基于SNP 和InDel 標(biāo)記的巴西木薯遺傳多樣性與群體遺傳結(jié)構(gòu)分析
2021-12-17 08:15:36鄒枚伶張辰笈江思容EderJorgedeOliveira張圣奎夏志強(qiáng)王文泉李有志
作物學(xué)報(bào) 2021年1期
關(guān)鍵詞:分析
孫 倩 鄒枚伶 張辰笈,4 江思容,5 Eder Jorge de Oliveira張圣奎 夏志強(qiáng),3,4,* 王文泉,3,4,5,* 李有志
1 廣西大學(xué)生命科學(xué)與技術(shù)學(xué)院 / 亞熱帶農(nóng)業(yè)生物資源保護(hù)與利用國家重點(diǎn)實(shí)驗(yàn)室, 廣西南寧 530004; 2 中國熱帶農(nóng)業(yè)科學(xué)院熱帶生物技術(shù)研究所, 海南海口 571101; 3 中國熱帶農(nóng)業(yè)科學(xué)院熱帶生物組學(xué)大數(shù)據(jù)中心, 海南海口 571101; 4 海南大學(xué), 海南海口570203; 5 南京農(nóng)業(yè)大學(xué), 江蘇南京 210095; 6 Embrapa Mandioca e Fruticultura, Cruz das Almas, Bahia 44380-000, Brazil; 7 齊魯工業(yè)大學(xué), 山東濟(jì)南 250306
木薯(Manihot esculentaCrantz) 是大戟科(Euphorbiaceae)木薯屬(ManihotP. Mill.)的多年生灌木植物, 具有高生物量、抗貧瘠、抗病蟲能力強(qiáng)等特點(diǎn), 被廣泛種植于亞、非、美三洲等多個(gè)國家或地區(qū)[1]。木薯起源于亞馬遜河流域, 于19 世紀(jì)20 年代傳入中國, 最初種植于廣東省, 之后逐漸在海南、廣西、貴州、云南等地大量種植。它是世界三大薯類作物之一, 同時(shí)也是世界上第六大糧食作物, 僅次于小麥、水稻、玉米、馬鈴薯和大麥[2]。其用途廣泛, 除可食用、飼用外, 還可用于生產(chǎn)加工, 如造紙、紡織、生物燃料等[3]。木薯塊根還可用于提取淀粉, 加工成薯?xiàng)l、面包, 以及生產(chǎn)燃料乙醇等; 莖稈可用來進(jìn)行木薯繁殖、粉碎還田、做燃料等; 葉片可作蔬菜食用或喂魚、養(yǎng)蠶等[4]。
