甘露寡糖對養殖魚類生長性能和非特異性免疫的研究進展
2021-12-18 02:28:44翟浩杰任同軍王福強韓雨哲
安徽農業科學 2021年23期
翟浩杰 任同軍 王福強 韓雨哲
摘要 由于抗生素的使用被限制,新型綠色飼料添加劑成為水產動物營養與飼料的研究熱點,甘露寡糖作為一種功能性寡糖被認為是抗生素的替代品,研究發現,甘露寡糖能夠促進動物生長,改善腸道健康并增強其非特異免疫功能。為此,綜述了甘露寡糖對魚類生長性能、腸道健康和非特異性免疫能力的影響,對其在水產養殖中的應用做出展望。
關鍵詞 甘露寡糖;魚類;生長性能;腸道健康;非特異性免疫
中圖分類號 S 963.73? 文獻標識碼 A
文章編號 0517-6611(2021)23-0017-05
doi:10.3969/j.issn.0517-6611.2021.23.005
Research Progress of Mannan Oligosaccharides on Growth Performance and Nonspecific Immunity of Cultured Fish
ZHAI Hao-jie1,REN Tong-jun1,2,WANG Fu-qiang1,2 et al
(1.College of Fisheries and Life Sciences,Dalian Ocean University,Dalian,Liaoning 116023;2.Key Laboratory of Applied Biology and Aquaculture of Northern Fishes in Liaoning Province,Dalian Ocean University,Dalian,Liaoning 116023)
Abstract Due to the limited use of antibiotics,new green feed additives have become the research hotspot of aquatic animal nutrition and feed.As a functional oligosaccharide,mannan oligosaccharide is considered as the substitute of antibiotics.Studies have found that mannan oligosaccharide can promote animal growth,improve intestinal health and enhance its non-specific immune function.Therefore,the effects of mannan oligosaccharides on growth performance,intestinal health and non-specific immunity of fish were reviewed,and its application in aquaculture was expected.
Key words Mannan oligosaccharides;Fish;Growth performance;Intestinal health;Non-specific immunity
作者簡介 翟浩杰(1997—),男,陜西寶雞人,碩士研究生,研究方向:水產動物營養與飼料。通信作者,副教授,博士,從事水產動物健康養殖和水產動物營養飼料學研究。
收稿日期 2021-03-25
近年來我國魚類養殖業快速發展,高密度集約化的水產養殖導致的魚類病害成為了影響漁業經濟的重要因素之一,傳統方法采用抗生素對病害進行防治,但由于抗生素等化學藥物會對自然環境造成污染,而且容易在水產品中殘留,嚴重影響食物安全和人類健康,因此,目前主要通過一些無毒無害無殘留、結構穩定的飼料添加劑(寡糖、多糖、益生菌、氨基酸、維生素等)來提高動物的抗病能力和生產性能。
