蘭州北山不同海拔3種典型綠化樹種光合特性研究
2022-01-07 09:32:04劉旻霞于瑞新穆若蘭夏素娟
生態(tài)環(huán)境學(xué)報(bào) 2021年10期
關(guān)鍵詞:植物
劉旻霞,于瑞新,穆若蘭,夏素娟
西北師范大學(xué)地理與環(huán)境科學(xué)學(xué)院,甘肅 蘭州 730070
植物作為整個(gè)碳循環(huán)過程的紐帶,對生態(tài)系統(tǒng)碳固定、水平衡以及養(yǎng)分動(dòng)態(tài)等各方面都起著至關(guān)重要的作用,光合作用是植物生長發(fā)育的基礎(chǔ),其對環(huán)境變化較為敏感(王海珍等,2017;劉旻霞等,2020)。海拔梯度由于包含了溫度、濕度、光照等環(huán)境因子的劇烈變化,這些因子的變化極易影響高山植物的葉片形態(tài)和光合作用,因此成為研究植物在生理格局變化的理想?yún)^(qū)域(李鑫豪等,2019)。植物的光合生理特性在一定程度上能夠體現(xiàn)其對生境的響應(yīng)情況,光合-光響應(yīng)曲線反映了植物光合速率與光照強(qiáng)度的相關(guān)關(guān)系,通過曲線可得出植物表觀量子效率(AQE)、光補(bǔ)償點(diǎn)(LCP)、最大凈光合速率(Pnmax)、光飽和點(diǎn)(LSP)和暗呼吸速率(Rd)等重要光合參數(shù)(陳曉英等,2020),這些參數(shù)有助于了解光合作用中植物利用光強(qiáng)的能力及其對生境的適應(yīng)情況,對于研究植物的光合生理特性、生長情況具有重要意義(姬明飛等,2013)。海拔變化作為體現(xiàn)植物生境空間異質(zhì)性的重要方面,隨著海拔升高,植物會(huì)進(jìn)行一系列功能性狀及生理生態(tài)的變化如:植株矮化,葉面積減小,氣孔密度增加,葉片氮含量增加,光合潛力增大等以適應(yīng)高海拔環(huán)境。相關(guān)研究也相繼展開,如韋玉等(2014)應(yīng)用直角雙曲線修正模型擬合分析不同海拔高度矮嵩草(Kobresia humilis)的光合響應(yīng)差異,發(fā)現(xiàn)生長于高海拔地區(qū)的矮嵩草具有較高的 LSP和LCP,表現(xiàn)出對高海拔良好的適應(yīng)性。高成杰等(2015)研究發(fā)現(xiàn)高海拔地區(qū)滇重樓(Pairs polyphylla)花萼對光強(qiáng)的適應(yīng)范圍較大;低海拔地區(qū)滇重樓花萼Pnmax和LSP顯著低于葉片,AQE高于葉片,對光強(qiáng)的適應(yīng)范圍較小,對弱光的利用能力更高。樊瑩(2019)在對長白山不同海拔下的樹種研究發(fā)現(xiàn),白樺(Betula platyphylla)幼樹的Rd隨海拔高度的增加而增加,而花揩槭(Acer ukurunduense)幼樹和紫椴(Tilia amurensis)幼樹的Rd沒有顯著的海拔差異。