木薯基因組具有高度雜合的特性, 原因是其異花授粉, 且長期進(jìn)行無性繁殖。由于基因組的高度雜合, 從而增加了木薯遺傳變異的多樣性, 這些多樣性可為木薯育種人員提供更多可選擇的良好親本,但同時(shí)由于木薯的基因組高度雜合、親緣關(guān)系不清晰、遺傳改良周期長等特點(diǎn)也增加了育種的工作難度[5]。目前已有一些利用相關(guān)序列擴(kuò)增多態(tài)性(sequence-related amplified polymorphism, SRAP)、簡單重復(fù)序列(simple sequence repeat, SSR)、擴(kuò)增片段長度多態(tài)性 (amplified fragment length polymorphism, AFLP)、單核苷酸多態(tài)性(single nucleotide polymorphisms, SNP)等分子標(biāo)記進(jìn)行木薯遺傳多樣性的研究。Fregene 等[6]利用SSR 標(biāo)記對(duì)來源于哥倫比亞、巴西和秘魯?shù)鹊氐哪臼淼胤狡贩N的種質(zhì)資源多樣性評(píng)價(jià)發(fā)現(xiàn), 不同國家來源的木薯種質(zhì)的遺傳多樣性水平都很高, 其中來自巴西和哥倫比亞材料的基因多樣性水平最高。Alex 等[7]利用13 對(duì)SSR
標(biāo)記對(duì)巴西多地的傳統(tǒng)甜木薯品種的群體結(jié)構(gòu)和遺傳多樣性評(píng)估結(jié)果顯示, 該群體的遺傳多樣性平均值為0.5407, 范圍為0.3138 (GA21)~0.6502(GA140), 表明該群體的遺傳變異性寬泛。Carvalho等[8]采用SSR 標(biāo)記和RAPD 標(biāo)記的研究中發(fā)現(xiàn), 巴西的木薯種質(zhì)資源的地理來源和遺傳聚類具有顯著正相關(guān)關(guān)系。
豐富木薯種質(zhì)資源的遺傳多樣性, 并對(duì)其遺傳背景和性狀進(jìn)行綜合評(píng)價(jià), 發(fā)掘控制優(yōu)良性狀的優(yōu)異等位基因, 對(duì)今后木薯育種具有重大意義。全基因組關(guān)聯(lián)分析(genome-wide association study,GWAS)能夠鑒定目的表型性狀與遺傳標(biāo)記或基因間的關(guān)系, 并檢測出控制相關(guān)性狀的優(yōu)良等位基因位點(diǎn)[9]。而進(jìn)行關(guān)聯(lián)分析需要先評(píng)估實(shí)驗(yàn)群體的遺傳多樣性、遺傳結(jié)構(gòu)及親緣關(guān)系[10]。但目前利用 SNP 和 InDel 標(biāo)記對(duì)木薯進(jìn)行遺傳多樣性、親緣關(guān)系及群體結(jié)構(gòu)分析等的相關(guān)研究還鮮為報(bào)道。
本研究擬利用SNP 和InDel 分子標(biāo)記, 對(duì)由巴西Embrapa 機(jī)構(gòu)提供的來源于巴西多地的192 份木薯種質(zhì)資源進(jìn)行遺傳多樣性和群體結(jié)構(gòu)分析。本研究將為以后木薯育種親本選配提供材料和理論指導(dǎo), 也可為下一步通過關(guān)聯(lián)分析發(fā)掘控制木薯種質(zhì)中優(yōu)良性狀的優(yōu)異等位基因提供理論依據(jù),從而促進(jìn)利用分子標(biāo)記輔助選擇技術(shù)培育木薯新品種。