寡糖作為一種能源物質,自20世紀80年代被開發為飼料添加劑以來,一直是動物營養免疫方面研究的熱點。寡糖又稱低聚糖或寡聚糖,是由2~10個單糖單位經脫水縮合由糖苷鍵連接而成的小聚合體,寡糖又分為普通寡糖和功能性寡糖。甘露寡糖(mannan oligosaccharides,MOS)又稱甘露低聚糖或葡甘露寡聚糖,是功能性寡糖的一類,是從酵母培養細胞壁中提取的一類新型抗原活性物質,廣泛存在于魔芋粉、瓜兒豆膠、田菁膠及多種微生物細胞壁內。甘露寡糖是由幾個甘露糖分子或甘露糖與葡萄糖通過α-1,2、α-1,3和α-1,6糖苷鍵組成的寡聚糖,易溶于水和極性溶劑,一般在生理pH和飼料加工制粒時較為穩定,能夠保持其結構和功能的完整性。甘露寡糖作為一種新型綠色飼料添加劑,已經廣泛應用于養殖動物飼料的開發,而作為飼料添加劑的甘露寡糖一般為二糖、三糖和四糖的混合物[1],研究表明甘露寡糖可以提高養殖動物的生產性能、改善腸道微生物區系以及提高免疫功能等[2-4]。大量研究表明,甘露寡糖具有促進魚類生長性能,改善魚類腸道健康,提高其非特異性免疫功能的作用。
1 甘露寡糖對魚類生長性能的影響
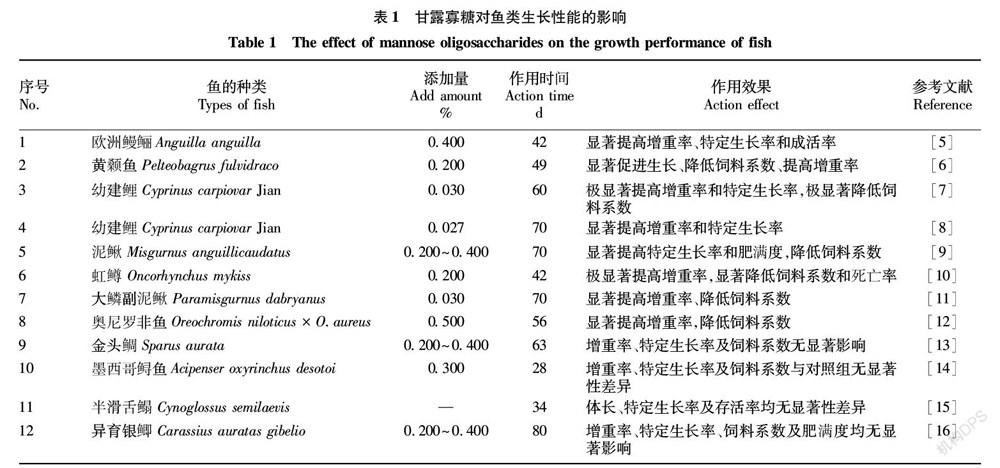
國內外關于甘露寡糖(MOS)對魚類生長性能影響的研究結果尚存在差異,部分研究表明在魚類日糧中添加甘露寡糖可以提高魚類的生長性能,其作用方式主要通過提高魚類體增重率、降低飼料系數等方面來實現(表1)。在對歐洲鰻鱺(Anguilla anguilla)[5]、黃顙魚(Pelteobagrus fulvidraco)[6]、幼建鯉(Cyprinus carpiovar Jian)[7-8]、泥鰍(Misgurnus anguillicaudatus)[9]、虹鱒(Oncorhynchus mykiss)[10]及大鱗副泥鰍(Paramisgurnus dabryanus)[11]的研究中均發現,甘露寡糖能夠通過提高增重率、特定生長率,降低飼料系數等來提高魚類的生長性能。但在對奧尼羅非魚(Oreochromis niloticus × O.aureus)[12]的研究中發現,甘露寡糖在魚類飼料中的添加量有一個合適的范圍,過量的添加可能會對魚類的生長產生負面影響,原因可能是較高濃度的甘露寡糖促進腸道絨毛高度過度發育導致腸道阻塞,抑制魚類對營養物質的消化吸收,從而對魚類的生長性能產生了負面影響。日糧中添加甘露寡糖有助于動物的生長性能,其作用機制可能歸因于改善了腸道健康,在一定的添加濃度范圍內,甘露寡糖促進腸道絨毛高度、改善腸道菌群、提高腸道消化酶活力,導致腸道吸收面積增加、腸道菌群結構平衡、消化能力增強,營養物質吸收效率提高,從而促進魚類的生長性能。
也有一些與之相反的研究,在金頭鯛(Sparus aurata)[13]、墨西哥鱘魚(Acipenser oxyrinchus desotoi)[14]、半滑舌鰨(Cynoglossus semilaevis)[15]及異育銀鯽(Carassius auratas gibelio)[16]的研究上發現,基礎飼料中添加甘露寡糖對其增重率、特定生長率、飼料系數等生長指標均無顯著性影響。原因可能是試驗魚類種類或魚體生長發育階段不同,對試驗結果產生了不同的影響;試驗過程差異,試驗者選擇甘露寡糖的作用時間、添加量、養殖環境以及飼養周期的長短都有可能導致甘露寡糖對魚類的生長作用產生差異。但從總體來看,甘露寡糖能夠提高魚類的生長性能。
2 甘露寡糖對魚類腸道功能的影響
2.1 甘露寡糖對魚類腸道菌群結構的影響
