EGCG通過激活AMPK、抑制PPARγ調(diào)控3T3-L1脂肪細胞中脂滴蓄積
2022-01-06 02:31:40管巧麗吳曉云
食品科學 2021年23期
關(guān)鍵詞:質(zhì)量
管巧麗,吳曉云,李 芳,*,盛 軍
(1.云南農(nóng)業(yè)大學 普洱茶學教育部重點實驗室,云南 昆明 650201;2.云南農(nóng)業(yè)大學食品科學技術(shù)學院,云南 昆明 650201;3.云南農(nóng)業(yè)大學理學院,云南 昆明 650201)
肥胖是近年來世界范圍內(nèi)的主要健康問題之一,通常是由于食物攝入與能量消耗不平衡,脂肪細胞肥大和增生而導致的脂肪量過多和脂肪組織擴張[1]。早在2013年肥胖被美國醫(yī)學協(xié)會(American Medical Association)認定為一種疾病之前,肥胖的發(fā)生率一直在穩(wěn)步增長[2]。截至2016年的全球統(tǒng)計數(shù)據(jù)顯示,13%(6.5億)的成年人肥胖,19億人超重,據(jù)估計,2025年全球60%的死亡將由肥胖相關(guān)疾病引起[3]。在遺傳易感性個體中,高熱量飲食和缺乏體力活動等環(huán)境因素可能會引發(fā)肥胖。作為一種慢性疾病,抗肥胖治療依賴于生活方式的改變、手術(shù)和藥物治療[4-6]。相對于使用手術(shù)和藥物減輕體質(zhì)量,運動或者通過飲食干預更容易被人們接受。
茶是一種被人們廣泛接受的飲料。綠茶已被證明具有許多潛在的健康益處[7],包括抗氧化、抗癌和保護心臟活性,并可能影響能量平衡的多個方面[8-9],綠茶對健康的好處主要是由于其高濃度的多酚,這些多酚統(tǒng)稱為兒茶素。綠茶多酚被廣泛報道可以改善人類和動物模型的肥胖和代謝綜合征[10-11]。有研究表明綠茶提取物和表沒食子兒茶素沒食子酸酯(epigallocatechin gallate,EGCG)可顯著提高脂質(zhì)活性并降低脂肪組織質(zhì)量[12-13]。在探究綠茶多酚對減輕體質(zhì)量作用的研究中,報道了幾種多酚可以刺激腺苷一磷酸(adenosine monophosphate,AMP)活化蛋白激酶(AMP-activated protein kinase,AMPK)的活性[14-16]。激活AMPK會降低糖異生和脂肪酸合成,增加分解代謝[17-19],有研究者提出“AMPK假說”,AMPK在介導EGCG對脂肪酸合成和分解代謝的作用中起重要作用[6]。他們提出了EGCG促進減輕體質(zhì)量的兩個主要機制:1)減少腸道內(nèi)脂質(zhì)和蛋白質(zhì)的吸收,從而減少熱量攝入;2)AMPK在肝臟、骨骼肌和白色脂肪組織中的激活。激活AMPK途徑可能為肥胖、胰島素抵抗、內(nèi)分泌和心血管疾病的治療提供新的靶點[20]。Moreno等在EGCG影響高脂飲食誘導肥胖模型的研究中發(fā)現(xiàn),EGCG可以降低小鼠肥胖和附睪白色脂肪組織質(zhì)量,并且部分是通過激活AMPK實現(xiàn)的[21]。也有文獻指出EGCG可以抑制脂肪細胞的分化[6],但EGCG減輕體質(zhì)量和脂肪組織質(zhì)量的具體機制不清楚,也鮮有文獻報道EGCG改善細胞中脂滴蓄積的具體機制。因此,本研究以EGCG為實驗原料,通過體外誘導分化培養(yǎng)3T3-L1前脂肪細胞,研究EGCG對3T3-L1脂肪細胞中脂滴蓄積的影響,為改善肥胖癥的分子機制提供理論參考。