1 材料與方法
1.1 試驗(yàn)材料
供試木薯材料共192 份, 均為巴西栽培種木薯(表1)。
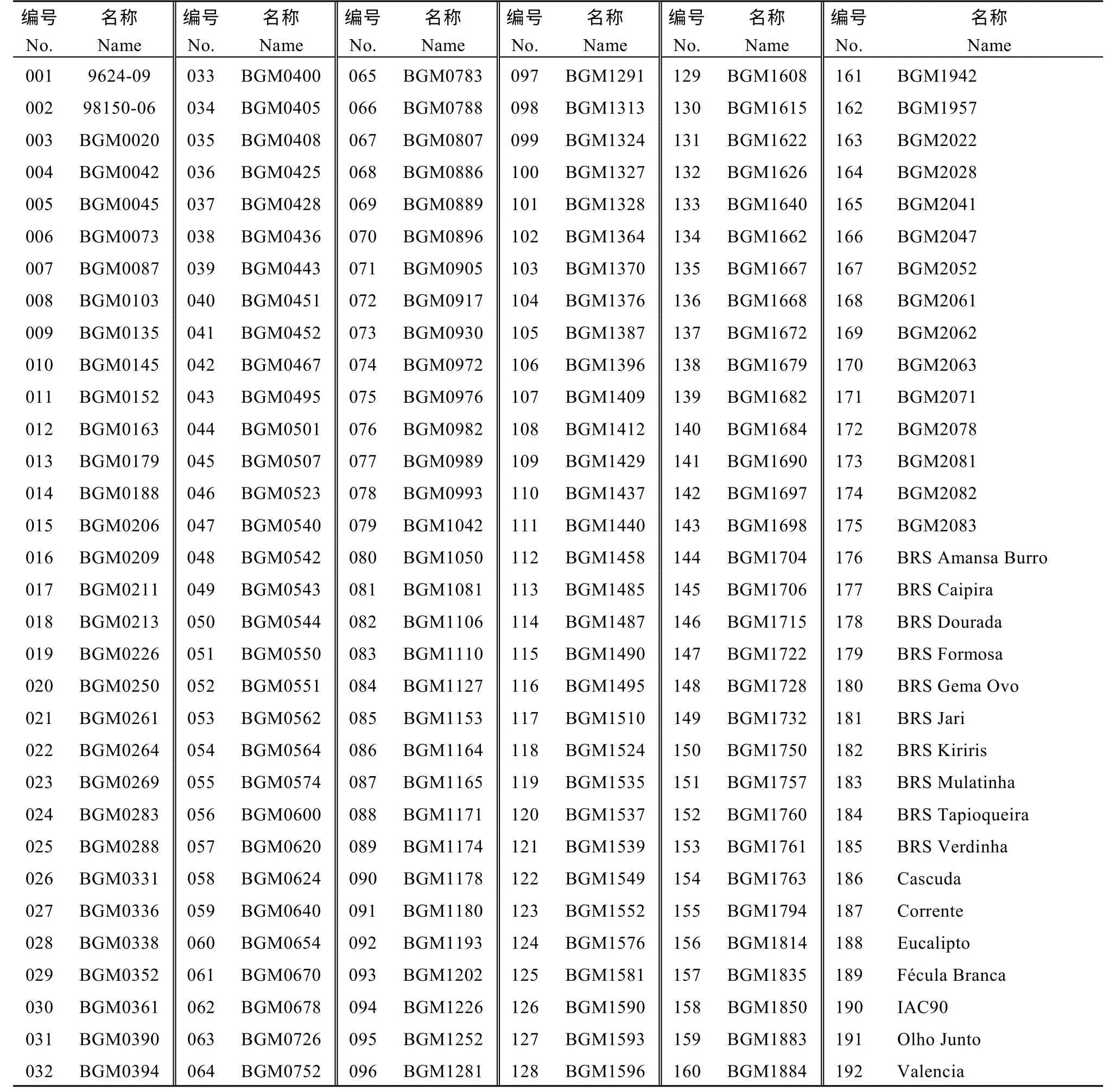
表1 192 份木薯栽培種Table 1 List of 192 cassava cultivars
1.2 木薯基因組DNA 提取及建庫測序
采用改良的CTAB 法[11]提取木薯葉片的基因組DNA, 經(jīng)1%的瓊脂糖凝膠電泳檢測及濃度測定后,將工作液濃度稀釋到100 ng μL-1, -20℃保存。然后利用AFSM[12]技術(shù)對(duì)192 份木薯DNA 樣品使用96孔PCR 板分別構(gòu)建EcoR I-MspI 和EcoR I-HpaII文庫, 單克隆檢測達(dá)到要求后, 將相應(yīng)的EcoR IMspI 和EcoR I-HpaII 文庫按1∶1 比例混合成1 個(gè)文庫, 總共192 個(gè)樣本, 構(gòu)建了2 個(gè)AFSM 文庫, 并利用Hiseq 2500 對(duì)構(gòu)建好的測序文庫進(jìn)行雙端150 bp 測序。
1.3 SNP 和InDels 分子標(biāo)記檢測
利用Perl 腳本(http://afsmseq.sourceforge.net/)對(duì)原始測序數(shù)據(jù)進(jìn)行過濾, 同時(shí)統(tǒng)計(jì)測序得到的總reads 數(shù), 再將reads 根據(jù)AFSM 技術(shù)設(shè)計(jì)的Barcodes分配到每個(gè)個(gè)體中, 并統(tǒng)計(jì)每個(gè)個(gè)體的reads 數(shù)[12]。使用Bowtie 2 軟件[13]將優(yōu)化后的測序reads 比對(duì)到木薯AM560 參考基因組[14], 再使用SAMtools[15]和VCFtools (http://vcftools.sourceforge.net/)檢測 SNP和InDels 位點(diǎn)。基于木薯AM560 參考基因組V6.1,利用snpEff 軟件[16]識(shí)別其變異位置(間隔區(qū)、非翻譯區(qū)/UTR、基因上游區(qū)或基因下游區(qū))、變異類型(同義突變和錯(cuò)義突變、移碼突變和非移碼突變), 同時(shí)對(duì)其進(jìn)行注釋。
1.4 巴西木薯群體結(jié)構(gòu)、遺傳多樣性及群體分化分析
