基于轉(zhuǎn)錄組分析不同溫度下池蝶蚌性別相關(guān)基因的表達(dá)
2021-12-16 11:50:42胡蓓娟徐紅琴洪一江
南昌大學(xué)學(xué)報(bào)(理科版) 2021年5期
關(guān)鍵詞:差異
曾 起,胡蓓娟,徐紅琴,洪一江*
(南昌大學(xué)a.生命科學(xué)學(xué)院;b.江西省水產(chǎn)動(dòng)物資源與利用重點(diǎn)實(shí)驗(yàn)室,南昌 330031)
池蝶蚌是中國(guó)主要的淡水養(yǎng)殖珍珠蚌之一,因育珠能力強(qiáng),產(chǎn)出珍珠具備粒徑大,光澤度好和品相佳等特點(diǎn),養(yǎng)殖規(guī)模迅速擴(kuò)大,成為了我國(guó)重要的經(jīng)濟(jì)貝類,應(yīng)用前景非常巨大[1]。內(nèi)臟團(tuán)作為培育珍珠的重要部位,其產(chǎn)出的珍珠光澤好,顆粒大和形狀圓[2],其中性腺是很重要的因素。
蔡英亞等[3]闡明,產(chǎn)在丹麥的食用牡蠣的性別比例會(huì)隨著水溫變化而發(fā)生改變,當(dāng)生存環(huán)境的水溫高時(shí),其雌性個(gè)體數(shù)量處于優(yōu)勢(shì)地位,反之,雄性個(gè)體數(shù)量占優(yōu)。金啟增等[4]闡述馬氏珠母貝在培養(yǎng)過(guò)程中水溫升高會(huì)導(dǎo)致雌貝的數(shù)量占比增加;反之,雄貝占比增加。據(jù)觀察,水溫與貝類性別的轉(zhuǎn)變有著很重要的關(guān)系,當(dāng)水平處于13.2 ℃~19.9 ℃時(shí),雄性個(gè)體占比高;當(dāng)水溫在20 ℃~29.3 ℃時(shí),雌性個(gè)體和雄性個(gè)體占比幾乎相等;但水溫再次降低時(shí),雄性個(gè)體的數(shù)目又會(huì)重新增加。這表明,水溫低,雄性貝數(shù)量上是占優(yōu)勢(shì)的。Zapata等人[5]研究發(fā)現(xiàn),歐洲牡蠣配子的發(fā)生和性別決定主要受到溫度影響,類固醇不會(huì)作為內(nèi)源性調(diào)節(jié)因子參與性別發(fā)育,而海水溫度的上升和氣候的變暖,可能導(dǎo)致其自然種群中的性別比例傾斜。
最近的研究表明,某些基因會(huì)影響牡蠣或淡水貽貝的性別決定和分化。Teaniniuraitemoana等[6]采集了不同發(fā)育階段的雌性珠母貝和雄性珠母貝樣本,并進(jìn)行了轉(zhuǎn)錄組分析,揭示了其性別決定和性別分化基因,例如影響雄性分化的dmrt和fem1,以及影響雌性發(fā)育的foxl2和vit。Zhou等[7]利用了WCGNA(加權(quán)基因共表達(dá)網(wǎng)絡(luò)分析)方法,篩選了扇貝性別分化和決定的關(guān)鍵基因,并構(gòu)建扇貝性別確定和分化的途徑。在這一途徑中,dmrt1具有主導(dǎo)功能。Wang等[8]報(bào)道foxl2在三角帆蚌的性別分化中起關(guān)鍵作用,而β-catenin[9]在免疫反應(yīng)和性別決定中起關(guān)鍵作用。此外,Hashiyama[10]僅在XX型果蠅性腺的原始生殖細(xì)胞中檢測(cè)到sxl的表達(dá)。