1 材料與方法
1.1 材料與試劑
3T3-L1前脂肪細胞 中國科學院昆明動物研究所;EGCG(純度大于98%) 成都生物凈化植物化工有限公司;Dublecco改良Eagle培養(yǎng)基(Dulbecco’s modified Eagle medium,DMEM)和高糖培養(yǎng)基 美國HyClone公司;二喹啉甲酸(bicinchoninic acid,BCA)試劑盒上海碧云天生物技術(shù)有限公司;二甲基亞砜(dimethyl sulfoxide,DMSO) 美國Amresco公司;質(zhì)量分數(shù)30%聚丙烯酰胺貯液 中國Bio-RAD公司;0.5 mol/L Tris-HCl 緩沖液(pH 6.8)、1.5 mol/L Tris-HCl緩沖液(pH 6.8)、四甲基乙二胺、3-異丁基-1-甲基黃嘌呤(3-isobutyl-1-methyl-7H-xanthine,IBMX) 美國Sigma公司;噻唑藍(thiazolyl blue tetrazolium bromide,MTT) 博美生物有限公司;抗兔免疫球蛋白、抗鼠免疫球蛋白 美國R&D公司;無水乙醇、油紅O 天津市風船化學試劑科技有限公司;異丙醇 四川西隴化工有限公司;TransZol Up 北京全式金生物技術(shù)有限公司;三氯甲烷(氯仿) 重慶川東化工集團有限公司;質(zhì)量分數(shù)4%多聚甲醛濟南百博生物技術(shù)有限公司;馬來酸羅格列酮中國蓋德化工廠;胰島素 江蘇萬邦生物醫(yī)藥股份有限公司;PrimeScript RT reagent kit with gDNA試劑盒日本Takara公司;去污劑、裂解液、三羥甲基氨基甲烷、十二烷基硫酸鈉、硫酸銨、甘氨酸、甘油、β-巰基乙醇、溴酚藍、考馬斯亮藍G250、牛血清白蛋白、青鏈霉素混合液、10×磷酸鹽緩沖液(phosphate-buffered saline,PBS,pH 7.4) 北京索萊寶生物科技有限公司;脫脂奶粉 美國BD公司。
1.2 儀器與設(shè)備
C150型CO2氣體培養(yǎng)箱 德國Binder公司;DM-1L LED倒置顯微鏡 德國Leica公司;CKX41倒置熒光顯微鏡 日本Olympus公司;U-2910紫外分光光度計日本日立公司;7900HT型熒光定量聚合酶鏈式反應(polymerase chain reaction,PCR)儀 美國應用生物系統(tǒng)公司;Z36HK型高速臺式冷凍離心機 德國Hermle Labortechnik GmbH公司;VORTES QL-902型漩渦儀 海門市其林貝爾儀器制造有限公司;掌上離心機日本TOMY公司;DHG-9240A電熱鼓風干燥箱 上海一恒科學儀器有限公司;FST-III-80純水機 普力菲爾純凈水有限公司;96 孔板 美國Thermo Fisher Scientific公司。
1.3 方法
1.3.1 3T3-L1前脂肪細胞的分化及EGCG處理細胞