先利用 PHYLIP (http://evolution.genetics.washington.edu/phylip.html)計(jì)算樣品的遺傳距離矩陣,然后用Notepad++軟件將遺傳距離矩陣的文件調(diào)整成合適的格式, 采用鄰接法構(gòu)建系統(tǒng)進(jìn)化樹結(jié)構(gòu),生成tree 文件后, 再使用iTOL (https://itol.embl.de/)繪制進(jìn)化樹圖。通過GCTA 軟件[17]利用檢測出的SNP 對(duì)參試木薯群體材料進(jìn)行主成分分析(PCA)。再使用R 軟件計(jì)算各個(gè)主成分的向量, 繪制PCA散點(diǎn)圖。
此外, 使用ADMIXTURE 軟件[18]進(jìn)行群體結(jié)構(gòu)分析, 估算出最佳群體亞群數(shù)。先用PLINK 軟件[19]調(diào)整ADMIXTURE 軟件的輸入文件格式, 并輸入文件, 然后將亞群數(shù)K值范圍設(shè)置為1~12, 根據(jù)得到的cross-validation error 值選擇合適的亞群數(shù)K值,利用各個(gè)材料占各亞群的遺傳成分系數(shù)(Q)構(gòu)成群體遺傳結(jié)構(gòu)矩陣。
利用VCFtools 軟件(https://vcftools.github.io/index.html)計(jì)算群體遺傳多樣性指數(shù)(π)和群體分化指數(shù)(Fst)[20]。根據(jù)Wright 的研究, 當(dāng)群體分化指數(shù)(Fst)等于0 或1 時(shí), 分別表明亞群間沒有分化, 或亞群間完全分化。而當(dāng)0 < Fst < 0.05、0.05 ≤ FST <0.15、0.15 ≤ FST < 0.25, 或0.25 ≤ FST < 1 時(shí),則分別表明亞群間具有較弱、中等、比較強(qiáng)或非常強(qiáng)的遺傳分化[21]。
2 結(jié)果與分析
2.1 巴西木薯群體基因型分析
通過對(duì)192 份巴西栽培種木薯基因組DNA 進(jìn)行AFSM 建庫及測序, 總共得到了155 G 數(shù)據(jù), 過濾后得到134 G 數(shù)據(jù), 893,020,018 條reads。再利用木薯參考基因組 AM560 V6.1, 通過 SAMtools 和VCFtools 軟件對(duì)192 份木薯樣品基因組進(jìn)行掃描,得到796,006 個(gè)SNPs 和116,821 個(gè)InDels。