貝類性腺發(fā)育過(guò)程中,發(fā)生了一定程度的性逆轉(zhuǎn)或者雌性同體的出現(xiàn)。例如,一些珠母貝在受到溫度或者其他因素影響時(shí),在生存過(guò)程中表現(xiàn)出連續(xù)性的性別逆轉(zhuǎn)[11];Christelle[12]通過(guò)不同溫度的養(yǎng)殖,發(fā)現(xiàn)長(zhǎng)牡蠣存在性別逆轉(zhuǎn)和雌雄同體現(xiàn)象,并伴隨著性別決定相關(guān)基因表達(dá)的變化;Wu等[13]在研究池蝶蚌發(fā)育過(guò)程中,發(fā)現(xiàn)26~32月齡存在雌性同體現(xiàn)象。因此,為了探究溫度對(duì)其性別分化和和性腺發(fā)育的影響,本研究采用不同養(yǎng)殖溫度下的池蝶蚌為原材料,并采用Pacbio三代測(cè)序技術(shù)和Illumina二代測(cè)序技術(shù)對(duì)雌雄蚌進(jìn)行轉(zhuǎn)錄組測(cè)序,通過(guò)差異基因探究池蝶蚌性腺發(fā)育和性別分化的原因。
1 材料與方法
1.1 實(shí)驗(yàn)材料
池蝶蚌取自于江西省撫州市池蝶蚌良種場(chǎng)。挑選無(wú)插核、健康的29月齡池蝶蚌共50只,放置在充分曝氣的養(yǎng)殖箱進(jìn)行暫養(yǎng)。1周后,各取雌雄蚌5只作為一組,共分5組,分別在15 ℃,20 ℃,25 ℃,30 ℃和33 ℃下進(jìn)行為期2周的養(yǎng)殖試驗(yàn)。養(yǎng)殖結(jié)束后,每個(gè)溫度組各取3只雌蚌和雄蚌性腺混樣,進(jìn)行二代RNA測(cè)序;另取常溫下(23 ℃)暫養(yǎng)的雌雄蚌性腺作為對(duì)照進(jìn)行三代全長(zhǎng)轉(zhuǎn)錄組測(cè)序,性腺樣品液氮速凍,隨后放入-80 ℃冰箱保存?zhèn)溆谩?/p>
1.2 實(shí)驗(yàn)方法
1.2.1 文庫(kù)的構(gòu)建和測(cè)序
總RNA的提取,cDNA文庫(kù)的構(gòu)建,轉(zhuǎn)錄組測(cè)序工作由北京諾禾致源科技股份有限公司完成。二代轉(zhuǎn)錄組測(cè)序平臺(tái)為Illunima Hiseq 2500,三代全長(zhǎng)轉(zhuǎn)錄組測(cè)序平臺(tái)為Pacbio Sequel。
1.2.3 基因表達(dá)水平分析
利用CD-HIT軟件將雌雄蚌轉(zhuǎn)錄本合并去冗余后,得到的轉(zhuǎn)錄本作為參考序列,使用bowtie2軟件將clean data比對(duì)到全長(zhǎng)轉(zhuǎn)錄組,并通過(guò)RSEM軟件獲得單個(gè)樣品中每個(gè)轉(zhuǎn)錄本的reads數(shù)量。考慮到序列深度和基因長(zhǎng)度對(duì)片段的影響,并將所有reads計(jì)數(shù)標(biāo)準(zhǔn)化為FPKM,以將FPKM大于0.3的相對(duì)水平定義為顯著表達(dá)基因,并用其做基因表達(dá)水平統(tǒng)計(jì)。FPKM計(jì)算公式如下:
注:Xi:單個(gè)轉(zhuǎn)錄本的讀取數(shù);Li:轉(zhuǎn)錄本長(zhǎng)度;N:轉(zhuǎn)錄本的讀取總數(shù)。
1.2.4 基因差異表達(dá)分析和聚類分析
實(shí)驗(yàn)采用了DEseq2 R包分析有重復(fù)組的數(shù)據(jù),對(duì)各個(gè)溫度下雌雄之間進(jìn)行差異表達(dá)分析。其中將差異倍數(shù)(FoldChange)大于2及調(diào)整后的p值(padj)小于0.05作為標(biāo)準(zhǔn),篩選符合條件的基因作為差異表達(dá)基因。FoldChange是兩個(gè)樣品間某一轉(zhuǎn)錄本表達(dá)量的比值,padj是通過(guò)對(duì)p值進(jìn)行校正后得到的以不同實(shí)驗(yàn)條件下的差異基因的FPKM值為表達(dá)水平,做層次聚類分析。