細胞復蘇:取凍存的3T3-L1前脂肪細胞,將其置于37 ℃流水下快速融化,融化后加入含體積分數(shù)10%胎牛血清、體積分數(shù)1%青鏈霉素的DMEM高糖培養(yǎng)基10 mL,混勻后離心(1500 r/min、3 min),吸棄離心后的上清液,加入10 mL含體積分數(shù)10%胎牛血清、體積分數(shù)1%青鏈霉素的DMEM高糖培養(yǎng)基吹打細胞并混勻,將含細胞的培養(yǎng)基轉(zhuǎn)移至培養(yǎng)皿中,在37 ℃、5% CO2培養(yǎng)箱中培養(yǎng)。待細胞長滿至80%~90%,融合4~5 d觀察細胞狀態(tài)穩(wěn)定后,吸除培養(yǎng)皿中的培養(yǎng)液,加入37 ℃預熱的PBS清洗細胞,吸除PBS,加入誘導培養(yǎng)基(含10%胎牛血清、1%青鏈霉素、10 μg/mL胰島素、0.5 μmol/L 3-異丁基-1-甲基黃嘌呤、1 μmol/L羅格列酮、1 μmol/L地塞米松的DMEM高糖培養(yǎng)基)。培養(yǎng)3 d后再換胰島素誘導培養(yǎng)基(含10%胎牛血清、1%雙抗、10 μg/mL胰島素的DMEM高糖培養(yǎng)基)培養(yǎng)3 d,最后換含有10%胎牛血清、1%青鏈霉素的高糖DMEM培養(yǎng)基培養(yǎng)2 d,誘導分化10 d左右,顯微鏡下觀察3T3-L1細胞90%表現(xiàn)為成熟脂肪細胞表型(油紅O染色鑒定),即可用于實驗。EGCG處理細胞時使用pH 6.5的DMEM酸性培養(yǎng)基,作用質(zhì)量濃度分別為25、50 μg/mL,以不含EGCG的培養(yǎng)基為對照組(即0 μg/mL的EGCG,后同),孵育時間分別為72 h和144 h。
1.3.2 細胞存活率測定
將處于對數(shù)生長期的3T3-L1細胞接種于96 孔板(2×104個/孔),加入含有體積分數(shù)10%胎牛血清、1%青鏈霉素的DMEM培養(yǎng)基(200 μL/孔),37 ℃孵育24 h后吸棄培養(yǎng)液,更換為含有不同質(zhì)量濃度EGCG(0、25、50 μg/mL)的DMEM培養(yǎng)基(200 μL/孔),每組設(shè)置3個平行,在37 ℃、5% CO2培養(yǎng)箱中培養(yǎng)72 h后,每孔加20 μL 5 mg/mL MTT儲備液,避光,繼續(xù)孵育培養(yǎng)4 h,吸去孔內(nèi)培養(yǎng)上清液,每孔加入150 μL DMSO,搖床上振蕩10 min,使結(jié)晶物充分溶解。在酶標儀上設(shè)置492 nm波長,檢測各孔光密度值并按下式計算細胞存活率。
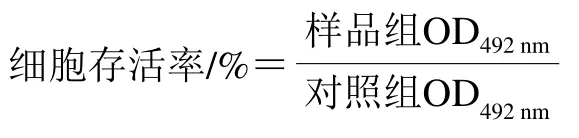
1.3.3 3T3-L1脂肪細胞的鑒定(油紅O染色鑒定)和甘油三酯相對含量的測定
取1.3.1節(jié)不同質(zhì)量濃度EGCG(0、25、50 μg/mL)處理144 h后的3T3-L1細胞培養(yǎng)板,PBS清洗兩次;4%多聚甲醛(4 ℃冰箱存放)固定30 min后,使用PBS清洗兩次;使用異丙醇配制0.5%油紅O儲液,再按油紅O儲液與蒸餾水體積比3∶2配制油紅O工作液,混勻靜置10 min;取500 μL油紅O工作液覆蓋培養(yǎng)板,染色10 min左右;PBS清洗一次,棄上清液;在倒置顯微鏡下調(diào)整物鏡、目鏡,在相同倍數(shù)下觀察并拍照保存圖片。
甘油三酯相對含量的測定:取1.3.1節(jié)不同質(zhì)量濃度EGCG(0、25、50 μg/mL)處理72、144 h后的3T3-L1細胞,參考甘油三酯試劑盒說明書測定甘油三酯的相對含量。