通過哈迪溫伯格檢測(HWE)>0.001、次等位基因頻率(MAF)≥0.05 過濾, 并舍去低質(zhì)量的變異位點(diǎn)后, 僅保留了9443 個(gè)高質(zhì)量的變異位點(diǎn)(7946 個(gè)SNPs 和1997 個(gè)InDels)用于后續(xù)分析。其中, 3287個(gè)SNPs 和InDels 位于基因間隔區(qū), 4005 個(gè)SNPs 和InDels 位于基因上游區(qū), 471 個(gè)SNPs 和InDels 位于基因下游區(qū), 2 個(gè)SNPs 和InDels 位于5′端UTR。845個(gè)SNPs 和InDels 屬于錯(cuò)義突變, 745 個(gè)SNPs 和InDels 屬于同義突變, 417 個(gè)SNPs 和InDels 屬于移碼突變, 另有171 個(gè)SNPs 和InDels 屬于其他類型突變(表2)。
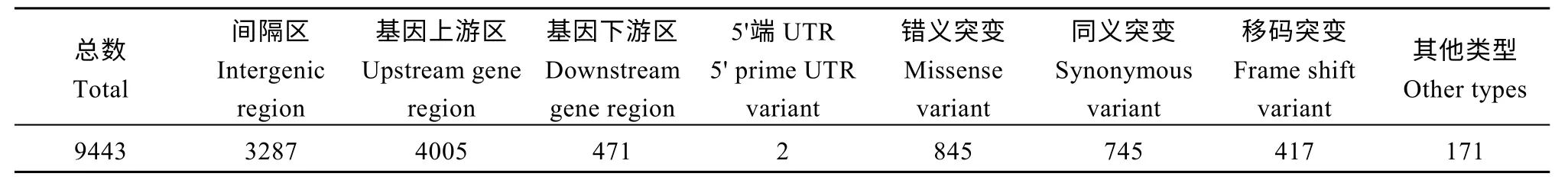
表2 SNPs 和InDels 的統(tǒng)計(jì)Table 2 Summary of SNPs and InDels
2.2 巴西木薯群體結(jié)構(gòu)分析
通過ADMIXTURE 軟件利用9943 個(gè)高質(zhì)量的SNPs 和InDels 分子標(biāo)記對(duì)192 份巴西栽培種木薯進(jìn)行群體遺傳結(jié)構(gòu)分析。將亞群數(shù)K值范圍設(shè)置為1 ~1 2, 計(jì)算不同K值下的交叉驗(yàn)證錯(cuò)誤率(cross-validation error, CV error)。當(dāng)K從1 到2 時(shí), CV error 值迅速減小;K從2 到4 時(shí), CV error 值又逐漸增加; 當(dāng)K從4 到9 時(shí), CV error 值逐漸減小并趨于平緩; 當(dāng)K大于9 時(shí), CV error 值又出現(xiàn)一定的增幅(圖1-a)。說明在K等于9 時(shí), CV error 值最小, 因此巴西栽培種木薯群體可分為 9 個(gè)亞群(Subgroup 1~Subgroup 9)。
192 份巴西木薯可以被分為9 個(gè)亞群, 再根據(jù)每個(gè)個(gè)體在這9 個(gè)亞群的Q 值, 將每個(gè)個(gè)體歸類到Q值最大所在的亞群(圖1-b)。9 個(gè)亞群中分別含有3份、22 份、27 份、6 份、20 份、25 份、24 份、12份和53 份材料。
主成分分析以所有的高質(zhì)量SNPs 和InDels 為基礎(chǔ), 通過R 軟件分析繪圖, 得到如下結(jié)果: 該木薯群體的9 個(gè)亞群在PC1 軸上可以看出一定的分布差距, 大部分亞群可以聚類在一起, 該結(jié)果說明聚類結(jié)果與群體結(jié)構(gòu)的劃分具有一致性(圖1-c)。