魚類腸道內定植著數量龐大的微生物群落,主要分為有益微生物菌群(乳酸桿菌屬、雙歧桿菌屬等)和有害微生物菌群(大腸桿菌、嗜水氣單胞菌屬等)[17],這些微生物菌群共同組成了魚體腸道的微生物區系,直接影響著機體的營養和健康。大量研究結果表明,在魚類日糧中添加甘露寡糖與魚類腸道菌群平衡之間有著直接的關系。張紅梅[18]在鯉魚(Cyprinus carpio)的基礎飼料中添加甘露寡糖,隨著劑量的增加,魚體腸道中的大腸桿菌數量減少與對照組有顯著性差異(P<0.01),雙歧桿菌數量增加與對照組相比差異極顯著(P<0.01),乳酸桿菌數量增加與對照組相比差異不顯著(P>0.05)。李云蘭[8]在幼建鯉基礎飼料中添加甘露寡糖,試驗結果表明,當添加含量在180~270 mg/kg時,幼建鯉前、中和后腸大腸桿菌數量與對照組相比極顯著降低(P<0.01);當甘露寡糖含量在270 mg/kg時,幼建鯉腸道中乳酸桿菌和雙歧桿菌濃度相比對照組有顯著性提高(P<0.05)。Torrecillas[19]在歐洲海鱸(Dicentrarchus labrax)的基礎飼料中添加4 g/kg的甘露寡糖,8周試驗后結果顯示,甘露寡糖能夠顯著提高海鱸腸道中乳酸桿菌和雙歧桿菌的數量,且顯著降低了大腸桿菌和嗜水氣單胞菌的數量。汪立平等[20]在奧尼羅非魚飼料中添加自制酵母甘露寡糖,30 d試驗結果表明在羅非魚飼料中添加0.5%的甘露寡糖能夠顯著提高其腸道中的乳酸桿菌數量,降低腸道中大腸桿菌數量。蘇健[21]在泥鰍的日糧中添加甘露寡糖,試驗結果表明,當添加量為200和400 mg/kg時,甘露寡糖顯著降低了泥鰍腸道中大腸桿菌的數量,且顯著增加了腸道中乳酸桿菌和雙歧桿菌的數量。甘露寡糖可以與有害細菌的受體競爭結合,降低有害細菌在腸道定值的幾率,加速有害細菌從腸道中排出,同時這種競爭性排斥作用也有利于有益菌的生長,此外甘露寡糖還能夠通過降低腸道pH值,抑制有害菌的生長[22]。因此,甘露寡糖對腸道菌群的調節主要在于促進腸道有益菌增殖抑制有害菌,優化腸道菌群結構,促進腸道微生物區系的平衡,從而提高動物的生長性能和免疫能力。
2.2 甘露寡糖對魚類腸道形態結構的影響
腸道是魚類容納消化食物和運輸吸收營養物質的主要場所,腸道健康是魚類正常生長發育的重要保證[23]。腸道形態是最能直觀反映魚類腸道健康的指標,腸道絨毛高度、肌層厚度與腸道的吸收能力相關,直接影響著魚體的消化吸收營養物質的能力[24]。小腸絨毛的主要功能是吸收營養物質,小腸絨毛高度的變化決定了腸道有效吸收面積的大小,腸道皺襞高度寬度的增加也有利于營養物質的消化吸收,體現著腸道的健康狀況。甘露寡糖能夠改變腸道形態結構,可以增加豬[25]、兔[26]、雞[27]等畜禽動物的小腸絨毛高度和寬度。在水產動物的研究中,胡凌豪等[28]在斜帶石斑魚(Epinephelus coioides)基礎飼料中添加不同劑量的果寡糖,試驗結果表明,果寡糖能夠增加斜帶石斑魚的腸道絨毛高度和寬度,且0.1%試驗組腸道的肌層厚度顯著高于對照組(P<0.05);劉愛君等[12]在奧尼羅非魚的基礎飼料中添加甘露寡糖,對各組腸道做組織切片后發現,添加0.25%、0.50%和0.75%的甘露寡糖均能顯著增加腸道絨毛高度、絨毛寬度和絨毛密度(P<0.05),且0.50%組的效果最顯著;徐磊[16]在異育銀鯽的日糧中添加甘露寡糖養殖80 d,120 mg/kg組的腸絨毛長度相比對照組提高了8.1%,但各試驗組腸絨毛長度與腸直徑的比值、皺襞間質寬、黏膜下層寬、肌層寬與對照組無顯著性差異(P>0.05);Torrecillas等[19] 在歐洲海鱸的基礎飼料中添加4 g/kg的甘露寡糖,試驗結束后與對照組相比試驗組魚體腸道前腸黏膜的褶皺高度,寬度和褶皺表面積都有顯著性增加(P<0.05),后腸總表面積增加(P<0.05),但后腸褶皺較對照組短(P<0.05);Dimitroglou等[29]在虹鱒魚的基礎飼料中添加0.2%的甘露寡糖,用光鏡和電子顯微鏡觀察前腸和后腸形態發現,與對照組相比試驗組的腸道吸收面積增加,微絨毛長度和密度顯著增加(P<0.05),但在仔魚的研究上結果差異并不顯著,可能的原因是甘露寡糖對不同生長發育階段魚體的影響有差異。Dimitroglou等[30]在白鯛(Diplodus sargus L.)