1.2.5 差異表達(dá)基因功能注釋
使用NT,NR,Swiss-Prot,KEGG和GO數(shù)據(jù)庫(kù)對(duì)差異轉(zhuǎn)錄本進(jìn)行注釋。
1.2.6 差異基因功能富集分析
差異表達(dá)基因的功能富集分析主要包括GO富集分析和KEGG通路富集分析,其中GO富集分析和KEGG富集分析是通過(guò)clusterProfiler R包完成。
1.2.7 qRT-PCR定量驗(yàn)證
為了驗(yàn)證轉(zhuǎn)錄組數(shù)據(jù),選擇了6個(gè)差異表達(dá)基因的mRNA進(jìn)行qRT-PCR。qRT-PCR引物設(shè)計(jì)使用Oligo7.0工具完成,引物序列由北京擎科生物公司合成,具體引物見表1。β-actin作為內(nèi)參基因,采用2-ΔΔCt相對(duì)定量法定量,每個(gè)樣本每個(gè)基因進(jìn)行3次重復(fù)實(shí)驗(yàn)。

表1 qRT-PCR選用基因及引物序列
2 結(jié)果
2.1 測(cè)序結(jié)果與序列分析
PacBio單分子熒光測(cè)序?yàn)榄h(huán)狀測(cè)序,測(cè)序過(guò)程的單分子產(chǎn)出的有效片段為Subreads,經(jīng)過(guò)自我矯正和去冗余后,獲得雌雄全長(zhǎng)轉(zhuǎn)錄本數(shù)目分別為96 867和111 520條,平均長(zhǎng)度分別為4 095和3 876 bp,主要分布在2.5~3.5 kbp之間(圖1)。
2.2 轉(zhuǎn)錄本的表達(dá)定量
將clean data的reads比對(duì)到全長(zhǎng)轉(zhuǎn)錄本上發(fā)現(xiàn),雌性性腺轉(zhuǎn)錄本比對(duì)上的序列在88%左右,雄性性腺轉(zhuǎn)錄本比對(duì)上的序列在75%左右,雌性轉(zhuǎn)錄本比對(duì)上數(shù)目多于雄性轉(zhuǎn)錄本。用FPKM統(tǒng)計(jì)不同表達(dá)水平下基因的數(shù)量以及單個(gè)基因的表達(dá)水平,發(fā)現(xiàn)在所有轉(zhuǎn)錄組中,F(xiàn)PKM分布最多的在0~1之間,占了50%左右,其中FPKM>5的大約占25%圖2(A)。為了直觀的比較各樣本整體的表達(dá)水平,以及對(duì)每個(gè)處理溫度組表達(dá)水平的離散程度,采用箱線圖展示FPKM的分布顯示10個(gè)組樣本表達(dá)水平整體上一致,其中雌性25 ℃處理組樣本表達(dá)水平略低,雌性30 ℃處理組表達(dá)水平略高圖2(B)。
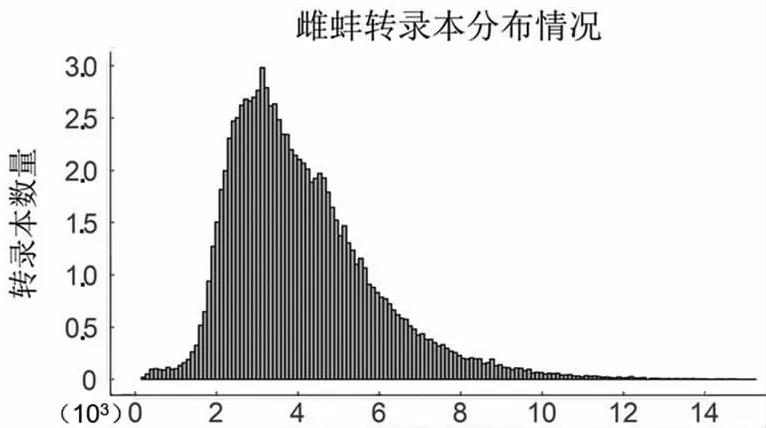
轉(zhuǎn)錄本長(zhǎng)度
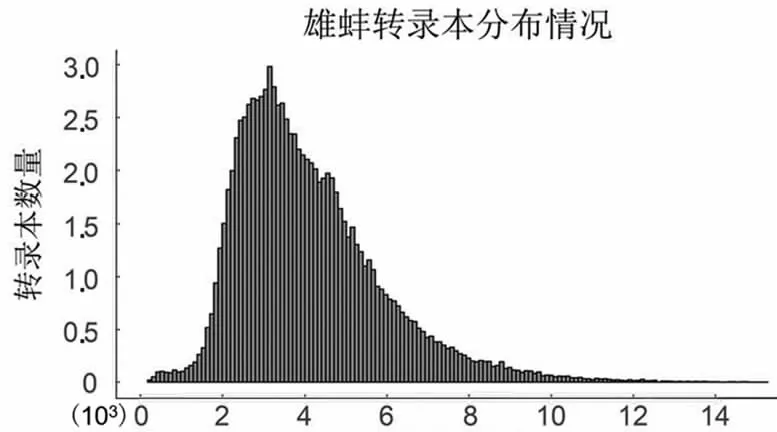
轉(zhuǎn)錄本長(zhǎng)度圖1 轉(zhuǎn)錄本長(zhǎng)度分布圖
2.3 差異表達(dá)基因統(tǒng)計(jì)