1.3.4 AMPK抑制劑Dorsomorphin處理3T3-L1脂肪細胞的方法
按1.3.1節(jié)方法誘導3T3-L1脂肪細胞以及EGCG(0、50 μg/mL)處理,孵育3T3-L1細胞144 h:加入含有EGCG(50 μg/mL)和AMPK抑制劑Dorsomorphin(40 μmol/L)的DMEM培養(yǎng)基共同孵育3T3-L1細胞144 h,即Dorsomorphin抑制組;只含40 μmol/L Dorsomorphin的DMEM培養(yǎng)基孵育3T3-L1細胞144 h,即單獨抑制劑組。
1.3.5 蛋白質(zhì)免疫印跡法測定蛋白表達水平
取1.3.1節(jié)不同質(zhì)量濃度EGCG(0、25、50 μg/mL)處理72、144 h后的3T3-L1細胞以及1.3.4節(jié)中Dorsomorphin抑制組和單獨抑制劑組細胞,使用含體積分數(shù)1%苯甲基磺酰氟的裂解液提取總蛋白,采用BCA定量法測定蛋白質(zhì)量濃度。在每孔加入蛋白溶液(含60 μg蛋白),用質(zhì)量分數(shù)8%的十二烷基硫酸鈉-聚丙烯酰胺凝膠進行電泳,然后轉(zhuǎn)移至聚偏二氟乙烯膜,將膜用質(zhì)量分數(shù)5%脫脂奶粉試劑封閉1 h,在4 ℃下與一抗Bax、cleaved-Caspase 3、β-actin、磷酸化-AMPK(phospho-AMPK,p-AMPK)、AMPK、過氧化物酶體增殖物激活受體(peroxisome proliferators-activated receptors,PPARγ)、Tubulin、PGC-1α(1∶1 000)孵育12 h以上,然后與4 μL相應的二抗(抗兔免疫球蛋白和抗鼠免疫球蛋白,1∶5 000)室溫搖床孵育1 h。采用Western blotting圖像用超靈敏增強型化學發(fā)光底物試劑盒成像,通過FluorChem E系統(tǒng)獲得結(jié)果圖像,使用AlphaView軟件對相應的條帶進行量化。
1.3.6 實時定量聚合酶鏈式反應測定基因表達水平
取1.3.1節(jié)不同質(zhì)量濃度EGCG(0、25、50 μg/mL)處理72、144 h后的3T3-L1細胞,使用TRIzol提取總RNA,采用TaKaRa PrimeScript?RT reagent Kit With gDNA Eraser試劑盒進行反轉(zhuǎn)錄,采用SYBR Green I reaction system進行PCR,基因表達水平采用2-ΔΔCt法計算。引物序列見表1。

表1 實時定量PCR基因引物序列Table 1 Sequences of primers for real time quantitative polymerase chain reaction used in this study
1.4 數(shù)據(jù)處理與分析
實驗數(shù)據(jù)采用SPSS 17.0軟件和GraphPad Prism 6軟件進行統(tǒng)計學分析,結(jié)果以平均值±標準差表示。采用單因素方差分析和最小顯著差異(least significant difference,LSD)檢驗進行顯著性分析,其中P<0.05表示差異顯著,P<0.01表示差異極顯著性,P<0.001為差異高度顯著。