的鹵蟲飼料中添加甘露寡糖,結果表明,與對照組相比添加甘露寡糖可將其微絨毛長度增加26%;在大菱鲆腸道健康(Scophthalmus maximus)的研究上,發現在基礎飼料中添加甘露寡糖能改善腸道絨毛高度和黏膜皺襞,抑制微絨毛和黏膜皺襞的改變,改善大菱鲆腸道健康狀況[31]。甘露寡糖對魚類腸道形態結構產生有益影響,腸道結構的改善導致營養物質消化率和飼料利用率的提高,從而促進魚類的生長性能。Pryor等[14]在墨西哥鱘魚飼料中添加0.3%的甘露寡糖,26 d試驗結果表明MOS對其腸道絨毛高度、寬度和密度均無顯著性影響;Salem等[32]在歐洲黑鱸日糧中添加的0.2%、0.4%甘露寡糖對腸道絨毛長度沒有影響。以上差異可能是由于甘露寡糖的添加量不同、魚的種類不同、魚類機體腸道內的腸道微生物群不同或者試驗方法不同等。
2.3 甘露寡糖對魚類腸道消化酶的影響
動物機體的消化能力除了與腸道生態環境、形態結構相關聯外還與腸道中消化酶的活力密切聯系。魚類腸道消化酶是反映機體消化能力和吸收營養成分的重要指標。研究表明功能性寡糖能夠促進腸道上皮細胞生長,提高內源酶活性,促進魚類生長發育。楊敏等[5]在歐洲鰻鱺的飼料中添加不同含量的甘露寡糖,42 d試驗后發現,甘露寡糖能夠提高歐洲鰻鱺腸道胰蛋白酶、淀粉酶和脂肪酶活性,0.4%組胰蛋白酶、脂肪酶和淀粉酶活性與對照組有顯著性差異(P<0.05);強俊等[33]在奧尼羅非魚幼魚飼料中添加不同水平的低聚木糖(XOS),結果表明添加低聚木糖組提高了奧尼羅非魚幼魚腸道淀粉酶和脂肪酶活力,分別在0.030%和0.045%最高,且各組之間有顯著性差異(P<0.05);李洋[7]在幼建鯉基礎飼料中添加甘露寡糖,試驗結果表明,添加量為300 mg/kg時糜蛋白酶和腸淀粉酶極顯著高于對照組(P<0.05),腸脂肪酶顯著高于對照組(P<0.05),但胰蛋白酶與對照組差異不顯著(P>0.05);此外,Safai等[34]在小龍蝦日糧中分別添加甘露寡糖和低聚果糖(FOS)及其組合,結果表明,MOS和FOS及組合添加對其生長性能、消化酶活力等均有顯著性提高,但MOS和FOS的組合添加比起單獨添加作用效果更好。
3 甘露寡糖對魚類免疫功能的影響
動物機體的免疫系統包括特異性免疫和非特異性免疫,非特異性免疫系統是阻止病原菌入侵機體的第一道防線,在魚類機體中發揮著重要作用,對其生長和抵御病原入侵有著重要的影響[35]。當病原微生物和異物入侵時,非特異性免疫通過物理屏障、吞噬作用、溶菌作用和凝集作用等清除病原菌和異物[36]。魚類非特異性免疫能力的提高主要表現在抑菌類物質的產生,吞噬細胞吞噬能力及相關免疫酶活力等方面的提高。
3.1 甘露寡糖對魚類免疫酶活力的影響
溶菌酶是非特異性免疫系統的重要組成,是存在于魚類體表黏液、血清和巨噬細胞中的一種水解酶,在動物機體排斥異物攻擊時發揮著重要作用;酸性磷酸酶存在于巨噬細胞溶酶體,是巨噬細胞內最具代表性的水解酶之一,能夠快速對異物識別、吞噬和清除達到預防感染的目的[37];堿性磷酸酶廣泛存在于動物血液和各種器官內,能夠通過改變病原體表面結構,增強機體對病原體的識別和吞噬能力[38]。因此其酶活力可作為免疫指標來間接反映魚類非特異免疫功能。大量研究表明,甘露寡糖可以增強魚類免疫酶活力,Lu等[39]在草魚(Ctenopharyngodon idella)基礎飼料中添加甘露寡糖,研究結果表明MOS可以增強其溶菌酶和磷酸酶活力;張紅梅等[40]在鯉魚的基礎飼料中添加不同水平的甘露寡糖,結果顯示隨著添加量的增加血清中溶菌酶活力呈增長趨勢,其中0.3%添加組與對照組有極顯著差異;劉愛君等[41]報道在奧尼羅非魚基礎飼料中添加甘露寡糖能顯著提高其血清溶菌酶活力和血清堿性磷酸酶活力;Ayce Genc等[42]在羅非魚(Oreochromis mossambicus)基礎飼料中添加3 g/kg的甘露寡糖顯著提高了其血清溶菌酶活力;王銳等[43]在異育銀鯽幼魚飼料中添加半乳甘露寡糖,試驗結果表明甘露寡糖顯著提高了異育銀鯽幼魚血清溶菌酶的活力。