采用DEseq2分析鑒定5個(gè)溫度下不同性別處理組間的差異表達(dá)基因,在15 ℃雌雄比較組至33 ℃雌雄比較組中,分別檢測(cè)到6 311,3 013,6 673,6 798,5 685個(gè)差異表達(dá)基因(表2),結(jié)果發(fā)現(xiàn),雌雄之間的差異基因非常豐富。在20 ℃和33 ℃時(shí)差異基因數(shù)量比其它組更少,15 ℃時(shí)雌雄對(duì)比組含3 013個(gè)差異表達(dá)基因,其中1 045個(gè)基因上調(diào),1 968個(gè)基因下調(diào);33 ℃對(duì)比組含5 703個(gè)差異表達(dá)基因,2426個(gè)上調(diào),3277下調(diào)。在5個(gè)處理組中,共有17696條轉(zhuǎn)錄本在所有組間表達(dá)。用層次聚類分析5個(gè)溫度脅迫處理組間差異表達(dá)基因的分布情況發(fā)現(xiàn),雌蚌組與雄蚌組具有類似的表達(dá)情況,其中15 ℃和20 ℃聚為一支,25 ℃,30 ℃和33 ℃聚為一支(圖3)。
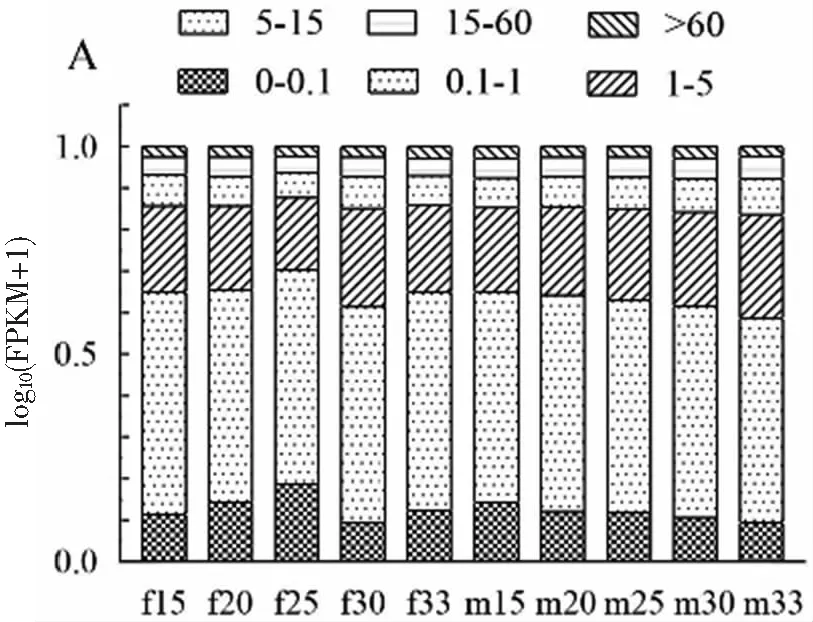
(A) 不同表達(dá)水平區(qū)間的基因數(shù)量統(tǒng)計(jì)圖

(B) 基因表達(dá)水平對(duì)比圖圖2
2.4 不同溫度下差異基因的KEGG富集
差異基因的富集分析是通過(guò)超幾何分布,其富集分析以通路為單位,將轉(zhuǎn)錄本中的基因作為背景,在差異表達(dá)基因中顯著性富集的通路。在所有富集中,均展現(xiàn)富集程度最高的前20個(gè)通路。對(duì)5個(gè)不同溫度下雌雄間所鑒定到的6311,3013,6673,6798和5685個(gè)差異表達(dá)基因進(jìn)行KEGG注釋,分別完成4585,2203,4896,4932和4069個(gè)。其中在15 ℃組,腎素分泌(ko04924),甘氨酸、絲氨酸和蘇氨酸代謝(ko00260),淀粉和蔗糖代謝(ko00500)顯著富集等;在20 ℃組中,顯著富集的有卵母細(xì)胞減數(shù)分裂(ko04114),MAPK信號(hào)通路(ko04016),醛固酮合成與分泌(ko04925)和GnRH信號(hào)傳導(dǎo)途徑(ko04912)等,腎素分泌(ko04924)及磷脂酰肌醇信號(hào)系統(tǒng)(ko04070)也被顯著富集;在25 ℃情況下,主要富集到卵母細(xì)胞減數(shù)分裂(ko04114),細(xì)胞周期(ko04110)和Notch信號(hào)通路和腎素分泌(ko04924)等;在30 ℃的時(shí),富集到細(xì)胞周期(ko04110),細(xì)胞周期-酵母(ko04111)和孕激素介導(dǎo)的卵母細(xì)胞成熟(ko04914)。33 ℃時(shí),可以看出顯著富集的包括丙酮酸代謝(ko00620),腎素分泌(ko04924),胰高血糖素信號(hào)通路(ko04922)。KEGG富集顯示在各溫度的處理組均含有卵母細(xì)胞分裂,但是在20 ℃和25 ℃的時(shí)呈現(xiàn)出顯著富集(qvalue<0.05);在30 ℃出現(xiàn)了孕激素介導(dǎo)的卵母細(xì)胞成熟;在25 ℃的時(shí)候開始具有細(xì)胞周期。

表2 相同溫度下雌雄性腺之間的差異基因結(jié)果

注:聚類分析圖,將log10(FPKM+1)值進(jìn)行歸一化轉(zhuǎn)換并進(jìn)行聚類,深色表示高表達(dá),淺色表示低表達(dá)。顏色從深到淺,表示log10(FPKM+1)從大到小。
2.5 差異基因中性別相關(guān)基因的篩選和分析