由圖2 可知, 聚類結(jié)果與群體結(jié)構(gòu)的劃分相一致, 亞群1、亞群2、亞群4、亞群6 和亞群8 能較好地分別聚在一起, 而其他亞群樣品大致能聚在一起, 且樣品間有一定的交叉。巴西木薯各栽培種之間并未聚類到一起, 可能是由于木薯栽培歷史比較短, 來源于巴西多地的木薯栽培種還未產(chǎn)生明顯的分化。
2.3 巴西栽培種木薯遺傳多樣性分析
利用9943 個(gè)高質(zhì)量的SNPs 和InDels, 通過計(jì)算遺傳多樣性指數(shù)(π), 評(píng)估巴西栽培種木薯群體和各個(gè)亞群的遺傳多樣性。通過vcftools 計(jì)算發(fā)現(xiàn), 巴西栽培種木薯群體的遺傳多樣性指數(shù)為0.274, 亞群1~9 的遺傳多樣性指數(shù)在0.192~0.289 之間, 其中亞群1 具有最低的遺傳多樣性指數(shù)(0.192), 亞群7、亞群2、亞群6 和亞群3 具有相對(duì)較高的遺傳多樣性指數(shù), 分別達(dá)到0.284、0.281、0.264 和0.261, 而亞群5 具有9 個(gè)亞群中最高的遺傳多樣性指數(shù)(0.289)(表3)。說明巴西栽培種木薯群體具有相對(duì)較高的遺傳多樣性水平。
利用群體分化指數(shù)(Fst)評(píng)估巴西栽培木薯亞群間的差異程度(表4)發(fā)現(xiàn), 除亞群1 和亞群4 之間有較強(qiáng)遺傳分化外, 其他亞群之間均為中等或較弱遺傳分化, 亞群間的遺傳分化指數(shù)在0.031~0.152 之間。其中, 亞群8 與其他各亞群之間均為中等分化;而除亞群4 與亞群1 外, 亞群4 和亞群1 分別與其他各亞群之間也均為中等分化。表明, 除亞群1 與亞群4 間遺傳分化較強(qiáng)、親緣關(guān)系較遠(yuǎn)外, 其余各亞群間的為中等或較弱遺傳分化程度, 即亞群間的親緣關(guān)系相對(duì)均較近。
另外, 本研究對(duì)試驗(yàn)所用的巴西栽培木薯的遺傳距離分析發(fā)現(xiàn), 這些木薯種質(zhì)間的遺傳距離為0.084~0.297, 平均遺傳距離為 0.228。其中, BGM 1883 與Valencia 遺傳距離最近(0.084); BGM0640 與BRSJari 遺傳距離最遠(yuǎn)(0.297)。
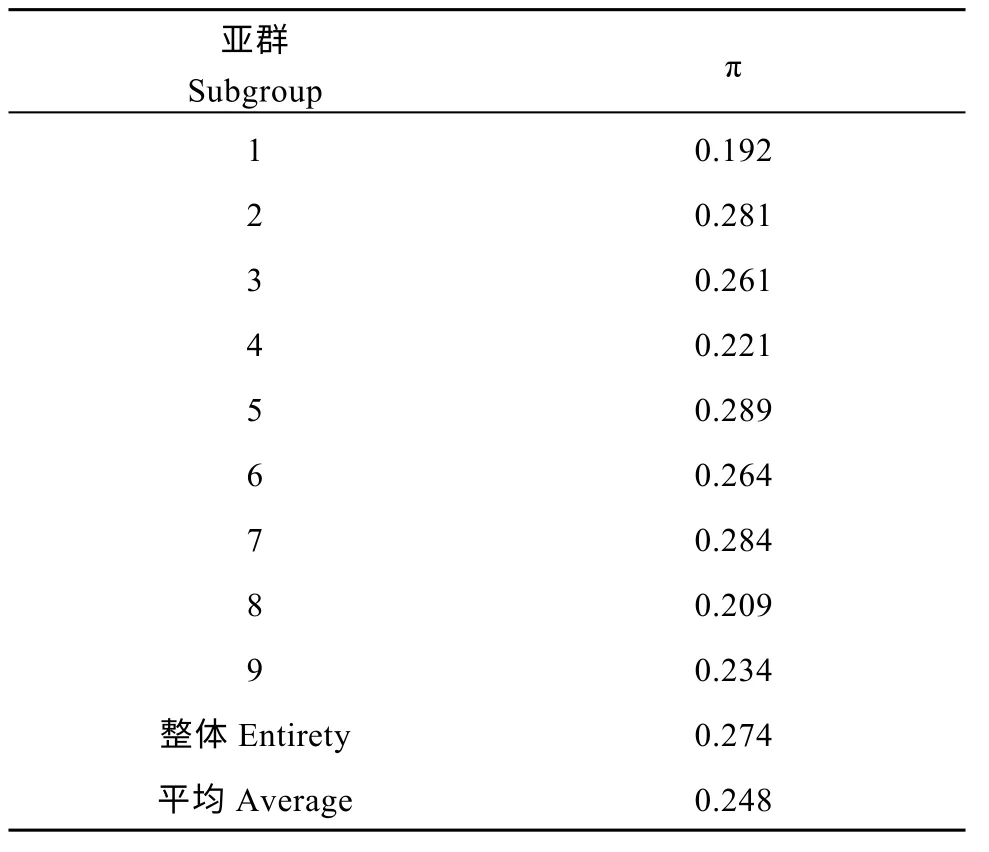
表3 遺傳多樣性指數(shù)(π)的統(tǒng)計(jì)Table 3 Statistics of genetic diversity index (π)
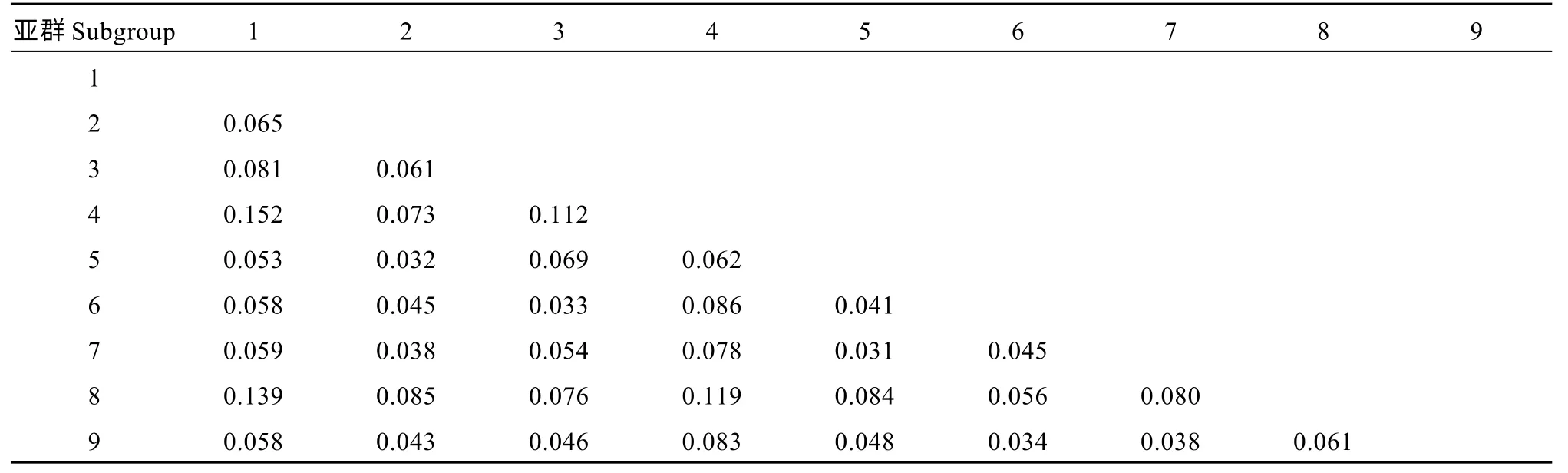
表4 群體分化指數(shù)(Fst)的統(tǒng)計(jì)Table 4 Statistics of population differentiation index (Fst)
3 討論
3.1 木薯群體結(jié)構(gòu)分析
