玉米淀粉調控的研究進展
2021-12-13 10:35:28李潛龍康云海張從合
中國農學通報 2021年33期
杜 明,方 玉,2,李潛龍,康云海,王 慧,2,張從合,2
(1上海中科荃銀分子育種技術有限公司,上海 200030;2安徽荃銀高科種業(yè)股份有限公司,合肥 230088)
0 引言
玉米(Zea mays L.)是中國第一大糧食作物,同時也是重要的工業(yè)原料和飼料原料的主要來源[1]。近年來,隨著國民生產總值以及人民生活水平質量的提高,玉米的產量已經不是生產中的限制因素,相反,玉米的品質成為現(xiàn)代玉米育種工作者的重點。因此改善玉米的品質性狀對于玉米產業(yè)以及國民經濟的發(fā)展具有重要的意義。淀粉作為和谷類作物種子的主要儲存形式,玉米胚乳中貯藏的淀粉占整個世界淀粉市場的90%以上[2],研究表明,玉米籽粒的增重過程主要是淀粉合成和積累的過程,通過不斷改良玉米淀粉的含量從而可以實現(xiàn)“以質增重”[3]。
長期以來,玉米科研工作者通過分子生物學方法和轉錄組測序手段發(fā)現(xiàn)了大量調節(jié)玉米淀粉合成和代謝的轉錄因子,因其對植物的生理生化反應以及生長有重要的作用而被廣泛研究[4],其中很多調控因子的功能近年來也陸續(xù)得到證明。如抑制玉米ZmZHOUP1的表達可以導致籽粒發(fā)育的異常[5];SUSIBA2,屬于WRKY轉錄因子家族,能夠與淀粉合成基因的啟動子結合,從而參與淀粉的合成[6];有研究表明淀粉合成關鍵酶可以和其他蛋白相互作用從而以蛋白異聚體的形式調控淀粉的合成[7],但是,關于玉米淀粉在蛋白水平上的研究較少。玉米中,ISA1和非催化功能ISA2形成異聚復合物或者僅含有ISA1的同聚復合物來參與淀粉的合成[8]。本研究列出了相關酶,進一步澄清了淀粉合成代謝的調控網(wǎng)絡,調控淀粉代謝的轉錄因子和順式作用元件的系統(tǒng)表征為玉米品質的遺傳改良提供依據(jù)。
1 玉米淀粉結構
淀粉根據(jù)其連接方式可以分為直鏈淀粉和支鏈淀粉,直鏈淀粉和支鏈淀粉的組成、比值決定著玉米籽粒的品質[9],以D-吡喃葡萄糖為單位連接形成。直鏈淀粉是由α-1,4-糖苷鍵連接而成的,相對分子質量大約為3萬~16萬,由300~1200個葡萄糖殘基構成。直鏈淀粉在其分子的一端為非還原末端基,另一端為還原末端基,聚合度大約為100~6000。直鏈淀粉的通過其內部的氫鍵相互作用,使其以卷曲螺旋的形式存在,每圈包括6個葡萄糖單元[10]。當?shù)庖号c直鏈淀粉相接觸時,碘分子進入淀粉分子內部的螺旋內部,平均每圈螺旋可以束縛一個碘分子,直鏈淀粉可以束縛很多的碘分子,因此直鏈淀粉的與碘顯色呈現(xiàn)深藍色[11]。支鏈淀粉的組成既含有α-1,4-糖苷鍵,也含有α-1,6-糖苷鍵,相對分子質量要大于直鏈淀粉,分子質量大約為10萬~100萬,α-1,6-糖苷鍵連接的分支上有許多形如樹枝狀的分支[12,13]。支鏈淀粉分子具有一個還原末端基和許多非還原末端基,聚合度大約為1000~300萬。支鏈淀粉雖然也可以形成螺旋卷曲,但是其每個分枝較短,與碘分子形成絡合物的數(shù)目較少,因此,支鏈淀粉與碘的顯色呈現(xiàn)紫紅色或紅色[14]。
2 玉米淀粉的生物合成
淀粉合成的場所為葉綠體和淀粉體,其生物合成需要一系列酶(AGPase、SS、SBE、DBE)的參與和調控。