2 結(jié)果與分析
2.1 EGCG對3T3-L1脂肪細胞活性及脂代謝的影響
2.1.1 EGCG處理對3T3-L1脂肪細胞活性的影響
為了檢測EGCG對3T3-L1前脂肪細胞的影響,使用EGCG不同質(zhì)量濃度處理3T3-L1前脂肪細胞72 h,與對照組相比,細胞活性顯著增強(圖1A),然后對細胞凋亡相關(guān)的蛋白表達水平進行蛋白免疫印跡分析。結(jié)果顯示,不同質(zhì)量濃度的EGCG對細胞凋亡促進蛋白Bax的表達沒有影響(圖1B、C),而細胞凋亡過程中最主要的Cleaved-caspase 3的表達水平在使用50 μg/mL EGCG 處理72 h后顯著降低(圖1B)。以上研究結(jié)果表明,EGCG可能不會引起3T3-L1成熟脂肪細胞的凋亡。
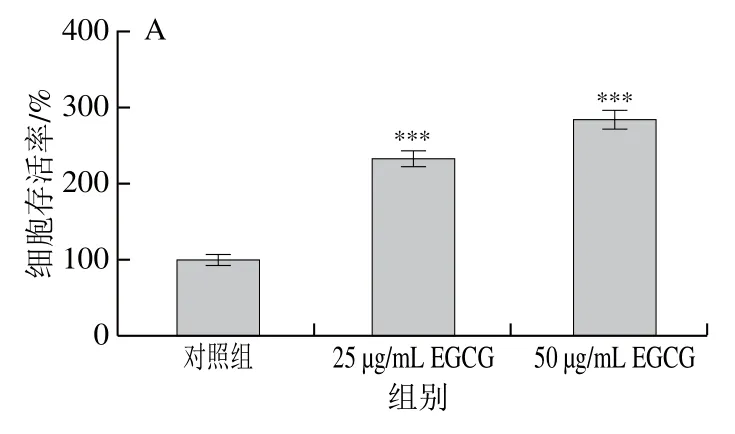

圖1 EGCG處理對3T3-L1成熟脂肪細胞增殖、凋亡相關(guān)蛋白Bax、Cleaved-caspase 3表達水平的影響Fig. 1 Effect of EGCG treatment on cell viability and the expression of Bax and Cleaved-caspase 3 related to apoptosis in mature 3T3-L1 adipocytes
2.1.2 EGCG對成熟3T3-L1脂肪細胞脂滴蓄積的影響
脂肪細胞的數(shù)量和大小決定了人體的胖瘦程度。以脂滴體積增大為特征的脂肪細胞肥大已被證實是導致肥胖者脂肪組織擴張的主要機制。脂滴是一種分布廣泛、進化高度保守的細胞器,由磷脂單分子層、中性脂核和大量脂滴相關(guān)蛋白組成[22-23]。因此在3T3-L1前脂肪細胞誘導分化成為成熟的脂肪細胞后,檢測了EGCG對3T3-L1成熟脂肪細胞脂滴大小的影響。在EGCG處理時間相同的條件下,隨著EGCG處理質(zhì)量濃度的增加,3T3-L1成熟脂肪細胞脂滴數(shù)量以及大小減小,當EGCG處理質(zhì)量濃度為50 μg/mL時,脂滴變小得最為明顯(圖2A)。而在EGCG處理質(zhì)量濃度相同的條件下,隨著處理時間的延長,3T3-L1成熟脂肪細胞脂滴數(shù)量以及大小也減小(結(jié)果未顯示),當EGCG處理時間為144 h時,脂滴變小得最明顯(圖2A)。分析EGCG處理72、144 h后3T3-L1成熟脂肪細胞中的甘油三酯相對含量,可發(fā)現(xiàn)EGCG處理減少了脂肪細胞中甘油三酯的相對含量(圖2B)。以上結(jié)果表明,EGCG的處理可以減小成熟3T3-L1脂肪細胞脂滴的大小并減少甘油三酯的相對含量。

圖2 EGCG處理3T3-L1細胞油紅O染色結(jié)果及甘油三酯相對含量的變化Fig. 2 Oil red O staining and relative content of total triglycerides in 3T3-L1 cells treated with EGCG
2.1.3 EGCG對成熟3T3-L1細胞脂代謝相關(guān)因子mRNA表達水平的影響