研究表明, 基因型與性狀之間會(huì)產(chǎn)生假關(guān)聯(lián),其原因可能是群體結(jié)構(gòu)分層、等位基因分布不均等[22]。為了消除造成關(guān)聯(lián)分析不準(zhǔn)確的因素, 我們需要先對(duì)試驗(yàn)群體進(jìn)行群體結(jié)構(gòu)分析。本研究利用ADMIXTURE 軟件對(duì)巴西木薯自然群體的群體結(jié)構(gòu)分析表明, 當(dāng)K=9 時(shí), CV error 值最小, 由此將192份木薯種質(zhì)劃分為9 個(gè)亞群, 該結(jié)果與聚類分析、主成分分析的結(jié)果大概相符, 它們之間相互補(bǔ)充及印證, 說明該木薯群體的遺傳結(jié)構(gòu)較為可靠。在這9個(gè)亞群中, 群體分化指數(shù)在0.03~0.15 之間, 且大部分亞群間的群體分化指數(shù)均小于0.09, 表明該木薯群體存在一個(gè)中等偏弱的遺傳分化。前人的研究結(jié)果顯示, 中國熱帶農(nóng)業(yè)科學(xué)院熱帶品種資源種質(zhì)圃收集的158 份木薯種質(zhì)的群體分化指數(shù)在0.03~0.07之間[23]; 在其他地區(qū)的栽培木薯中, 群體分化指數(shù)在0.01~0.05 之間[14], 表明國內(nèi)收集的木薯遺傳分化程度較低。比較看來, 本研究中的巴西木薯種質(zhì)的群體分化指數(shù)高于國內(nèi)收集的木薯種質(zhì), 可挑選優(yōu)質(zhì)巴西木薯品種并引進(jìn)中國, 從而豐富已有的木薯種質(zhì)資源。
3.2 木薯遺傳多樣性分析
本研究對(duì)遺傳多樣性指數(shù)(π)進(jìn)行了計(jì)算, 從而評(píng)估該木薯群體的遺傳多樣性。Ramu 等[14]的研究表明, 來源于不同地區(qū)(尼日利亞、哥倫比亞、巴西等地)的國外栽培木薯的遺傳多樣性指數(shù)為0.0036,低于其祖先(M. esculentassp.flabellifolia, π =0.0051); Fregene 等[6]對(duì)來源于哥倫比亞、巴西和秘魯?shù)鹊氐哪臼淼胤狡贩N的種質(zhì)資源多樣性評(píng)價(jià)發(fā)現(xiàn),巴西和哥倫比亞的木薯種質(zhì)具有最高的遺傳多樣性水平; 在張圣奎對(duì)中國熱帶農(nóng)業(yè)科學(xué)院熱帶品種資源種質(zhì)圃收集的158 份木薯種質(zhì)的遺傳多樣性研究中發(fā)現(xiàn), 該群體的遺傳多樣性指數(shù)為1.21×10-4, 表明該群體的遺傳多樣性較低[23], 同時(shí)也表明目前國內(nèi)的木薯種質(zhì)資源豐富度較為缺乏。