蘭州市位于我國西北部,是我國向西開放及一帶一路上的節(jié)點(diǎn)城市。蘭州北山地處黃土高原溝壑區(qū),屬于典型的溫帶大陸性氣候,自然生態(tài)環(huán)境脆弱,面臨著植被破壞、水土流失等一系列嚴(yán)峻的問題。蘭州北山人工林生態(tài)系統(tǒng)對蘭州市城市生態(tài)環(huán)境有著舉足輕重的影響,但由于造林成活率低,植被稀疏,水土流水嚴(yán)重,現(xiàn)已成為黃河的主要沙源區(qū)之一。為更好地恢復(fù)與重建該地區(qū)生態(tài)環(huán)境,關(guān)鍵是要對植樹造林工程中樹種的生理特性有更深刻的認(rèn)識,從而在未來植樹造林中合理選擇綠化樹種、提高植被成活率。側(cè)柏(Platycladus orientalis)、山桃(Prunus davidiana)和甘蒙檉柳(Tamarix austromongolica)具有生長速度快,耐鹽堿、耐貧瘠等特點(diǎn),為干旱半干旱、黃土高原及山坡的主要水土保持林和造林樹種。本研究通過直角雙曲線修正模型對蘭州北山典型綠化樹種側(cè)柏、山桃和甘蒙檉柳的光響應(yīng)曲線進(jìn)行擬合,對3種綠化樹種的Pnmax、AQE、LSP、LCP、Rd及WUE等參數(shù)比較,闡明其對不同海拔的響應(yīng)策略,以期為蘭州北山營建生態(tài)林和綠化建設(shè)提供實(shí)踐指導(dǎo),以選擇出適應(yīng)不同生境的最優(yōu)樹種,為黃土高原半干旱區(qū)植物的栽培、提高植被成活率提供理論基礎(chǔ)和數(shù)據(jù)支持。
1 材料與方法
1.1 研究區(qū)概況
研究區(qū)位于甘肅省蘭州市北山九州臺,坐標(biāo)為 35°58′54″— 36°10′20″N , 103°41′65″—103°52′47″ E,氣候?qū)儆诒睖貛О敫珊禋夂颍艽箨懠撅L(fēng)性氣候的影響,該地區(qū)干燥少雨,年均降雨量211 mm,蒸發(fā)量1446 mm,晝夜溫差大,年均氣溫 5—9 ℃,土壤為黃土母質(zhì)上發(fā)育起來的灰鈣土,有機(jī)質(zhì)含量在 5—15 g·kg?1之間,pH 8—9(劉旻霞等,2020)。區(qū)內(nèi)造林樹種主要有:刺槐(Robinia pseudoacacia)、側(cè)柏、山桃、甘蒙檉柳、油松(Pinus tabulaeformis)、檸條(Caragana korshinkii)、紅砂(Reaumurta soongorica)和新疆楊(Populus alba)等。
1.2 研究方法
1.2.1 樣地選擇及樣品確定
本研究于2018年7月上旬,應(yīng)用GPS準(zhǔn)確定位并選取海拔為 1600、1800、2000 m的 3個(gè)樣區(qū)(表 1),每個(gè)樣區(qū)內(nèi)分別設(shè)置 3個(gè) 30 m×30 m的樣地,每個(gè)樣地間相隔距離50 m,樣地內(nèi)植被均為人工林,樹齡大約15年左右,每個(gè)樣地選取生長狀況相似,無病蟲害的側(cè)柏、山桃和甘蒙檉柳各5株,每株分別選取東、西、南、北 4個(gè)方位中上層受光方向一致,葉位相同、長勢相近、生長健康的 3片葉子進(jìn)行光響應(yīng)曲線的測定。測量中葉片面積小于葉室面積的葉片,將其裝入自封袋后帶回實(shí)驗(yàn)室,用掃描儀(CanoScan LiDE 120,Japan)掃描,利用Image J軟件進(jìn)行計(jì)算。
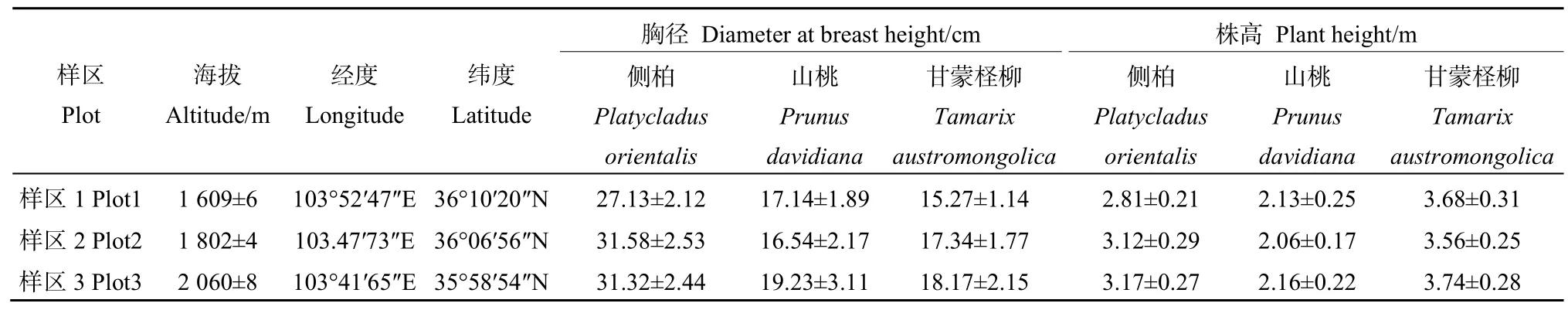
表1 樣區(qū)植被概況Table 1 Survey of Vegetation in Sample plot