對(duì)5個(gè)溫度處理組之間的差異表達(dá)基因的進(jìn)行組間比較,維恩圖有802個(gè)差異表達(dá)基因在5個(gè)對(duì)比組中重疊(見圖5)。其中共上調(diào)的基因?yàn)?19個(gè)數(shù),共下調(diào)的基因個(gè)數(shù)為483個(gè)。將5個(gè)組共表達(dá)差異的差異基因進(jìn)行GO富集分析發(fā)現(xiàn),下調(diào)基因的GO富集顯示在幾丁質(zhì)的結(jié)合,幾丁質(zhì)代謝過(guò)程和蛋白結(jié)合,鈣離子的結(jié)合,細(xì)胞外區(qū)域,膜,胞外空間上,這些均與與珍珠質(zhì)的分泌和形成有關(guān);上調(diào)基因的GO富集顯示在蛋白質(zhì)結(jié)合,ATP的結(jié)合和酶的活動(dòng)等,這些與精子的生成和活動(dòng)有關(guān)(見圖6)。
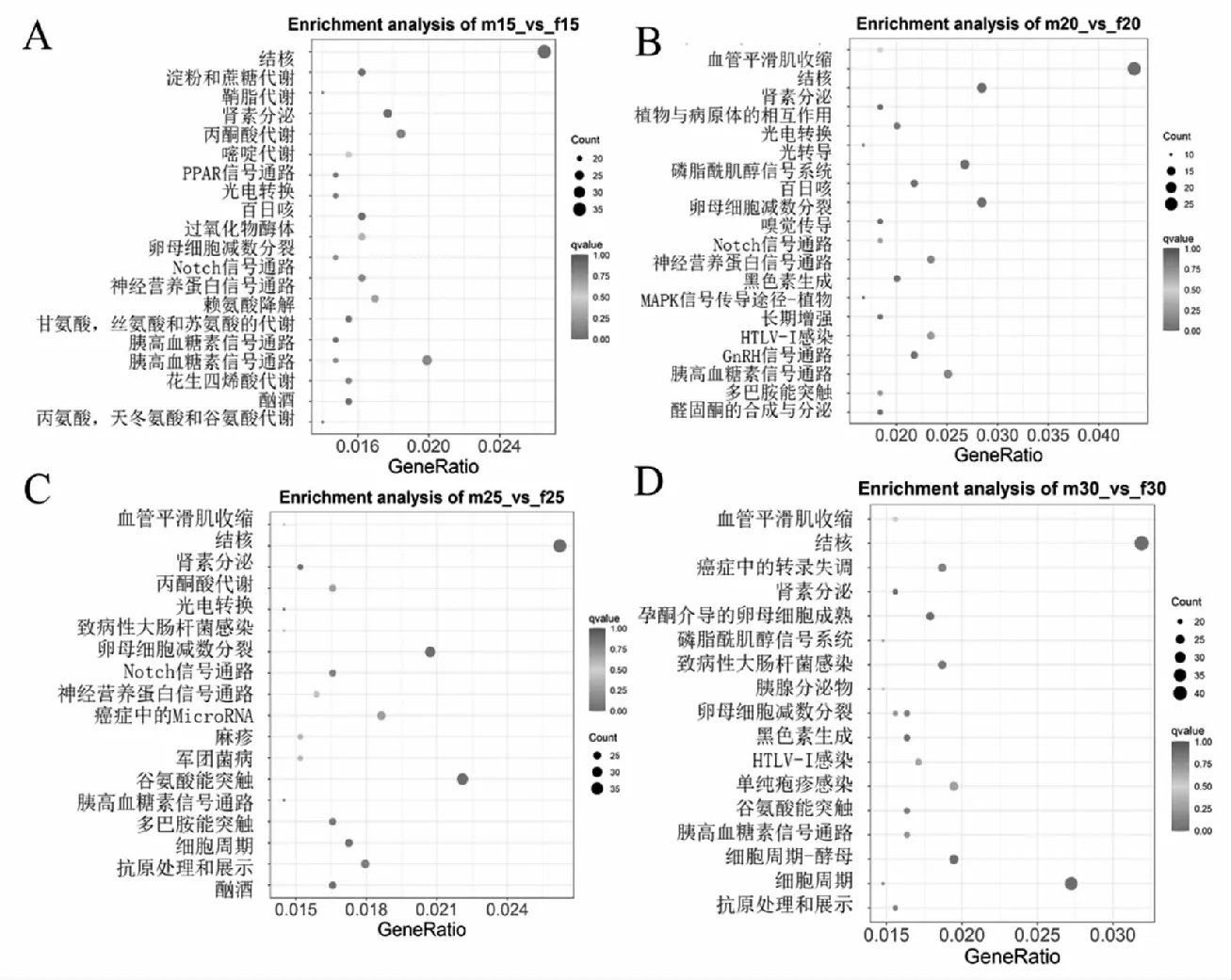
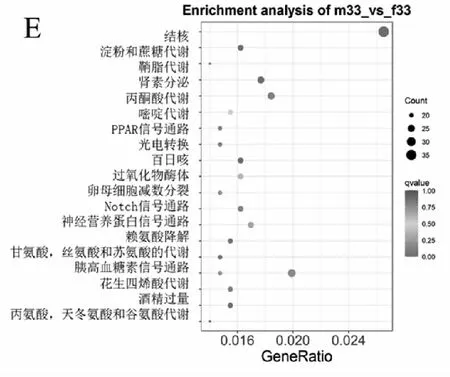
注:A:15 ℃;B:20 ℃;C:25 ℃;D:30 ℃;E:33 ℃。圖4 各個(gè)溫度下差異基因KEGG富集結(jié)果
2.6 差異基因中性別相關(guān)基因的篩選和分析
在差異表達(dá)基因中篩選出大量卵巢發(fā)生相關(guān)基因,其中包括中蛋黃鐵蛋白(FRIY),卵黃囊蛋白(SIAE),核黃素結(jié)合蛋白(RBP),卵黃囊衍生的胚胎大分子(ELYS)等,還有卵細(xì)胞發(fā)生相關(guān)基因,包括卵母細(xì)胞表達(dá)核仁蛋白(ANO39),卵母細(xì)胞成熟因子(MOS),卵母細(xì)胞特異性母體影響因子(ZAR1)和卵黃原蛋白(VIT)等。
在差異表達(dá)基因中鑒別到大量精子發(fā)生相關(guān)基因,精子發(fā)生相關(guān)蛋白4,7,17,22(spat4,spat7,spt17,spt22),精子發(fā)生和中心粒相關(guān)蛋白1(sper1)和精子特異性接頭組蛋白H1樣蛋白(HILS1);精子運(yùn)動(dòng)相關(guān)蛋白精子鞭毛蛋白1、2(spef1,spef2),精子表面蛋白17(sp17)等;還發(fā)現(xiàn)大量精巢相關(guān)基因,例如,睪丸特異性絲氨酸/蘇氨酸蛋白激酶2(TSSK2),T復(fù)合物睪丸表達(dá)蛋白1(Tcte-1)和精巢表達(dá)蛋白10,11,30(Tex10,Tex11,Tex30)等。
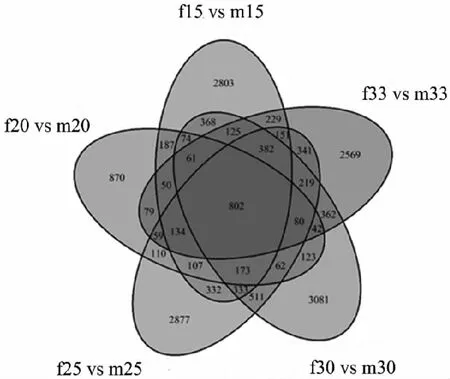
注:m15 vs f15:在15 ℃下雄雌間差異基因的數(shù)量,其他類似。
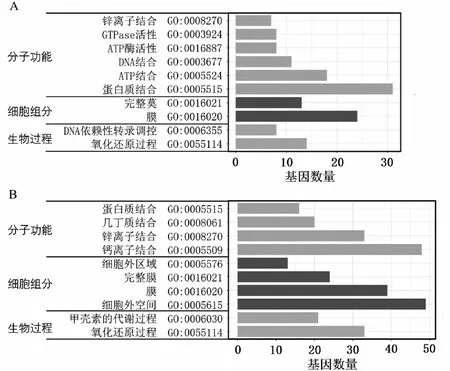
注:A:上調(diào)差異表達(dá)基因的GO富集;B:下調(diào)差異表達(dá)基因的GO富集。
為了進(jìn)一步探究性別決定和性別分化,從各個(gè)組間中挑選出差異基因,列出出部分性別決定和性別分化相關(guān)基因如表4.7,其中sox2,wnt4,fem1,rspo1,vit在5個(gè)溫度對(duì)比組均顯著表達(dá)差異,但是sox5,sox9和folx2在20 ℃時(shí)沒有表達(dá)差異。其中tra1,wnt4,fem1,rspo1,sox9和folx2等,文獻(xiàn)記載它們與性別決定和性別分化有關(guān)。