白天臨時性淀粉在葉片中合成積累,夜間進行降解供植株進行生理代謝,其顆粒淀粉幾乎全部為支鏈淀粉,體積較小[15-16]。在葉綠體中,通過卡爾文循環(huán)固定的CO2,經過一系列的化學反應最終形成1-磷酸葡萄糖,隨后在ADP-葡萄糖焦磷酸化酶的作用下形成ADPG,ADPG是淀粉合成的前體物質,在淀粉合成酶和淀粉分支酶的作用下合成直連淀粉和支鏈淀粉[17-18]。
在淀粉體中,以蔗糖作為合成淀粉的碳源,經過水解后的蔗糖以磷酸葡萄糖的形式進入淀粉體參與淀粉的合成。目前,有超過30個基因參與玉米淀粉的合成[19],且編碼這些關鍵酶的主效基因和功能已經比較明確。ADP-葡萄糖焦磷酸化酶作為一種速效酶,是由兩個大小相似的異源四聚體組成[20]。AGPase突變體中,其活性下降了90%~95%,且淀粉含量只有野生型玉米的25%~30%[21-22]。淀粉合成酶(SS)是一種葡萄糖轉移酶,主要包括游離態(tài)的淀粉粒合成酶(SSS)和束縛態(tài)淀粉粒合成酶(GBSS)[23],其中,GBSSⅠ與淀粉顆粒緊密地結合,GBSSⅠ與直連淀粉的合成相關,GBSSⅡ主要存在于非貯藏組織中,與支鏈淀粉的合成有關[23]。SSⅣ對支鏈淀粉結構作用較小,對淀粉粒的形成及顆粒形態(tài)起作用[25]。研究表明ZmEREB156可以和ZmSSⅢa的啟動子結合,利用糖和ABA的協(xié)同作用正向調控淀粉的生物合成[26]。玉米中,wx突變體中則不含直連淀粉[27]。淀粉分支酶(SBE)具有雙重的作用,一方面可以切開α-1,4糖苷鍵連接的葡聚糖,另一方面又能把切開的短鏈通過α-1,6糖苷鍵連接于受體鏈上。其中,SBEⅡa和SBEⅡb氨基酸序列的同源性達到80%[28],在SBE突變體中,在SBEⅡb失去其活性的前提下SBEⅡa才會發(fā)生支鏈淀粉結構的改變,sbe1或者SBEⅡa其中一個發(fā)生突變并不會改變胚乳淀粉的鏈長分布[29]。淀粉去分支酶(SDBE)能消除支鏈淀粉中出現(xiàn)的異常支鏈分支,從而形成正常的分支[30]。在玉米突變體ZmPUL中,其在白天合成和在夜間分解能量的速率是低于野生型的,進一步表明PUL與生物代謝和淀粉的合成有關[31]。
3 淀粉合成相關基因在玉米各個組織中的表達
目前,通過轉錄組測序手段,研究者們已經篩選并且解析了大量調控玉米淀粉合成的相關基因[32]。利用TBtools軟件對近年來發(fā)表的一些基因構建熱圖分析,大部分的基因在玉米胚乳中有較高的表達或者只在玉米胚乳中表達,少數(shù)在葉片中表達,這與淀粉的合成場所剛好相對應。
4 相關轉錄因子家族基因在淀粉合成中的作用
4.1 DOF家族轉錄因子
對于玉米DOF家族中,DOF蛋白是植物特有的一類轉錄因子,含有鋅指結構。據(jù)報道,DOF蛋白在植物的發(fā)育過程中可以作為轉錄激活劑或者抑制劑,可以結合AAAG順勢作用元件[33-35]。在玉米ZmDOF36中,ZmDOF36可以與ZmAGPL1、ZmAGPS1a、ZmSSⅡa、ZmGBSSⅠ、ZmISA1以及ZmPUL中的AAAG序列特異性結合,這6個基因參與了淀粉的合成[36]。ZmDOF1可以與C4磷酸烯醇丙酮酸鹽基因的啟動子相互作用,從而增強啟動子的活性。ZmDOF2是一種抑制劑抑制轉錄[37]。ZmDOF3可以識別和結合NKd1啟動子中的AAAG序列,對正向調節(jié)胚乳中淀粉的合成[38]。
4.2 NAC家族轉錄因子