1.2.2 光合速率-光響應(yīng)曲線的測定
采用Li-6400XT(Li-COR6400Inc. USA)便攜式光合測定儀對植物光合-光響應(yīng)曲線進(jìn)行測定。選擇晴朗無風(fēng)的天氣于09:00—11:30之間測定 3個(gè)樹種的凈光合速率(Net photosynthetic rate,Pn)、胞間 CO2濃度(Intercellular CO2concentration,Ci)、氣孔導(dǎo)度(Stomatal conductance,Gs)等光合指標(biāo)。測定前對每個(gè)葉片進(jìn)行充分的光誘導(dǎo),設(shè)置光照強(qiáng)度梯度為:2000、1800、1500、1200、1000、800、500、200、150、100、50、0 μmol·m?2·s?1。測定時(shí)在每個(gè)設(shè)置的光合有效輻射下適應(yīng)3 min。設(shè)置流速為500 μmol·s?1,安裝配套儀器 CO2鋼瓶,設(shè)定葉室CO2摩爾分?jǐn)?shù)為 (400±5) μmol·mol?1,葉溫為(25±1) ℃,相對濕度變化范圍為55%左右。用直角雙曲線修正擬合光響應(yīng)曲線,得到一系列光合參數(shù),進(jìn)行分析后,計(jì)算水分利用效率 (water use efficiency, WUE)=Pn/Tr,氣孔限制值Ls=1?Ci/Ca。
1.2.3 環(huán)境因子的測定
使用儀器 JZ-RHXL3SD溫濕度記錄儀,DP-GH光合有效輻射記錄儀、測量樣區(qū)內(nèi)大氣溫度(Atmospheric temperature,Ta)、空氣相對濕度(Relative air humidity,RH)、光合有效輻射(Photosynthetically active radiation,PAR)等環(huán)境因子。選前3天內(nèi)沒有發(fā)生降雨的天氣進(jìn)行土壤采集,在每個(gè)樣地內(nèi)按照對角線取樣法用直徑為5 cm的土鉆取大約1000 g左右的土壤樣品(土層厚度為0—20 cm),混合均勻后裝入自封袋,通過烘干法(Gardener,1986)測定土壤含水率(Soil moisture content,SMC)。
1.2.4 光響應(yīng)曲線擬合模型
采用直角雙曲線修正模型對光響應(yīng)曲線進(jìn)行擬合。

式中:
Pn——凈光合速率;
I——光合有效輻射;
α——光響應(yīng)曲線的初始斜率;
Pnmax——最大凈光合速率;
Rd——暗呼吸速率。
γ——光飽和項(xiàng),其值為α/Pnmax。
1.2.5 資源利用效率的計(jì)算
水分利用效率公式(Water use efficiency,WUE,μmol·mmol?1),根據(jù) Medrano et al.(2015)提出的公式計(jì)算:

光能利用效率(Light use efficiency,LUE,mol·mol?1)公式為:

式中:
PPFD——光合有效輻射(μmol·m?2·s?1)。
1.3 數(shù)據(jù)分析
本研究通過Microsoft Excel 2010對數(shù)據(jù)進(jìn)行分析處理,用SPSS 17.0軟件擬合光響應(yīng)曲線及其光合參數(shù)以及對數(shù)據(jù)進(jìn)行顯著性分析,用 Origing 17.0軟件進(jìn)行繪圖。