此外,Meng等[44]在甘露寡糖對皺紋盤鮑(Haliotis discus hannai Ino)幼鮑的研究上也發現,甘露寡糖能顯著提高其肝胰臟和血清中堿性磷酸酶、酸性磷酸酶和溶菌酶活力。因此,甘露寡糖可以提高魚類的免疫酶活性,進而增強其非特異性免疫功能。甘露寡糖能夠優化腸道微生物區系促進有益菌的增殖,而雙歧桿菌和乳酸桿菌的增殖提高了機體抗體水平,促進了巨噬細胞的吞噬能力[45],從而提高相關免疫酶活性對機體非特異性免疫能力產生有益影響。
3.2 甘露寡糖對魚類紅細胞數量的影響
紅細胞是魚類機體中重要的免疫細胞,能夠影響機體的吞噬能力,在魚類的非特異性免疫防御中發揮著重要作用[46]。李云蘭[7]發現甘露寡糖極顯著的影響了幼建鯉血液紅細胞數量(P<0.01),且當甘露寡糖添加量在619 mg/kg時,幼建鯉血液紅細胞數量最多增加58.23%;Andrews等[47]在南亞野鱗幼魚(Labeo rohita)飼料中添加1%的甘露寡糖,結果表明其血液紅細胞數量相比對照組提高了5.3%;李洋[7]報道添加甘露寡糖極顯著增加了幼建鯉血液紅細胞數量。此外,Rodriguez-Estrada等[48]在虹鱒日糧中分別添加糞腸球菌(EF)和甘露寡糖(MOS)及其組合,結果表明EF和MOS以及聯合使用顯著提高了虹鱒的紅細胞壓積值,但分析結果表明EF和MOS的聯合添加對虹鱒的紅細胞壓積值、粘液重量等免疫指標的提升達到最佳水平。上述研究結果說明,甘露寡糖能夠通過提高魚類血液紅細胞數量來增強機體非特異免疫能力。
3.3 甘露寡糖對魚類白細胞吞噬率的影響
水生動物體內的吞噬細胞主要包括中性粒細胞和單核巨噬細胞,是白細胞的主要成分。魚類血液中白細胞吞噬能力可以反映出機體的免疫狀態,白細胞吞噬能力一般用白細胞吞噬率和吞噬指數來表示。魚類機體吞噬細胞對病原微生物的吞噬和殺滅作用,是其非特異性免疫的一個重要方面[49]。王銳等[43]研究發現半乳甘露寡糖組的異育銀鯽白細胞數目、吞噬細胞百分比均顯著高于對照組(P<0.05),甘露寡糖組的吞噬指數相比對照組顯著提高(P<0.05),且0.2%添加組最高;馬志紅等[50]在鯉基礎飼料中添加0.1%的甘露寡糖,結果表明試驗組相比對照組白細胞吞噬率、吞噬指數均顯著增加(P<0.01);李洋[7]研究發現甘露寡糖極顯著提高了幼建鯉血液白細胞吞噬率(P<0.01);Torrecillas等[51]發現甘露寡糖可以顯著提高歐洲海鱸的白細胞數量和白細胞吞噬率;Yoshida等[52]發現在大西洋鯰(Clarias gariepinus)基礎飼料中添加甘露寡糖能夠顯著提高其嗜中性粒細胞的吞噬活性。因此,甘露寡糖能夠通過提高魚類的白細胞吞噬率從而提高其非特異性免疫功能。Spring[53]研究表明吞噬細胞吞噬能力提高的原因可能是甘露寡糖促進了輔助性T-淋巴細胞的活性,甘露寡糖能夠增強輔助性T-淋巴細胞分泌白細胞介素-2(IL-2)和γ-干擾素(IFN-γ)的能力,從而對機體的非特性免疫能力產生積極的影響。
4 小結和展望
作為一種綠色飼料添加劑,甘露寡糖能夠提高動物機體的生長性能,促進腸道發育,優化腸道菌群結構并增強其非特異免疫功能,能夠廣泛應用在動物飼料行業,是抗生素的良好替代品。但甘露寡糖在水產養殖中的應用仍有一些問題有待解決:
(1)關于甘露寡糖對水產動物的研究還不夠全面,未來仍然需要探索甘露寡糖在更多經濟水產動物飼料中的適宜添加量、不同生長發育階段的使用方式、不同種類甘露寡糖的使用效果等,從而使其在水產養殖中得到更深入廣泛地應用。
(2)甘露寡糖與其他飼料添加劑的聯合使用。已有部分試驗表明甘露寡糖與其他添加劑的聯合使用比起單獨添加對水產動物的作用效果似乎更好,因此,未來應該致力于功能性寡糖之間或者與其他營養素之間相關性的研究,以期更加有效率地使用飼料添加劑,從而實現高效率、低成本、無污染的水產養殖。