NAC轉錄因子作為植物特有的一類轉錄因子,在其N端包括A、B、C、D、E 5個結構域,是NAC轉錄因子的結合區(qū)域,由160個氨基酸殘基組成[39]。ZmNAC34是淀粉重要的調節(jié)劑,ZmNAC34可以識別結合下游基因啟動子的CATGTG序列,同時其過表達可以降低籽粒中的總淀粉含量,而總淀粉含量增加[40]。ZmNAC128和ZmNAC130可以通過一個常見的DNA結合位點ACGCAA直接激活Bt2,從而影響B(tài)t2的轉錄,從而降低其蛋白含量,最終導致淀粉合成受阻[41]。ZmNAC36可以增加5個淀粉合成基因的轉錄水平[42]。
4.3 MYB家族轉錄因子
MYB類轉錄因子廣泛存在于真核生物中,在其氨基酸末端存在高度保守的DNA結合結構域。根據(jù)保守結構域中串聯(lián)重復序列的數(shù)目分為四類:1R-、R2R3-、3R-和4R-MYB[43]。據(jù)報道,R2R3-MYB蛋白參與植物組織發(fā)育等多種生物學功能。ZmMYB14是典型的R2R3-MYB類轉錄因子,ZmBT1屬于線粒體載體蛋白家族(MCF)大類[44],在胚乳中特異性表達,在-176 bp到-168 bp處含有MBSI位點(TAACTG),ZmMYB14可以與TAACTG序列結合,從而增強ZmGBSSⅠ、ZmSSⅠ、ZmSSⅡa、ZmSBE1、ZmISA1和ZmISA2啟動子的活性,同時,ZmMYB14是玉米ZmBT1的關鍵調節(jié)因子,與淀粉的生物合成密切相關[45-46]。
4.4 bZIP家族轉錄因子
bZIP家族蛋白可以分為A、B、C、D、E、F、G、H、I、S共10個亞族[47]。蛋白可以識別的ACGT序列,包括G-box(CACGTG)、C-box(GACGTC)和A-box(TACGA)序列。bZIP家族轉錄因子ZmbZIP91可以直接與淀粉合成基因ZmAGPs1、ZmSS1、ZmISA1啟動子中的ACTCAT序列結合,進而調節(jié)葉片中淀粉的合成[48]。ZmbZIP22可以與淀粉合成基因ZmAGPL1、ZmAGPS1a、ZmGBSSⅠa、ZmSSⅠa、ZmSSⅢa、ZmISA1、ZmISA2、ZmBEⅡa啟動子中的ACGT序列結合,同時,ZmbZIP22基因的過表達導致玉米籽粒中的總淀粉、直連淀粉和支鏈淀粉的含量下降,ZmbZIP22負向調控淀粉的合成[49]。近年來研究較多的Opaque-2也是屬于bZIP家族,研究表明,玉米突變體中O2可以結合ZmSSⅢ啟動子中的O2-box以調節(jié)淀粉合成,PBF可以在ZmSSⅢ啟動子的O2-box附近結合P-box的核心元件并與O2共同調解淀粉合成基因的表達,同時PBF和O2可以間接調節(jié)ZmSSⅡa和ZmSBE1,最終導致淀粉含量的下降[49]。
4.5 MADS-BOX家族轉錄因子
在植物中,MADS-BOX有兩種類型,ZmMADS1a作為一種典型的MADS-BOX家族轉錄因子,屬于MIKCc二類。在過表達玉米中,通過對16個調節(jié)玉米淀粉合成與代謝的基因的表達量分析,結果表明ZmMADS1a基因廣泛參與玉米胚乳中淀粉和葡萄糖代謝的調控,對淀粉含量進行分析,總淀粉、直連淀粉和支鏈淀粉的含量都是顯著上升的[51]。異源過表達水稻中ZmES22可以與OsGIF1的啟動子序列CAGGT序列結合,對淀粉合成通路中17個基因表達量分析,除了OsISA2、OsSSⅠ和OsPUL,大多數(shù)基因的表達量都是下調,對其淀粉含量進行分析,發(fā)現(xiàn)其總淀粉和直連淀粉的含量顯著下降,ZmES22負向調控水稻胚乳中淀粉的合成[52]。
4.6 AP2/EREB家族轉錄因子