2 結(jié)果與分析
2.1 不同海拔環(huán)境因子的變化
樣區(qū)內(nèi)環(huán)境因子日均值如表 2,隨著海拔由1600 m升高至2000 m過程中,PAR有顯著差異(P<0.05),呈不斷增強(qiáng)的趨勢。Ta、RH、Ts和 SMC均隨海拔的上升而下降,除Ts外,3種環(huán)境因子均有顯著差異。其中Ta由23 ℃降低至19 ℃,RH降低幅度較大,由51%降低至43%、Ts降低了約0.5 ℃,SMC降低了約21%。
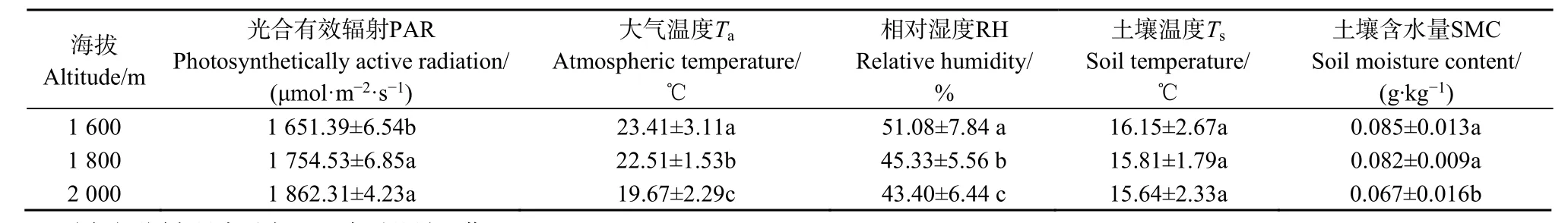
表2 樣區(qū)內(nèi)環(huán)境因子Table 2 Environmental factors in the sample area
2.2 不同海拔3種綠化樹種Pn的光響應(yīng)變化
通過直角雙曲線修正模型對不同海拔下3樹種光合作用的光響應(yīng)曲線進(jìn)行擬合,相關(guān)性均達(dá)到顯著水平(P<0.001)。圖1可以看出,3樹種Pn均隨PAR的增加而升高,而后逐漸趨于平穩(wěn)。低光輻射下不同的海拔梯度Pn的光響應(yīng)變化趨勢一致,隨著光合輻射的增強(qiáng),高海拔梯度Pn的光響應(yīng)變化高于低海拔處的光響應(yīng)變化,且3樹種均呈現(xiàn)出高海拔具有較高的Pn,表明海拔對側(cè)柏、山桃和甘蒙檉柳光合速率影響較大。

圖1 不同海拔Pn對光的響應(yīng)Fig. 1 Response of Pn to light at different altitudes
3樹種光響應(yīng)參數(shù)在不同海拔呈顯著差異(P<0.05),側(cè)柏、山桃和甘蒙檉柳的Pnmax隨海拔的升高均呈增大趨勢;山桃和甘蒙檉柳的LSP隨海拔增加而上升,在海拔2000 m處達(dá)到最大,而側(cè)柏LSP在1600 m處達(dá)到最大;3樹種的LCP和 Rd隨海拔升高均呈現(xiàn)降低趨勢;側(cè)柏、山桃和甘蒙檉柳的AQE均表現(xiàn)為海拔2000 m最高,1600 m處最低(表3)。
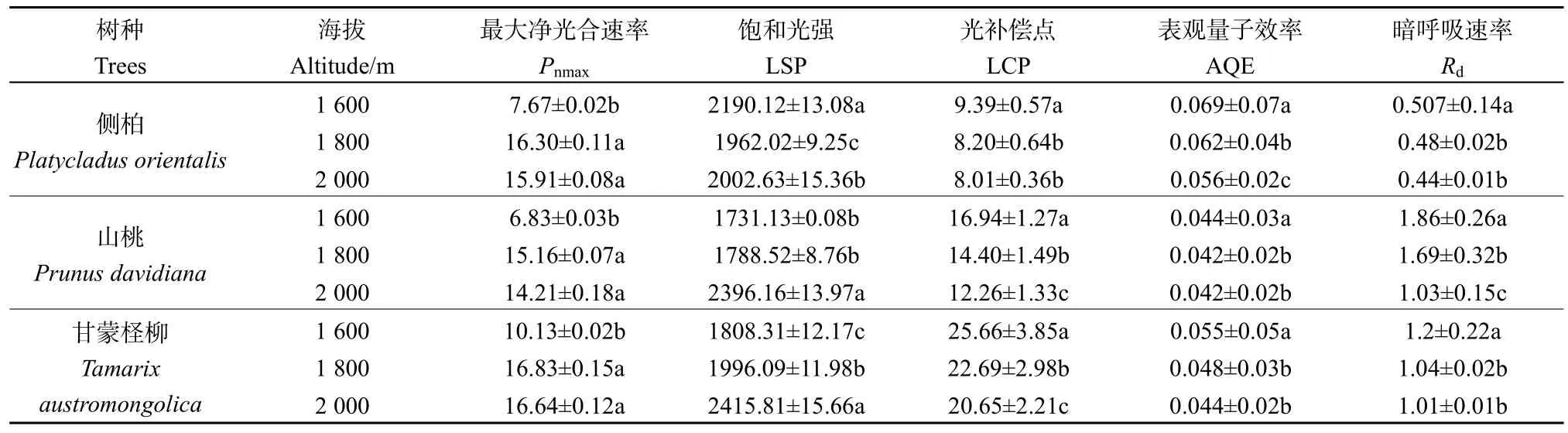
表3 不同海拔側(cè)柏、山桃和甘蒙檉柳的光合參數(shù)變化Table 3 Photosynthetic parameters of of P. orientalis, P. davidiana and T. austromongolica at different altitudes
2.3 不同海拔3種樹種的光合生理指標(biāo)
圖2表明,不同海拔下,側(cè)柏、山桃和甘蒙檉柳葉片Tr和Gs均呈現(xiàn)出隨PAR增加而上升的趨勢,分別在 PAR 大于 1273、1562、1816 μmol·m?2·s?1時(shí),Tr和Gs趨于穩(wěn)定。當(dāng)側(cè)柏、山桃和甘蒙檉柳的 PAR 值分別由 0 μmol·m?2·s?1增大至光飽和點(diǎn)過程中,3樹種的Ci呈現(xiàn)出下降趨勢,而Ls呈現(xiàn)出上升趨勢。在 PAR 值為 400 μmol·m?2·s?1之前,Ci下降幅度極大,超過 400 μmol·m?2·s?1后,Ci下降緩慢。在光飽和點(diǎn)處,Ci達(dá)到最小值。1600 m和1800 m處,3樹種Ci達(dá)到最低點(diǎn)后,有緩慢上升的趨勢,此時(shí)表現(xiàn)出明顯的光抑制現(xiàn)象。在海拔2000 m處,植物擁有較大的Tr、Gs和Ci以抵抗高溫等不良環(huán)境。3樹種平均Tr、Gs和Ci由大到小為:山桃>甘蒙檉柳>側(cè)柏。