為了探討EGCG降低3T3-L1成熟脂肪細胞脂滴大小的具體機制,對3T3-L1成熟脂肪細胞中脂代謝相關(guān)基因的mRNA表達水平進行了檢測(圖3)。EGCG分別處理3T3-L1成熟脂肪細胞72 h和144 h后發(fā)現(xiàn),脂肪合成相關(guān)基因(acc1、srebp1c、fas、scd-1、pparγ、dgat1、elovl6)的mRNA水平的表達整體上顯著下降(圖3A、E)。參與脂肪酸轉(zhuǎn)運的相關(guān)基因(mttp、cd36、fatp1、fatp4)的mRNA水平的表達顯著下降(圖3B、F);與脂滴形成相關(guān)的基因cgi 58的mRNA水平的表達顯著下降(圖3C、G);與脂肪分解相關(guān)的基因atglmRNA水平的表達顯著下降(圖3D、H)。EGCG處理質(zhì)量濃度為50 μg/mL比25 μg/mL下各基因mRNA表達水平下降得更加明顯,結(jié)果表明,EGCG抑制了脂肪合成、脂肪分解、脂肪吸收以及脂滴形成這些脂代謝相關(guān)基因的mRNA水平的表達,且50 μg/mL的EGCG處理抑制效果更加顯著。結(jié)果表明EGCG處理使脂滴大小變小可能與EGCG抑制脂代謝相關(guān)基因的表達有關(guān)。
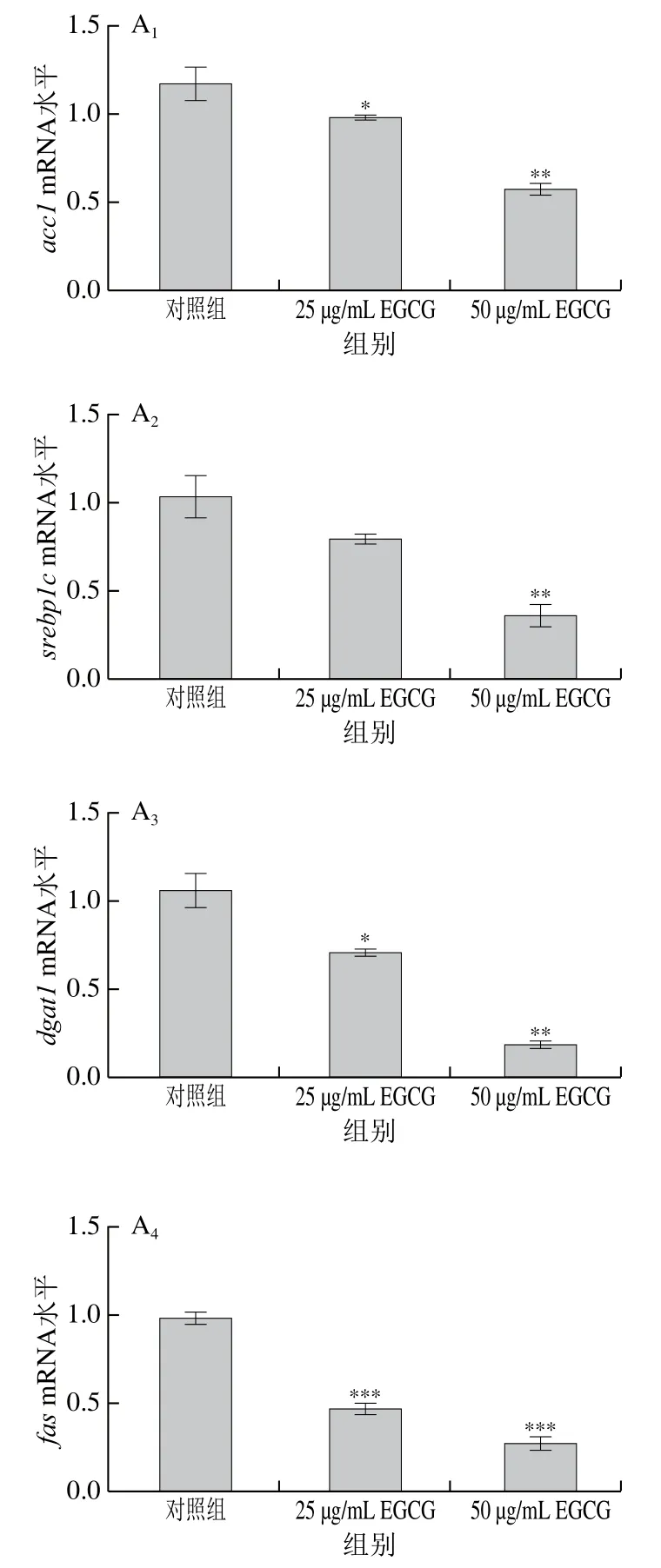
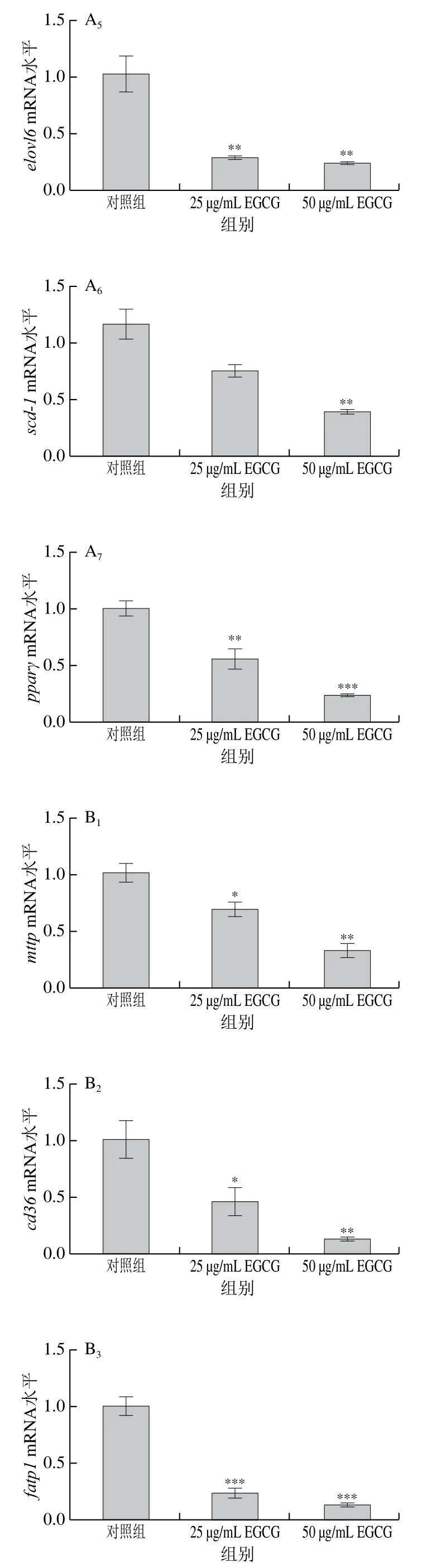

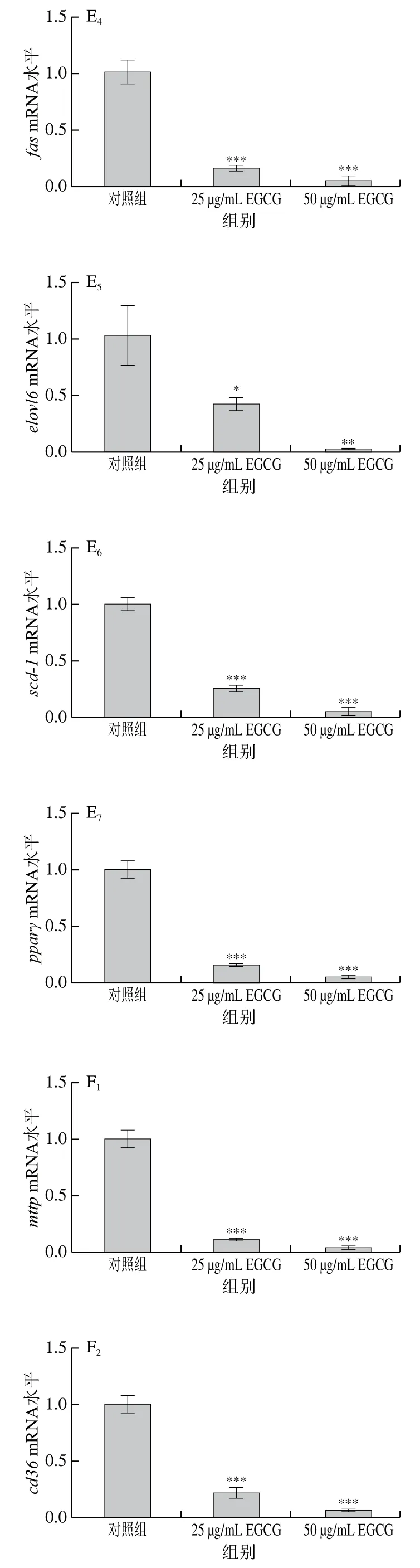
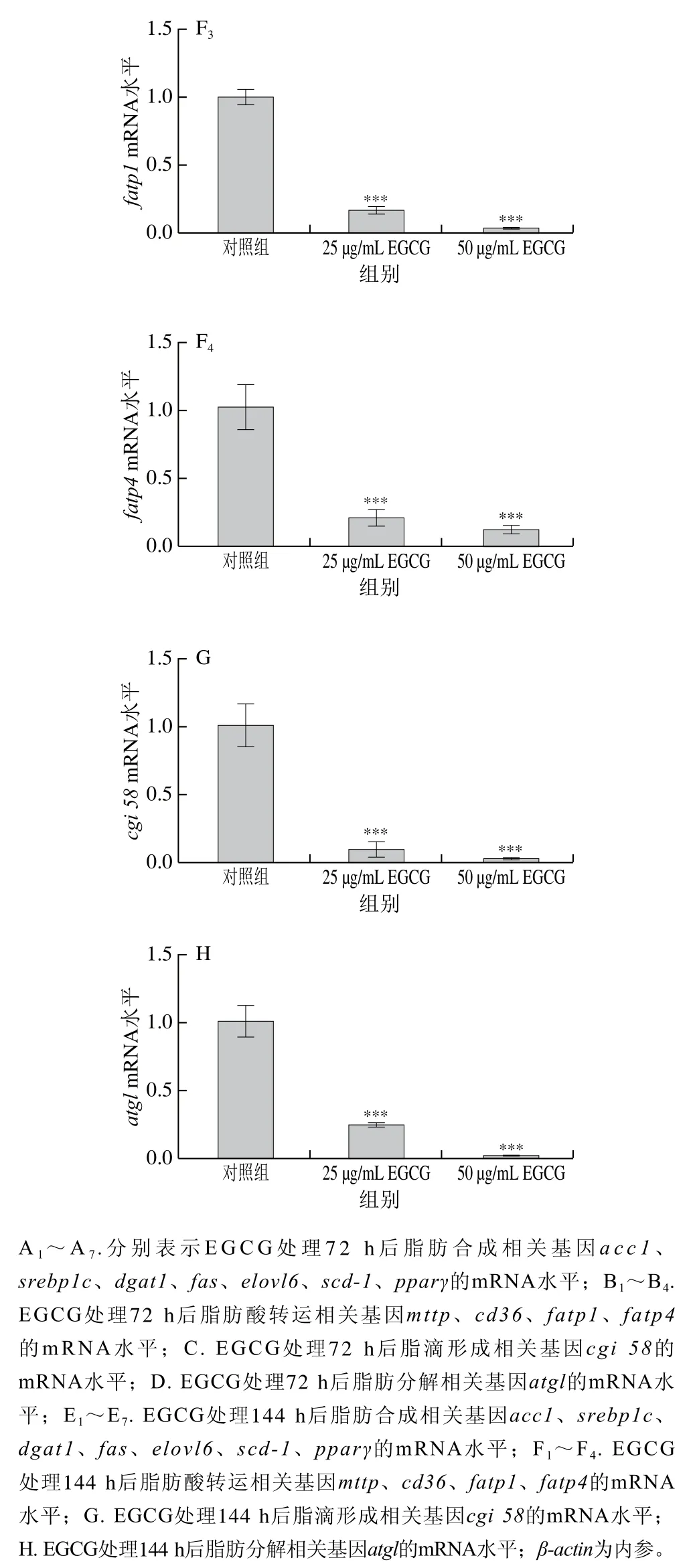
圖3 EGCG對3T3-L1細胞脂代謝相關(guān)基因mRNA水平的影響Fig. 3 Effect of EGCG on mRNA expression levels of genes related to lipid metabolism in 3T3-L1 cells
2.2 EGCG通過激活AMPK抑制成熟3T3-L1脂肪細胞PPARγ的表達
EGCG可以抑制脂代謝相關(guān)基因的表達,但機制并不清楚。Yang等提出AMPK在介導EGCG對脂肪酸合成和分解代謝的作用中發(fā)揮主要作用[6]。因此,在使用EGCG處理3T3-L1成熟脂肪細胞144 h后通過免疫印跡法檢測AMPK的蛋白表達,結(jié)果顯示EGCG能顯著激活AMPK的表達(圖4A)。有文獻報道,AMPK可以通過抑制PPARγ調(diào)控脂肪的合成及儲存[6]。通過檢測PPARγ和其輔活化因子PGC-1α的表達情況發(fā)現(xiàn),EGCG能明顯降低3T3-L1成熟脂肪細胞中PPARγ和PGC-1α的蛋白表達水平(圖4A、B)。另外,PPARγ的蛋白表達水平也隨著EGCG處理質(zhì)量濃度的提高而顯著下降(圖4B)。這些研究結(jié)果表明,EGCG可能通過抑制PPARγ的表達來調(diào)控成熟脂肪細胞中脂滴蓄積。


圖4 EGCG對3T3-L1細胞p-AMPK、AMPK、PPARγ蛋白表達水平的影響Fig. 4 Effect of EGCG on the expression levels of phospho-AMPK,AMPK and PPARγ in 3T3-L1 cells