AP2/EREB蛋白至少包含一個DNA結合結構域,AP2/EREB分為3個獨立的家族:ERF、AP2、RAV。過表達玉米中,ZmEREB156可以增強Zmsh2和ZmSSⅢa的啟動子活性,同時ZmEREb156可以與ZmSSⅢa的啟動子結合,不能與Zmsh2的啟動子結合。此外,ZmEREB156通過和ABA的協(xié)同效應,正向調節(jié)淀粉生物合成基因ZmSSⅢa[26]。ZmEREB94有較強的轉錄活性,可以直接與ZmSSⅠ的啟動子結合,同時也可以間接調節(jié)Zmsh2和ZmGBSSⅠ的表達[53]。
4.7 其他
OPAQUE11作為一種bHLH型轉錄因子,可以直接調控胚乳發(fā)育和營養(yǎng)代謝,同時,也直接調節(jié)cyPPDK(胞質型丙酮酸磷酸雙激酶)和多種碳水化合物的代謝,在突變體中對其淀粉含量進行分析,發(fā)現(xiàn)其淀粉含量顯著下降[54]。
據(jù)報道,在24個玉米淀粉合成基因中,有15個基因主要在胚乳中表達。研究表明,對玉米胚乳采用ABA處理后發(fā)現(xiàn)其可以抑制淀粉基因的表達,特別是ZmAGPS2和ZmSSⅢa;IAA同樣可以抑制淀粉基因的表達,ZmSSⅢa除外;而蔗糖可以上調15個在玉米胚乳中表達的淀粉基因。通過蔗糖和ABA結合誘導處理胚乳,發(fā)現(xiàn)大多數(shù)淀粉基因的表達水平是對照的2倍;而蔗糖和IAA結合誘導中,雖然表達水平受到抑制,但是大多數(shù)淀粉基因的表達都是高于對照組[55]。
5 結論
通過對不同家族的轉錄因子的研究,發(fā)現(xiàn)大量的轉錄因子都是通過與淀粉通路中的合成基因的啟動子結合或者是影響啟動子的活性來調節(jié)淀粉的生物合成(圖2),轉錄因子也可以通過影響一些基因的表達來間接影響淀粉的含量(圖3)。
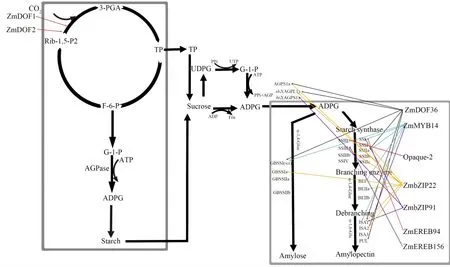
圖2 葉片和胚乳中淀粉代謝的調節(jié)因子
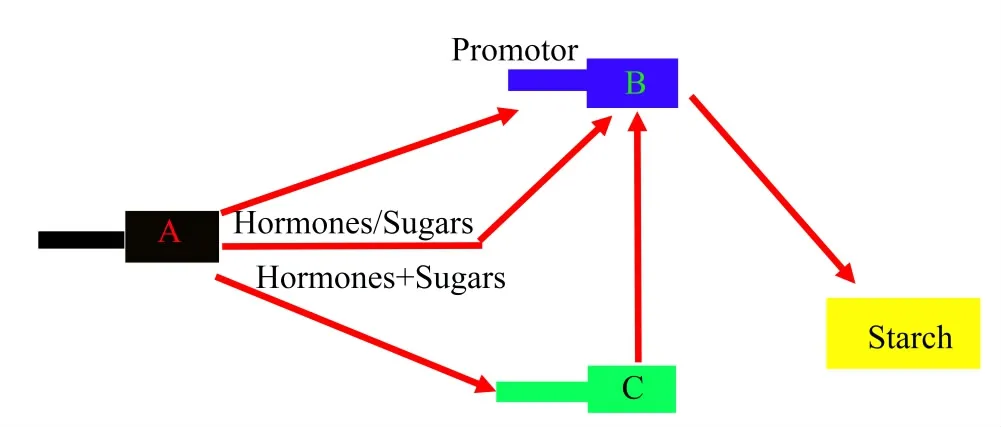
圖3 植物淀粉調節(jié)網(wǎng)絡
6 問題與展望
近年來,研究者通過各種手段和方法得到了大量與淀粉合成直接或者間接相關的基因。第一,利用轉錄組分析和GWAS挖掘,通過篩選解析獲得大量調控玉米淀粉合成的相關基因,第二,通過已知發(fā)表的其他物種(如水稻、擬南芥)的基因來尋找玉米中的同源基因,進一步驗證其是否參與淀粉的合成與代謝。第三,利用EMS誘變技術獲得大量突變體庫,通過觀察籽粒大小相關的突變體或者篩選胚乳相關的突變體得到起作用的基因。第四,利用酵母雙雜技術來篩選與淀粉酶互作的基因,通過進一步的驗證進一步獲得與淀粉合成代謝相關的基因。目前,這些手段雖然比較成熟,同時,得到的淀粉合成相關基因大量為轉錄因子,雖然這些轉錄因子的功能被后續(xù)證明,但是這些轉錄因子之間是否存在一定的聯(lián)系上不清楚。此外,研究者雖然發(fā)現(xiàn)淀粉合成關鍵酶多和其他蛋白相互作用從而以蛋白異聚體的形式來調控淀粉的合成,但是對于淀粉合成在蛋白水平上的調控機制尚不清楚,與淀粉合成關鍵酶的直接互作或間接互作調控蛋白還知之甚少。
隨著分子生物學和基因工程的發(fā)展,推測玉米淀粉的發(fā)展方向(1)通過篩選胚乳中特異性表達或者高表達的基因,通過突變其中的基因來研究其在淀粉合成中的功能;(2)既然淀粉合成通路中的相關酶的作用已經得到驗證,因此可以通過共表達模式分析,找到與其表達模式一致的基因,然后進一步研究其功能;(3)目前對蛋白水平上的研究較少,因此可以通過酵母文庫篩選與淀粉合成相關酶互作的基因研究其功能,此項工作應該是未來玉米淀粉研究的重點。