圖2 不同光照強(qiáng)度下側(cè)柏、山桃和甘蒙檉柳的光合生理指標(biāo)Fig. 2 Photosynthetic physiological indexes of P. orientalis, P. davidiana and T. austromongolica under different light intensity
2.4 3種樹種光能、水分利用效率比較
LUE是反映植物生長發(fā)育的重要生理指標(biāo)。由圖3可以看出,側(cè)柏、山桃和甘蒙檉柳在低光強(qiáng)(PAR<100 μmol·m?2·s?1)條件下,隨著 PAR的升高,LUE快速上升并達(dá)到峰值;此后 LUE逐漸下降。不同海拔梯度上3個(gè)樹種的LUE均表現(xiàn)出2000 m>1800 m>1600 m。其中,山桃LUE最大,甘蒙檉柳次之,側(cè)柏最小。3樹種的WUE分別在 PAR 小于 525、870、450 μmol·m?2·s?1光強(qiáng)下,WUE顯著增加,隨著PAR的增加,WUE逐漸趨于平緩。表明側(cè)柏、山桃和甘蒙檉柳WUE對 PAR 的適應(yīng)范圍分別在 525—2000 μmol·m?2·s?1、870—2000 μmol·m?2·s?1和 450—2000 μmol·m?2·s?1之間,可以看出甘蒙檉柳對PAR的適應(yīng)范圍更大。側(cè)柏和甘蒙檉柳呈現(xiàn)出在2000 m處具有較低的WUE,而在1600 m處具有較高的WUE。山桃整體表現(xiàn)出在海拔1800 m處WUE較高(圖3)。

圖3 不同光照強(qiáng)度下側(cè)柏、山桃和甘蒙檉柳的資源利用效率Fig. 3 Resource utilization efficiency of P. orientalis, P. davidiana and T. austromongolica under different light intensity
2.5 側(cè)柏、山桃和甘蒙檉柳生理生態(tài)因子的相關(guān)性分析
側(cè)柏、山桃和甘蒙檉柳生理生態(tài)因子顯示出不同的差異性。生態(tài)因子中,PAR、Ta、Ca和RH之間存在緊密聯(lián)系。其中,側(cè)柏和山桃的 Pn與PAR呈現(xiàn)顯著正相關(guān),甘蒙檉柳Pn與Ts呈極顯著正相關(guān)關(guān)系。生理因子中,側(cè)柏、山桃和甘蒙檉柳Pn均與LUE、WUE呈現(xiàn)顯著正相關(guān)關(guān)系,且相關(guān)系數(shù)均在0.6以上(表4)。
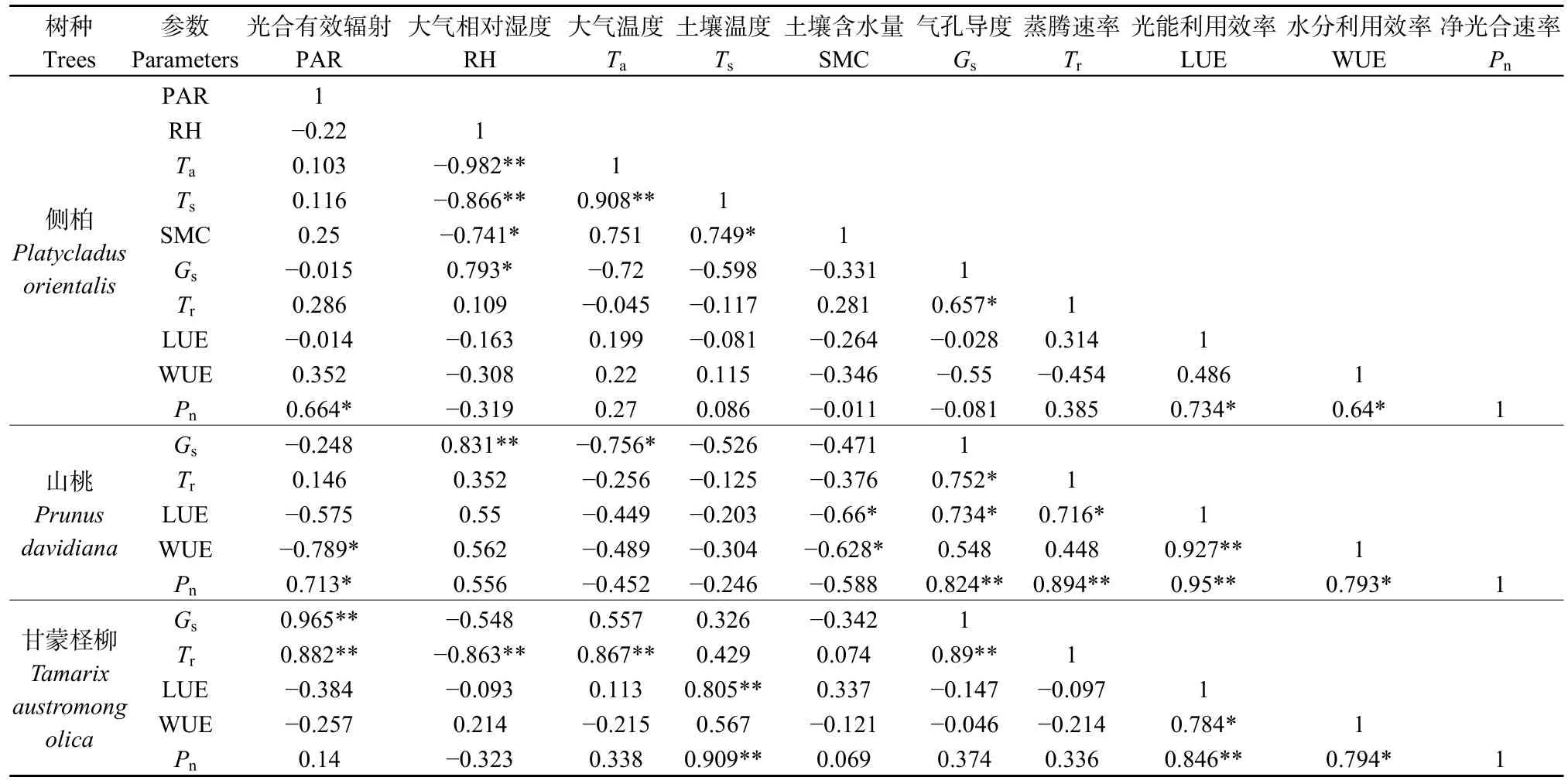
