少深入研究。
近年來,生物信息學家開始使用共現性網絡分析來探索微生物間相互作用并預測微生物群落中的關鍵核心類群[47-48]。比如,Agler等[49]研究發現,在擬南芥葉際微生物組共現性網絡中處于關鍵核心位置的特定微生物類群,能夠通過微生物間相互作用傳遞環境和宿主對微生物群落的影響效應,明確了關鍵核心類群的調控功能。Ye等[50]最新的研究表明,一株捕食性粘細菌Corallococcussp. EGB能夠通過趨化反應感應黃瓜的根系分泌物進而向根部遷移,在遷移過程中,捕食病原菌尖孢鐮刀菌Fusariumoxysporumf. sp.cucumerinum;共現性網絡分析的結果表明,這一捕食性粘細菌還能夠調控土壤微生物群落結構。該研究結果揭示了趨化細菌類群在土壤微生物群落結構調控中的生態功能,為后續的研究提供了重要借鑒。
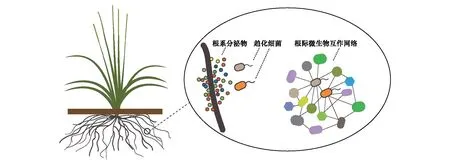
圖2 根際微生物組中的趨化細菌類群Fig.2 A model illustrates the function of chemotactic bacteria in rhizosphere microbiome
5 展望
綜上,研究總結提出根際微生物組中細菌趨化系統的作用機制,細菌趨化系統感應植物根系分泌物,介導趨化細菌與植物間的相互作用;進一步,趨化細菌類群利用趨化系統與其他微生物類群發生相互作用,作為關鍵類群在根際微生物富集中發揮重要的調控作用。因此,細菌趨化系統在根際微生物組募集組裝過程中發揮著關鍵的生態功能,這一功能的解析將是揭示植物對根際微生物組選擇機制的重要突破口之一(圖2)。
針對根際微生物組中細菌趨化系統的生態功能,建議未來研究方向從解析宿主植物對細菌趨化系統的選擇規律入手,明確趨化基因在根際的多樣性及分布規律;然后建立根系分泌物與趨化系統的聯系,闡明宿主植物對趨化系統特異性選擇的調控機制;并進一步探索關鍵趨化細菌類群在根際微生物群落結構中的調控功能。研究方法上,在擴增子高通量測序和宏基因組測序的基礎上,進行代謝組學檢測,并結合菌株分離回接驗證等,將分子生物學、生態學和微生物學等相關知識有效融合,最終揭示細菌趨化系統在植物對根際微生物組選擇中的作用機制,增進對根際微生物組募集組裝過程的認識,為根際微生物組的改良重組提供理論依據。
猜你喜歡
工業設計(2022年8期)2022-09-09 07:43:20
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
軍民兩用技術與產品(2021年10期)2021-03-16 06:05:30
北京測繪(2020年12期)2020-12-29 01:33:58
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
家庭影院技術(2017年9期)2017-09-26 03:41:45
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