參考文獻
[1]鮑延安,邢淑芳,徐慶龍.甘露寡糖對荷斯坦奶牛產奶量及乳常規的影響[J].飼料研究,2009(2):57-60.
[2] 熊阿玲.日糧添加甘露寡糖對肉雞生長性能及天然免疫相關基因表達的影響研究[D].南昌:江西農業大學,2014.
[3] CHACHER M F A,KAMRAN Z,AHSAN U,et al.Use of mannan oligosaccharide in broiler diets:An overview of underlying mechanisms[J].World′s poultry science journal,2017,73(4):831-844.
[4] AGAZZI A,PERRICONE V,ZORINI F O,et al.Dietary mannan oligosaccharides modulate gut inflammatory response and improve duodenal villi height in post-weaning piglets improving feed efficiency[J].Animals,2020,10(8):1-14.
[5] 楊敏,黎中寶,盧靜,等.甘露寡糖對歐洲鰻鱺(Anguilla anguilla)生長、消化酶活性及非特異性免疫的影響[J].中國漁業質量與標準,2016,6(6):40-46.
[6] 于艷梅,吳志新,陳孝煊,等.魔芋甘露寡糖對黃顙魚非特異性免疫功能及生長的影響[J].華中農業大學學報,2010,29(3):351-355.
[7] 李洋.甘露寡糖對幼建鯉生長性能和非特異性免疫的影響[D].雅安:四川農業大學,2011.
[8] 李云蘭.甘露寡糖對幼建鯉(Cyprinus carpio Var.Jian)腸道菌群和免疫功能的影響[D].雅安:四川農業大學,2004.
[9] 杜宗君,蘇健,巫紅萍,等.甘露寡糖對泥鰍生產性能、腸道菌群和非特異性免疫功能的影響[J].四川農業大學學報,2019,37(1):129-133.
[10] STAYKOV Y,SPRING P,DENEV S,et al.Effect of a mannan oligosaccharide on the growth performance and immune status of rainbow trout (Oncorhynchus mykiss)[J].Aquaculture international,2007,15(2):153-161.
[11] XU B,WU S J,HAN Q.Modulation of the growth performance and innate immunity of loaches (Paramisgurnus dabryanus) upon dietary mannan oligosaccharides[J].3 Biotech,2021,11(3):1-7.
[12] 劉愛君,冷向軍,李小勤,等.甘露寡糖對奧尼羅非魚(Oreochromis niloticus×O.aureus)生長、腸道結構和非特異性免疫的影響[J].浙江大學學報(農業與生命科學版),2009,35(3):329-336.
[13] DIMITROGLOU A,MERRIFIELD D L,SPRING P,et al.Effects of mannan oligosaccharide (MOS) supplementation on growth performance,feed utilisation,intestinal histology and gut microbiota of gilthead sea bream (Sparus aurata)[J].Aquaculture,2010,300(1/2/3/4):182-188.