本研究發(fā)現(xiàn),該巴西木薯群體中各亞群的遺傳多樣性指數(shù)在0.19~0.29 之間, 平均遺傳多樣性指數(shù)為0.248, 說明巴西木薯群體的遺傳多樣性較為豐富, 可引進(jìn)部分優(yōu)良巴西種質(zhì)以豐富國內(nèi)的木薯種質(zhì)資源。除此之外, 樣本之間的親緣關(guān)系也會(huì)對(duì)關(guān)聯(lián)分析的結(jié)果造成一定的影響。本研究對(duì)192 份木薯種質(zhì)間的遺傳距離進(jìn)行分析, 從而評(píng)估不同材料之間的親緣關(guān)系,結(jié)果發(fā)現(xiàn)這些木薯種質(zhì)的平均遺傳距離為0.228。
4 結(jié)論
本研究利用9943 個(gè)高質(zhì)量的SNPs 和InDels 對(duì)192 份巴西Embrapa 機(jī)構(gòu)提供的木薯種質(zhì)進(jìn)行了和群體遺傳結(jié)構(gòu)分析。遺傳多樣性分析結(jié)果顯示, 巴西木薯群體的遺傳多樣性水平較為豐富, 高于中國和哥倫比亞等地區(qū); 群體遺傳結(jié)構(gòu)分析結(jié)果顯示,該群體被劃分為9 個(gè)亞群, 此結(jié)果與主成分分析及聚類分析結(jié)果基本一致。另外, 該木薯群體的分化程度較低, 但高于國內(nèi)的木薯種質(zhì)資源。遺傳距離分析顯示, BGM1883 與 Valencia 遺傳距離最近,BGM0640 與BRSJari 遺傳距離最遠(yuǎn)。該研究將為之后關(guān)聯(lián)分析挖掘優(yōu)良基因及引進(jìn)優(yōu)良巴西木薯種質(zhì)提供依據(jù)。
猜你喜歡
現(xiàn)代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機(jī)設(shè)計(jì)與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業(yè)技術(shù)(2016年15期)2016-12-01 05:31:22
當(dāng)代經(jīng)濟(jì)研究(2016年5期)2016-12-01 03:12:05
現(xiàn)代農(nóng)業(yè)(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
財(cái)經(jīng)界(學(xué)術(shù)版)(2015年20期)2015-12-23 09:20:13
中國中醫(yī)藥現(xiàn)代遠(yuǎn)程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學(xué)學(xué)報(bào)(社會(huì)科學(xué)版)(2014年3期)2014-04-16 04:38:31