表4 側(cè)柏、山桃及甘蒙檉柳生理生態(tài)因子間的相關(guān)性分析Table 4 Correlation analysis of physiological and ecological factors of P. orientalis, P. davidiana and T. austromongolica
2.6 3種樹種生理生態(tài)因子的線性回歸分析
方程 (a) 可以看出,側(cè)柏Pn隨著 LUE、PAR的升高表現(xiàn)出增大的趨勢。方程 (b) 可知,LUE和Tr影響山桃的Pn。方程 (c) 表明,甘蒙檉柳隨Tr的增大表現(xiàn)出升高趨勢。
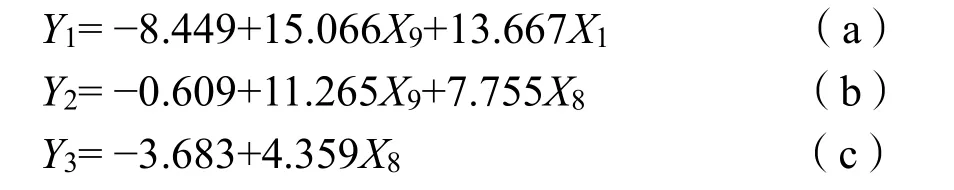
式中:
Y1、Y2、Y3——側(cè)柏、山桃和甘蒙檉柳的Pn;
X1——光合有效輻射PAR;
X2——相對濕度RH;
X3——空氣溫度Ta;
X4——土溫 Ts;
X5——土壤含水率SMC;
X6——?dú)饪讓?dǎo)度Gs;
X7——蒸騰速率Tr;
X8——水分利用效率WUE;
X9——光能利用效率LUE。
3 討論
3.1 不同海拔下 3種樹種的光響應(yīng)曲線及參數(shù)分析
光合-光響應(yīng)曲線是理解植物光化學(xué)效率的重要方式,曲線的參數(shù)能夠反映植物光合作用機(jī)制對環(huán)境的響應(yīng)。Pnmax代表植物葉片的最大光合能力,一定程度上反映了植物的遺傳特性和對環(huán)境的適應(yīng)能力,其值越大,表明其光合作用潛力越大。側(cè)柏、山桃和甘蒙檉柳的 Pnmax隨海拔的升高均呈增大趨勢,反映了植物在相對高海拔梯度上具有最大的光合能力,是植物對環(huán)境長期適應(yīng)的結(jié)果(范秀華等,2012)。其中甘蒙檉柳的最大光合能力優(yōu)于側(cè)柏和山桃。LSP能夠反映植物對強(qiáng)光的利用能力,本研究中山桃和甘蒙檉柳的 LSP隨海拔增加呈現(xiàn)上升趨勢,表明山桃和甘蒙檉柳對強(qiáng)光照的適應(yīng)能力較好。原因是隨海拔的升高,植物體內(nèi)類胡蘿卜素含量顯著增加(Liu et al.,2020)。類胡蘿卜素不僅可以吸收和傳遞光能,而且能夠通過一些熱耗散機(jī)制如葉黃素循環(huán)等來消耗多余的光能以避免光合機(jī)構(gòu)的損傷(孫小玲等,2010)。側(cè)柏的LSP呈一定的降低趨勢(表 3),表明側(cè)柏對強(qiáng)光的耐受能力相對較弱。通常植物具有較高的 AQE和較低的LCP,表明其對弱光的利用能力較強(qiáng)(Joseph et al.,2005;Wang et al.,2021),即具有較強(qiáng)的耐蔭性。3樹種不同海拔下的LCP均隨海拔升高呈現(xiàn)降低趨勢,LCP較低及LSP較高的植物對光環(huán)境的適應(yīng)性強(qiáng)(郭春燕等,2013),說明海拔升高,樹種對干旱環(huán)境適應(yīng)能力增強(qiáng)。本研究中,3樹種的AQE均隨海拔的升高而升高,這是由于光合作用中的關(guān)鍵酶Rubisco是一個(gè)調(diào)節(jié)光合作用和光呼吸的雙功能酶(Quimado et al.,2021)。有研究表明,降低空氣中的O2濃度或提高CO2濃度會(huì)促進(jìn)植物Rubisco酶的活性和數(shù)量,從而使AQE升高(潘璐等,2014)。植物葉片Rd是光強(qiáng)為0時(shí)的光合速率,較低的Rd可以降低呼吸作用對光合產(chǎn)物的消耗,有利于干物質(zhì)的積累(Chen et al.,2011)。側(cè)柏、山桃和甘蒙檉柳的 Rd在海拔梯度上呈現(xiàn)相同的變化規(guī)律,均隨海拔的升高而降低,其原因可能是溫度限制了植物的生理活性,從而減少對于能量的需求,使Rd下降。