[14] PRYOR G S,ROYES J B,CHAPMAN F A,et al.Mannanoligosaccharides in fish nutrition: Effects of dietary supplementation on growth and gastrointestinal villi structure in gulf of Mexico sturgeon[J].North American journal of aquaculture,2003,65(2):106-111.
[15] 于朝磊,常青,呂云云.甘露寡糖對半滑舌鰨(Cynoglossus semilaevis Günther)稚魚生長、腸道發育和非特異性免疫水平的影響[J].漁業科學進展,2014,35(6):53-59.
[16] 徐磊.甘露寡糖對異育銀鯽生長性能及抗嗜水氣單胞菌感染的研究[D].南京:南京農業大學,2010.
[17] LLEWELLYN M S,BOUTIN S,HOSEINIFAR S H,et al.Teleost microbiomes: The state of the art in their characterization,manipulation and importance in aquaculture and fisheries[J].Frontiers in microbiology,2014,5:1-17.
[18] 張紅梅.甘露寡聚糖對鯉魚生產性能及對魚體各項生物學指標的影響[D].保定:河北農業大學,2003.
[19] TORRECILLAS S,MAKOL A,BENTEZ-SANTANA T,et al.Reduced gut bacterial translocation in European sea bass (Dicentrarchus labrax) fed mannan oligosaccharides (MOS)[J].Fish & shellfish immunology,2011,30(2):674-681.
[20] 汪立平,馬相杰,冷向軍,等.甘露寡糖對羅非魚腸道菌群的影響[J].食品工業科技,2010,31(10):142-145.
[21] 蘇健.甘露寡糖對泥鰍生產性能、腸道菌群和非特異性免疫功能的影響[D].雅安:四川農業大學,2013.
[22] 李鳳娜,王繼成.寡糖對腸道菌群的調節及常用研究方法[J].獸藥與飼料添加劑,2005,10(4):23-25.
[23] 涂永鋒,宋代軍.魚類腸道組織結構及其功能適應性[J].江西飼料,2004(4):16-19.
[24] 李霞.水產動物組織胚胎學[M].北京:中國農業出版社,2006.
[25] 黃俊文,林映才,馮定遠,等.納豆菌、甘露寡糖對仔豬腸道pH、微生物區系及腸黏膜形態的影響[J].畜牧獸醫學報,2005,36(10):1021-1027.
[26] MOURO J L,PINHEIRO V,ALVES A,et al.Effect of mannan oligosaccharides on the performance,intestinal morphology and cecal fermentation of fattening rabbits[J].Animal feed science & technology,2006,126(1/2):107-120.
[27] IJI? P A,SAKI A A,TIVEY D R.Intestinal structure and function of broiler chickens on diets supplemented with a mannan oligosaccharide[J].Journal of the science of food & agriculture,2001,81(12):1186-1192.
[28] 胡凌豪,楊紅玲,趙蕓,等.果寡糖對斜帶石斑魚免疫功能和腸道形態的影響[J].水產科學,2019,38(5):589-594.
[29] DIMITROGLOU A,MERRIFIELD D L,MOATE R,et al.Dietary mannan oligosaccharide supplementation modulates intestinal microbial ecology and improves gut morphology of rainbow trout,Oncorhynchus mykiss(Walbaum)[J].Journal of animal science,2009,87(10):3226-3234.
[30] DIMITROGLOU A,DAVIES S J,SWEETMAN J,et al.Dietary supplementation of mannan oligosaccharide on white sea bream (Diplodus sargus L.) larvae:Effects on development,gut morphology and salinity tolerance[J].Aquaculture Research,2010,41(9):e245-e251.
[31] BAI N,GU M,XU X J,et al.Protective effects of mannan oligosaccharides on turbot Scophthalmus maximus suffering from soy enteropathy[J].Aquaculture,2017,476:141-151.
[32] SALEM M,GABER M M,ZAKI M A D,et al.Effects of dietary mannan oligosaccharides on growth,body composition and intestine of the sea bass ( Dicentrarchus labrax L.)[J].Aquaculture research,2016,47(11):3516-3525.
[33] 強俊,王輝,李瑞偉,等.低聚木糖對奧尼羅非魚幼魚生長、體成分和消化酶活力的影響[J].淡水漁業,2009,39(6):63-68.