為了驗證EGCG是否通過激活AMPK來抑制PPARγ的表達,使用AMPK的抑制劑Dorsomorphin處理3T3-L1成熟脂肪細胞。Dorsomorphin處理3T3-L1成熟脂肪細胞后,顯著抑制了p-AMPK的表達(圖4A);而當EGCG和Dorsomorphin同時存在時,EGCG能顯著增加p-AMPK的表達(圖4A)。與對照組相比,Dorsomorphin處理以后,PPARγ的表達顯著增加,而在Dorsomorphin存在下,加入EGCG可以使PPARγ的表達降低(圖4A)。以上結(jié)果表明,EGCG可以使AMPK活化,并降低PPARγ的表達,且通過活化的AMPK來抑制PPARγ的表達。
3 討 論
綠茶能夠預防肥胖,并調(diào)節(jié)脂肪合成和脂肪氧化途徑。綠茶和具有類似性質(zhì)的分子可能成為尋找能量平衡和健康的新靶標[24]。茶葉都有降脂減肥作用,直到近幾十年才發(fā)現(xiàn)是茶葉中單體成分EGCG起著重要的作用[25]。EGCG影響脂代謝的機制仍然是一個有趣的研究領(lǐng)域。本研究的目的是確定EGCG是否可以減少3T3-L1成熟脂肪細胞中的脂滴蓄積并探討其機制。本實驗通過體外實驗,誘導分化培養(yǎng)3T3-L1前脂肪細胞,當EGCG處理質(zhì)量濃度為50 μg/mL、處理時間為144 h時,與對照組相比,脂滴大小顯著減小(圖2A),甘油三酯相對含量降低(圖2B)。而脂滴大小減小是否影響了其脂類代謝以及信號傳導,本研究對脂類代謝相關(guān)因子的表達水平進行了檢測。研究發(fā)現(xiàn),EGCG處理降低了脂肪合成相關(guān)基因的表達,包括acc1、srebp1c、fas、scd-1、pparγ、dgat1、elovl6;降低了脂肪酸轉(zhuǎn)運相關(guān)基因的表達,包括mttp、cd36、fatp1、fatp4;降低了脂肪分解相關(guān)基因atgl的表達;也降低了與脂滴合成相關(guān)的基因cgi 58的表達(圖3),這與之前發(fā)表的動物實驗的結(jié)果[21]一致。
Yang等提出AMPK在介導EGCG對脂肪酸合成和分解代謝的作用中發(fā)揮主要作用[6]。AMPK是機體細胞主要的能量傳感器,參與脂肪酸氧化與葡萄糖轉(zhuǎn)運、甘油三酯的合成與代謝等多種細胞代謝過程[6,24]。本研究發(fā)現(xiàn)EGCG處理3T3-L1成熟脂肪細胞可以激活AMPK(圖4A),那么,AMPK在介導EGCG減小3T3-L1成熟脂肪細胞脂滴大小的過程中是否也發(fā)揮作用呢?本研究結(jié)果顯示EGCG可以降低3T3-L1成熟脂肪細胞的PGC-1α和PPARγ的表達水平(圖4B)。有文獻報道,AMPK可以通過抑制PPARγ調(diào)控脂肪的合成及儲存,而PGC-1α是PPARγ的輔活化子[6,26]。那么,EGCG是否通過激活AMPK抑制PPARγ調(diào)控脂肪的合成及儲存,從而來調(diào)節(jié)脂滴蓄積呢?本研究使用AMPK的抑制劑Dorsomorphin處理3T3-L1成熟脂肪細胞后,發(fā)現(xiàn)AMPK的抑制劑顯著抑制了p-AMPK的表達,而EGCG與AMPK抑制劑共同使用處理細胞時,發(fā)現(xiàn)p-AMPK的表達可以獲得部分恢復(圖4A)。與對照組相比,AMPK的抑制劑Dorsomorphin可以使PPARγ的表達顯著增加,而在Dorsomorphin存在下,加入EGCG可以使PPARγ的表達被顯著抑制(圖4A)。這些研究結(jié)果表明,EGCG通過激活AMPK抑制PPARγ的表達,從而達到減少脂滴蓄積的目的。