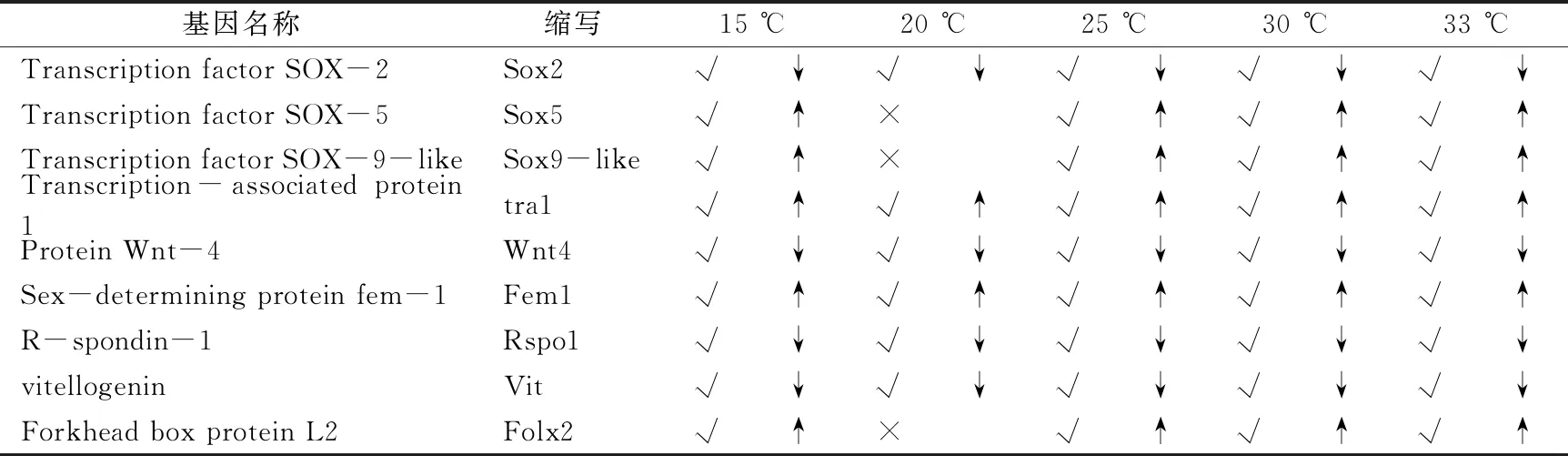
表3 差異基因中性別決定基因的篩選
2.7 qRT-PCR驗(yàn)證分析
隨機(jī)挑選了6個(gè)差異表達(dá)基因pif,tex43,wnt4,rsop1,tra1和fem1,進(jìn)行qRT-PCR驗(yàn)證。其中wnt4,rspo1,pif,calm在雌性中顯著表達(dá)(P<0.05,log2FC<-1),tra1和fem1在雄性中顯著表達(dá)(P<0.05,log2FC>1)。通過(guò)qRT-PCT進(jìn)行驗(yàn)證,篩選的6個(gè)基因雌雄間表達(dá)差異極顯著(P<0.01),且差異表達(dá)基因與轉(zhuǎn)錄組測(cè)序結(jié)果中的表達(dá)量變化倍數(shù)趨勢(shì)基本一致(圖7)。結(jié)果表明,轉(zhuǎn)錄組結(jié)果可靠,利用RNA-Seq技術(shù)對(duì)池蝶蚌雌雄株進(jìn)行轉(zhuǎn)錄組測(cè)序以及篩選與性別相關(guān)的差異基因是可行的。根據(jù)熒光定量結(jié)果,fem1,tra1,rspo1和wnt4的mRNA表達(dá)水平如圖8,在雌蚌中fem1在15 ℃和20 ℃時(shí)高于其他水平;在雌蚌和雄蚌中rspo1和tra1在15 ℃和20 ℃時(shí)均低于其他溫度水平;在雌蚌中wnt4在25 ℃時(shí)表達(dá)量最高。
3 討論
SMRT測(cè)序技術(shù)可以提升無(wú)參轉(zhuǎn)錄組的組裝效果[14]。在這項(xiàng)研究中,我們首次采用RNA-seq和SMRT測(cè)序結(jié)合的方式,生成了池蝶蚌全長(zhǎng)轉(zhuǎn)錄組。經(jīng)分析后獲得了96 867和111 520個(gè)高質(zhì)量的全長(zhǎng)序列。在這項(xiàng)研究中,我們首次采用RNA-seq和SMRT測(cè)序結(jié)合的方式,生成了池蝶蚌全長(zhǎng)轉(zhuǎn)錄組。經(jīng)分析后獲得了96867和111520個(gè)高質(zhì)量的全長(zhǎng)序列,為無(wú)需PCR的基因結(jié)構(gòu)探索和遺傳功能探究提供了寶貴數(shù)據(jù)。與之前使用Illumina平臺(tái)的池蝶蚌轉(zhuǎn)錄組研究相比[15],二代的雌雄轉(zhuǎn)錄本的平均長(zhǎng)度分別為484和472 bp,而本實(shí)驗(yàn)的轉(zhuǎn)錄本的長(zhǎng)達(dá)4 095和3 876 bp。研究調(diào)查了5個(gè)不同溫度下池蝶蚌雌雄性腺的基因表達(dá)。對(duì)差異表達(dá)基因研究發(fā)現(xiàn),在20 ℃和33 ℃的時(shí)候,差異基因數(shù)量最少,推測(cè)這兩個(gè)溫度下雌雄表達(dá)差異比其他組小。各雌雄對(duì)比組只有802個(gè)基因在各個(gè)溫度下以不同方式共同表達(dá)。KEGG富集分析發(fā)現(xiàn),25 ℃時(shí)出現(xiàn)細(xì)胞周期通路,說(shuō)明細(xì)胞活動(dòng)相對(duì)開始活躍;同時(shí)雌性個(gè)體生殖相關(guān)的活動(dòng)變得活躍起來(lái),在25 ℃時(shí)出現(xiàn)卵母細(xì)胞減速分裂通路和notch信號(hào)通路等。KEGG表明25 ℃時(shí),雌性相關(guān)活動(dòng)更加活躍,有利于雌性生長(zhǎng)發(fā)育。