[34] SAFARI O,SHAHSAVANI D,PAOLUCCI M,et al.Single or combined effects of fructo-and mannan oligosaccharide supplements on the growth performance,nutrient digestibility,immune responses and stress resistance of juvenile narrow clawed crayfish,Astacus leptodactylus leptodactylus Eschscholtz,1823[J].Aquaculture,2014,432:192-203.
[35] 錢云霞,王國良,邵健忠.魚類的非特異性免疫調節[J].寧波大學學報(理工版),2000,13(1):95-99.
[36] 慈麗寧,劉波,謝駿,等.影響水生動物免疫機能的因素(綜述)[J].江蘇農業科學,2012,40(3):205-210.
[37] CHENG T C.The role of lysosomal hydrolases in molluscan cellular response to immunologic challenge[M]//BULLA L A JR,CHENG T C.Invertebrate models for biomedical research.New York:Plenum Press,1978.
[38] 牟海津,江曉路,劉樹青,等.免疫多糖對櫛孔扇貝酸性磷酸酶、堿性磷酸酶和超氧化物歧化酶活性的影響[J].青島海洋大學學報(自然科學版),1999,29(3):463-468.
[39] LU Z Y,JIANG W D,WU P,et al.Mannan oligosaccharides supplementation enhanced head-kidney and spleen immune function in on-growing grass carp (Ctenopharyngodon idella)[J].Fish & shellfish immunology,2020,106(4):596-608.
[40] 張紅梅,姜會民.酵母甘露寡糖對鯉魚非特異性免疫的影響[J].飼料研究,2006(10):25-27.
[41] 劉愛君,冷向軍,李小勤,等.黃霉素和甘露寡糖對奧尼羅非魚的生長性能及血清非特異性免疫的影響[J].中國飼料,2009(3):29-32.
[42] AYCE GENC M,YILMAZ E,GENC E,et al.Effects of dietary mannan oligosaccharides (MOS) on growth,body composition,and intestine and liver histology of the hybrid tilapia (Oreochromis niloticus×O.aureus)[J].The Israeli journal of aquaculture-bamidgeh,2007,59(1):10-16.
[43] 王銳,劉軍,劉輝宇,等.半乳甘露寡糖對異育銀鯽幼魚生長和非特異性免疫的影響[J].上海水產大學學報,2008,17(4):502-506.
[44] MENG X X,YANG X Y,LIN G,et al.Mannan oligosaccharide increases the growth performance,immunity and resistance capability against Vibro Parahemolyticus in juvenile abalone Haliotis discus hannai Ino[J].Fish & shellfish immunology,2019,94:654-660.
[45] 唐凌,鄺聲耀,馮琳,等.甘露寡糖對水生動物腸道菌群和免疫功能的影響[J].中國畜牧獸醫,2008,35(6):21-25.
[46] 蔡完其,軒興榮.紅鯉4群體間紅細胞免疫功能及其差異[J].中國水產科學,2003(2):133-136.
[47] ANDREWS S R,SAHU N P,PAL A K,et al.Haematological modulation and growth of Labeo rohita fingerlings:Effect of dietary mannan oligosaccharide,yeast extract,protein hydrolysate and chlorella[J].Aquaculture research,2009,41(1):61-69.
[48] RODRIGUEZ-ESTRADA U,SATOH S,HAGA Y,et al.Effects of inactivated Enterococcus faecalis and mannan oligosaccharide and their combination on growth,immunity,and disease protection in Rainbow trout[J].North American journal of aquaculture,2013,75(3):416-428.
[49] 肖克宇.水產動物免疫與應用[M].北京:科學出版社,2007.
[50] 馬志紅,晉晶,劉洪梅,等.甘露寡糖對鯉魚生長性能和免疫功能的影響[J].安徽農業科學,2010,38(3):1291-1292,1296.
[51] TORRECILLAS S,MAKOL A,CABALLERO M J,et al.Immune stimulation and improved infection resistance in European sea bass (Dicentrarchus labrax) fed mannan oligosaccharides[J].Fish & shellfish immunology,2007,23(5):969-981.
[52] YOSHIDA T,KRUGER R,INGLIS V.Augmentation of non-specific protection in African catfish,Clarias gariepinus (Burchell),by the long‐term oral administration of immunostimulants[J].Journal of fish diseases,1995,18(2):195-198.
[53] SPRING P,WENK C,DAWSON K A,et al.The effects of dietary mannaoligosaccharides on cecal parameters and the concentrations of enteric bacteria in the ceca of salmonella-challenged broiler chicks[J].Poultry science,2000,79(2):205-211.