在對脂類代謝相關(guān)因子的表達水平進行檢測時發(fā)現(xiàn),EGCG顯著降低了參與脂肪酸轉(zhuǎn)運相關(guān)基因的表達(圖3B),這表明EGCG可以抑制脂肪的吸收。有文獻報道,EGCG可以抑制大鼠脂質(zhì)吸收[27],這與本研究結(jié)果一致,但具體機制仍不清楚。有文獻指出cd36的表達受PPARγ的轉(zhuǎn)錄調(diào)控,因為在cd36啟動子的近端區(qū)域存在PPARγ反應元件[28];PPARγ可以激活fatp和ap2基因的表達,ap2、fatp是PPARγ的靶基因,位于PPARγ下游,受PPARγ調(diào)控,在葡萄糖和脂質(zhì)代謝中起重要作用[29-30]。因此,EGCG可能通過抑制PPARγ從而降低參與脂肪酸轉(zhuǎn)運的相關(guān)基因如cd36、fatp的表達從而抑制了脂肪酸的轉(zhuǎn)運。另外,有文獻報道,CD36可抑制AMPK的活化,敲減cd36可以顯著增強AMPK的活化水平[31-32]。因此,根據(jù)上述討論內(nèi)容,推測EGCG調(diào)節(jié)脂肪細胞脂滴大小的機制可能是通過增強AMPK的活化,來抑制PPARγ的表達,從而抑制與脂肪合成相關(guān)基因的表達和脂肪吸收相關(guān)基因的表達,來降低脂滴的大小;另一方面,cd36基因表達的下調(diào)又反過來增強AMPK的活化水平(圖5)。
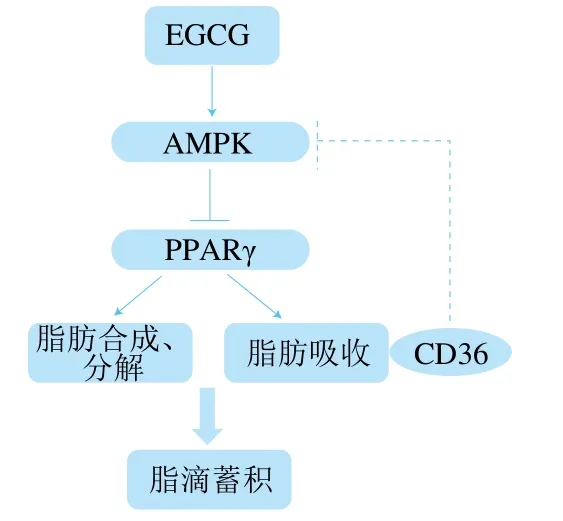
圖5 EGCG調(diào)控3T3-L1成熟脂肪細胞中脂滴蓄積機制示意圖Fig. 5 Schematic diagram of the mechanism of EGCG regulating lipid droplet accumulation in 3T3-L1 mature adipocytes
綜上,EGCG處理3T3-L1成熟脂肪細胞可以降低脂滴大小和脂肪細胞甘油三酯相對含量,其降低脂滴大小的機制可能與增強AMPK活性有關(guān)。一方面,AMPK活性增加使PPARγ的表達降低從而調(diào)控脂肪合成和分解,進而減小脂滴大小;另一方面,AMPK通過抑制PPARγ從而抑制cd36、fatp1、fatp4這些與脂肪吸收相關(guān)基因的表達,減少脂肪吸收進而減少脂肪細胞中脂滴蓄積。
猜你喜歡
中學生數(shù)理化·中考版(2022年10期)2022-11-10 09:37:42
中學生數(shù)理化·八年級物理人教版(2022年12期)2022-02-14 07:08:42
中學生數(shù)理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
中學生數(shù)理化·中考版(2020年10期)2020-11-27 01:59:48
中國生殖健康(2019年2期)2019-08-23 08:12:08
石油化工建設(shè)(2018年6期)2018-04-22 03:16:54
產(chǎn)品可靠性報告(2017年7期)2017-09-05 09:49:12
中學生數(shù)理化·八年級物理人教版(2017年12期)2017-04-18 12:59:38
汽車觀察(2016年3期)2016-02-28 13:16:26
民生周刊(2014年7期)2014-03-28 01:30:54