3.2 海拔梯度上3種樹種氣體交換參數(shù)分析
在PAR逐漸增強(qiáng)的過程中,3樹種的Tr和Gs顯著增加,其原因是植物在進(jìn)行光合作用過程中對水分及礦物質(zhì)營養(yǎng)元素的需求會(huì)增加,因此需要蒸騰作用將營養(yǎng)物質(zhì)傳輸?shù)街参锼璧母鱾€(gè)部位(劉旻霞等,2020)。而Ci的光響應(yīng)則相反,隨著PAR的增強(qiáng),Pn快速增加,消耗 CO2量較大,導(dǎo)致 Ci顯著下降,這時(shí)植物通過調(diào)節(jié)氣孔來限制光合作用,達(dá)到最大CO2同化量與最小水分散失量(荊天,2015)。當(dāng)PAR超過光飽和點(diǎn)LSP,Ci趨于穩(wěn)定且略呈上升趨勢,Ls呈略微下降趨勢,光合作用的主導(dǎo)限制因素由氣孔限制轉(zhuǎn)變?yōu)榉菤饪紫拗疲≧en et al.,2020),植物逐漸出現(xiàn)光抑制現(xiàn)象。側(cè)柏、山桃和甘蒙檉柳葉片 Tr和Gs都呈現(xiàn)出海拔升高而上升的趨勢,這是由于高海拔地區(qū)的低氣壓環(huán)境增加了氣體分子擴(kuò)散能力,以及通過增加葉片氣孔密度導(dǎo)致Gs增加,植物通過調(diào)節(jié)氣孔的大小控制植物吸收 CO2和散失水分,所以 Tr、Ci與 Gs變化趨勢一致,也隨著海拔的增加而增加。
3.3 3種樹種的光能、水分利用效率比較
LUE是表征植物固定太陽能效率的指標(biāo),指植物通過光合作用將所截獲/吸收的能量轉(zhuǎn)化為有機(jī)干物質(zhì)的效率,是植物光合作用的重要概念(Wang et al.,2021)。本研究中隨著海拔升高3樹種光合潛力增大,其原因是高海拔植物光合機(jī)構(gòu)通過增加LUE來減少過剩光能對光合器官的光抑制和光破壞,以適應(yīng)多變的生存環(huán)境。3樹種LUE表現(xiàn)為:山桃最大,甘蒙檉柳次之,側(cè)柏最小,表明山桃可以更有效地利用光能,從而轉(zhuǎn)化為干物質(zhì)。植物WUE是植物生長發(fā)育所需最佳水分供應(yīng)的重要指標(biāo)之一,3樹種WUE在PAR的較大范圍內(nèi)都能夠維持較高的水平,其中甘蒙檉柳表現(xiàn)出對PAR的適應(yīng)范圍更大,表明甘蒙檉柳對光的適應(yīng)生態(tài)幅度較寬。從海拔上來看,側(cè)柏和甘蒙檉柳表現(xiàn)為 WUE隨著海拔的升高而降低,而山桃整體表現(xiàn)出在海拔1800 m處WUE較高。原因可能是高海拔地區(qū)植物受到更多的干旱、光合輻射強(qiáng)、低溫等環(huán)境脅迫,側(cè)柏和甘蒙檉柳提高WUE以抵御環(huán)境脅迫。
3.4 3種綠化樹種生理生態(tài)因子之間的關(guān)系