注:Log2FC:雄雌之間的差異倍數(shù)。圖7 qRT-PCR定量結(jié)果
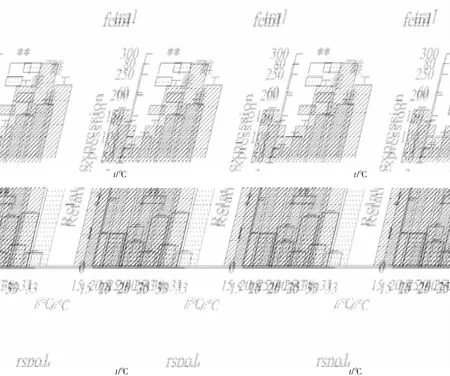
注:fem1,tra1基因在15 ℃下雌蚌性腺中的表達(dá)量分別定義1;rspo1,wnt4基因在15 ℃下雄蚌性腺中的表達(dá)量分別定義1;**表示P<0.01。
轉(zhuǎn)錄組分析結(jié)果表明,wnt4,rspo1,fem1和tra1等4個(gè)性別決定或性別分化基因在不同溫度下的池蝶蚌性腺中始終出現(xiàn)差異。wnt4和rsop1的表達(dá)量雌性高于雄性,fem1和tra1的表達(dá)量雄性高于雌性。wnt4是性別決定基因,且在雌雄哺乳動(dòng)物的形態(tài)發(fā)育中起到關(guān)鍵的作用[16]。小鼠性別決定前的性腺中wnt4存在表達(dá),性別分化后在卵巢中仍持續(xù)表達(dá),但精巢中的表達(dá)顯著下降[17]。李海龍等人[18]研究發(fā)現(xiàn)wnt4在櫛孔扇貝的性腺發(fā)育的各個(gè)時(shí)期均存在表達(dá),尤其成熟性腺表達(dá)最高,推測(cè)其在性腺生殖細(xì)胞成熟的整個(gè)過(guò)程中起著不可忽視的作用。Rspo1在雌性性別決定通路上游發(fā)揮作用,研究發(fā)現(xiàn)能啟動(dòng)wnt/β-catenin通路,對(duì)雌性卵巢的分化是不可或缺的,另外,rspo1能促進(jìn)卵巢發(fā)育并抑制精巢發(fā)育[19]。在青鳉中研究發(fā)現(xiàn)rspo1基因激活的通路,對(duì)雌性卵巢的分化是不可或缺的[20],rspo1在青鳉雄性胚胎中過(guò)表達(dá)能夠引起完全的性逆轉(zhuǎn)(雄轉(zhuǎn)雌),證明rspo1通路在青鳉雌性卵巢發(fā)育中發(fā)揮著關(guān)鍵的作用。在線蟲中,fem1是雄性個(gè)體發(fā)育,以及雄性和雌雄同體精子發(fā)生所必須的[21],它可以通過(guò)阻止tra1的作用,從促進(jìn)雄性發(fā)育產(chǎn)生。Doniach等[22]發(fā)現(xiàn)在櫛孔扇貝中,fem1只在成熟的雄性個(gè)體內(nèi)表達(dá),而不是雌性或者雌雄同體內(nèi),表明fem1在維持雄性化可能發(fā)揮作用。周祖陽(yáng)等[23]在長(zhǎng)牡蠣中成功克隆出fem1,并對(duì)長(zhǎng)牡蠣各個(gè)發(fā)育時(shí)期的fem1進(jìn)行表達(dá)分析,推測(cè)其參與了性別決定和性別分化的過(guò)程。fem1,rspo1和wnt4的qRT-PCR與轉(zhuǎn)錄測(cè)序的表達(dá)量變化倍數(shù)一致,且rspo1和wnt4在雌性中的表達(dá)水平高于雄性,而fem1在雄性中的表達(dá)水平高于雌性。推測(cè)fem1,rspo1和wnt4在性別調(diào)控中發(fā)揮作用,rspo1和wnt4在維持雌性化發(fā)生作用,而fem1在維持雄性化發(fā)揮作用。
Berkseth等[24]表明tra1在發(fā)育過(guò)程中通過(guò)參與阻止與雄性發(fā)育的基因來(lái)促進(jìn)雌性發(fā)育方面有普遍作用。Ronald[25]總結(jié)出tra1可以被視為是雌性動(dòng)物中的一種阻遏物,它可以關(guān)閉雄性相關(guān)基因然后發(fā)育成雌性。在此次研究中,通過(guò)差異基因分析發(fā)現(xiàn),tra1在雄性中顯著表達(dá),而且熒光定量結(jié)果分析也顯示tra1表達(dá)量在雄性中高于雌性。Wu等[13]發(fā)現(xiàn)池蝶蚌在26~32月齡時(shí),出現(xiàn)雌雄同體的現(xiàn)象,且大部分的雌性同體蚌從雄蚌中發(fā)育而來(lái)。此次研究的池蝶蚌處于30月齡,推測(cè)tra1的出現(xiàn),可能關(guān)閉池蝶蚌雄性相關(guān)基因,進(jìn)而產(chǎn)生雌雄同體的現(xiàn)象。熒光定量結(jié)果顯示,25 ℃以上時(shí),tra1的表達(dá)量高于10 ℃和15 ℃水平,溫度的升高,可能進(jìn)一步促進(jìn)池蝶蚌雌雄同體的形成。
Holleley等[26]發(fā)現(xiàn)當(dāng)孵化溫度在22 ℃~32 ℃之間時(shí),鬃獅蜥由性染色體決定性別;當(dāng)溫度超過(guò)32 ℃時(shí),后代中雌性的比例越來(lái)越高。Weber C和Zhou等[27]在巴西紅耳龜中發(fā)現(xiàn),早期的原始性腺在31 ℃時(shí)分化為卵巢,在26 ℃時(shí)分化為精巢。在具有溫度依賴性性別決定的紅耳龜中,rspo1的上調(diào)發(fā)生在溫度敏感期,即性腺的發(fā)育受到某個(gè)特定溫度的影響。因此,rspo1表達(dá)是對(duì)溫度敏感的,并且在處于胚胎從雌性轉(zhuǎn)變?yōu)樾坌缘姆趸瘻囟葧r(shí),rspo1基因就會(huì)下調(diào)[28]。熒光定量結(jié)果表明,rspo1在15 ℃和20 ℃時(shí),雌蚌和雄蚌中表達(dá)量較25 ℃,30 ℃和33 ℃下顯著(P<0.01)降低,而推測(cè)在15 ℃和20 ℃溫度下有利于雄性的發(fā)育;雌蚌中的wnt4在25 ℃時(shí)表達(dá)量最高,而在雄蚌中的wnt4隨著溫度升高而升高;雌蚌中的fem1在25 ℃之后表達(dá)量出現(xiàn)顯著(P<0.01)降低,而雄蚌中的fem1受溫度影響不大。
綜上,根據(jù)KEGG富集分析和rspo1,wnt4,tra1和fem1的表達(dá)情況判斷,推測(cè)25 ℃以上的溫度,適合雌性的生殖發(fā)育。本研究為探索池蝶蚌的性別分化和性腺發(fā)育做出一定的參考,關(guān)于溫度對(duì)池蝶蚌性腺發(fā)育和性別分化的影響,還涉及諸多基因與其產(chǎn)物之間的相互作用,依然有待進(jìn)一步的深入發(fā)掘和探索。
猜你喜歡
英語(yǔ)世界(2023年10期)2023-11-17 09:19:16
汽車實(shí)用技術(shù)(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級(jí)語(yǔ)數(shù)英綜合(2019年8期)2019-08-27 02:23:00
中國(guó)特種設(shè)備安全(2018年11期)2019-01-08 02:08:32
小學(xué)科學(xué)(學(xué)生版)(2018年7期)2018-08-13 09:33:04
中國(guó)非營(yíng)利評(píng)論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現(xiàn)代語(yǔ)文(2016年21期)2016-05-25 13:13:50