植物光合作用過程受生理因子和生態(tài)因子的共同影響,在相同的環(huán)境條件下,不同植物表現(xiàn)出的光合作用動(dòng)態(tài)變化的原因不同(徐春華等,2015)。本研究中影響3樹種光合作用的各因子存在著明顯的差異。側(cè)柏和山桃的PAR、Tr對Pn的影響較大,原因是PAR是植物光合作用的動(dòng)力,一方面可以影響葉面溫度,促進(jìn)水分的蒸發(fā),另一方面,影響氣孔的開放,從而影響植物的氣體交換,加速植物的光合速率;WUE與PAR、Ta、Ts、Pn呈正相關(guān),與RH、SMC、Tr、Gs呈負(fù)相關(guān),表明隨PAR和Ta的升高,植物的Pn增強(qiáng),使得WUE不斷增大,隨著植物Tr增大,植物通過氣孔散失水分,導(dǎo)致WUE減小。甘蒙檉柳的Ts對Pn的影響較大,有研究表明(范愛武等,2004),土溫對根系生長、代謝具有很大影響,過高或過低的土溫會(huì)抑制葉片中淀粉的降解和轉(zhuǎn)運(yùn)以及糖和蛋白質(zhì)之間的轉(zhuǎn)化,從而影響植物葉片的光合作用及生長。本研究中,山桃和甘蒙檉柳的WUE與Pn的相關(guān)性較強(qiáng),表明植物的Pn越高,樹種消耗相同的水分所產(chǎn)生的干物質(zhì)就越多。
綜上所述,本研究通過對蘭州北山3種綠化樹種的光響應(yīng)曲線及其參數(shù)進(jìn)行研究,發(fā)現(xiàn)在蘭州南北兩山多年的生態(tài)恢復(fù)過程中,側(cè)柏耐陰性較強(qiáng)且對光強(qiáng)的適應(yīng)范圍不廣,適合栽培在低海拔地區(qū);山桃可以很好利用光能,但其抗旱性能較差;甘蒙檉柳在較大的PAR范圍內(nèi)可以保持良好的 WUE,并且其對光能的適應(yīng)范圍較廣,可以大面積栽培。該研究結(jié)果可為今后該地區(qū)乃至西北干旱區(qū)半干旱區(qū)人工造林樹種的優(yōu)選及生態(tài)環(huán)境的恢復(fù)提供參考依據(jù)。
4 結(jié)論
(1)海拔對側(cè)柏和山桃的Pn影響較大;而對甘蒙檉柳的影響較小。
(2)隨海拔梯度的升高,3種樹種在海拔2000 m處具有較高的Pnmax和AQE,而在海拔1600 m處具有較高的 LCP及 Rd。除側(cè)柏外,山桃和甘蒙檉柳的LSP隨海拔增加呈現(xiàn)上升趨勢。
(3)高海拔 3種樹種的光合機(jī)構(gòu)通過增加LUE來減少過剩光能對光合器官的光抑制和光破壞,從而適應(yīng)多變的生存環(huán)境;高海拔生境植物受到更多的干旱、光輻射、低溫等環(huán)境脅迫,側(cè)柏和甘蒙檉柳提高WUE以抵御環(huán)境脅迫。
(4)PAR、Tr對側(cè)柏和山桃Pn的影響較大;Ts對甘蒙檉柳Pn的影響較大。
猜你喜歡
少兒科學(xué)周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學(xué)閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數(shù)英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數(shù)英綜合(2020年11期)2020-12-16 02:57:22
學(xué)苑創(chuàng)造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
