甜菜應答鹽脅迫PUB基因的生物信息學分析
2021-12-13 10:35:52李海英
中國農學通報 2021年33期
王 爽,李海英
(1黑龍江大學農業微生物技術教育部工程研究中心,哈爾濱 150500;2黑龍江大學生命科學學院/黑龍江省普通高校分子生物學重點實驗室,哈爾濱 150080)
0 引言
蛋白質作為生命的物質基礎,需要保證其正常合成以及及時的降解。蛋白質穩態的保持與泛素蛋白酶體途徑(ubiquitn proteasome system,UPS)密切相關[1]。在UPS中,由3種泛素化酶介導的級聯反應對靶蛋白進行修飾。這個系統從泛素分子開始,首先E1泛素激活酶激活泛素分子并轉移至E2泛素藕合酶上,E2負責將泛素分子通過E3泛素連接酶轉移至靶蛋白上,完成泛素化修飾過程[2]。通常來說,特定的E2與E3的配對確定泛素化的類型,從而賦予底物不同的命運。然而在此過程中E3起到特異性識別底物的作用,由于其功能的重要性使E3成為研究熱點。
植物中大部分U-box型蛋白(plant U-box protein,PUB)具有E3泛素連接酶活性,在UPS中扮演著重要的角色。據報道,在酵母和人類基因組中分別鑒定出2個和21個U-box型蛋白,而在擬南芥、水稻及大豆基因組中分別鑒定出64個、77個、125個U-box型蛋白[3-4]。植物中U-box型增加的數量表明這些蛋白可能在調節植物生長和發育過程中發揮作用。U-box型蛋白具有高度保守的約70個氨基酸序列,形成β-β-α-β折疊結構,暴露出促進E2與泛素分子結合的芳香族氨基酸殘基和疏水氨基酸殘基[5]。U-box蛋白被認為是修飾的RING蛋白,但與RING蛋白不同,U-box蛋白通過鹽橋或氫鍵傳遞泛素分子。除了含有U-box結構域,PUB蛋白往往包含其他結構域,例如ARM基序、激酶結構域、WD40結構域、MIF4G基序等,基于這一點,可以對PUB蛋白進行再次分類[6-7]。
鹽脅迫作為影響植物生長發育的重要的環境脅迫,可造成細胞內外Na+/K+離子不平衡、光合速率降低、細胞損傷甚至死亡[8]。研究表明,U-box基因家族作為重要的E3泛素連接酶家族,在植物應答非生物脅迫過程扮演著重要角色。擬南芥PUB18和PUB19轉錄水平可受鹽脅迫誘導,與野生型植株相比,pub18pub19雙突變株對鹽脅迫的抗性增加[9]。擬南芥PUB22和PUB23可以泛素化RPN12a,從而在干旱脅迫反應中起作用[10]。小麥中TaPUB1可以與甘露糖苷酶蛋白(TAMP)相互作用,在植物響應鹽脅迫反應過程中起到正向調控作用[11]。
甜菜作為中國重要的糖料作物,具有較強的耐鹽能力。目前還沒有關于甜菜PUB蛋白的相關報道。本實驗利用前期得到的鹽脅迫下甜菜轉錄組數據,從中篩選得到42個編碼包含U-box結構域蛋白的差異表達基因。本研究對這些差異表達基因進行生物信息學分析,為深入研究甜菜PUB基因功能奠定基礎。
1 材料與方法
1.1 材料來源
二倍體栽培甜菜T710-Mu品系。
1.2 甜菜PUB蛋白的確定
實驗室前期利用280 mmol/L NaCl[12]處理甜菜T710-Mu品系,得到鹽處理條件下的甜菜T710-Mu品系葉片轉錄組數據[13],從中篩選差異表達的PUB基因,得到42個差異表達的甜菜PUB基因,根據蛋白在轉錄組數據中的出現順序將甜菜PUB蛋白命名為BvPUB1-BvPUB42,進行接下來的生物信息學分析。
1.3 甜菜PUB蛋白理化性質檢測
該研究通過在PROTPARAM網站(https://web.expasy.org/protparam/)中輸入PUB蛋白質的氨基酸序列,選擇計算參數,分別對PUB蛋白質的氨基酸大小、分子量及等電點進行預測及分析。
1.4 基因在染色體上的分布
利用NCBI數據庫獲取甜菜PUB基因在染色體上的位置,并使用TBtools繪制染色體定位圖。
1.5 PUB基因及保守結構域分析
利用NCBI數據庫和TAIR擬南芥數據庫(https://www.arabidopsis.org/)分別獲得甜菜PUB蛋白質的氨基酸序列和擬南芥同源蛋白質的氨基酸序列,利用Pfam網站(http://pfam.xfam.org/)以及MEME在線軟件(https://meme-suite.org/)對甜菜PUB蛋白的結構域進行分析,下載結果。在NCBI上查找甜菜(Beta vulgaris L.)的GFF3文件,使用MEGA-X軟件并選用鄰位相連法構建系統發育樹。使用TBtools本地軟件對PUB進行基因結構分析,并將基因結構和保守結構域可視化。
1.6 甜菜PUB蛋白的網絡互作預測
利用STRING網站(https://string-db.org/)預測甜菜PUB蛋白的互作蛋白,繪制PPI,利用Cytoscape軟件進行美化。
2 結果與分析
2.1 PUB基因的鑒定與分析
對在鹽脅迫條件下差異表達的42個PUB基因序列進行分析(表1)。結果表明,它們編碼的蛋白質長度介于245~1034 aa之間,預測分子量介于27885.88~96138.82 Da之間,等電點介于5.02~8.59之間。

表1 甜菜PUB基因的詳細信息
2.2 染色體定位分析
利用基因位置信息以及Tbtools軟件繪制甜菜PUB基因的染色體定位圖(圖2),通過定位圖可以看出,42個甜菜PUB基因在9條染色體上均有定位,其中和chr1和chr2上包含較多的PUB基因,chr3、chr4和chr8上只包含1個PUB基因。
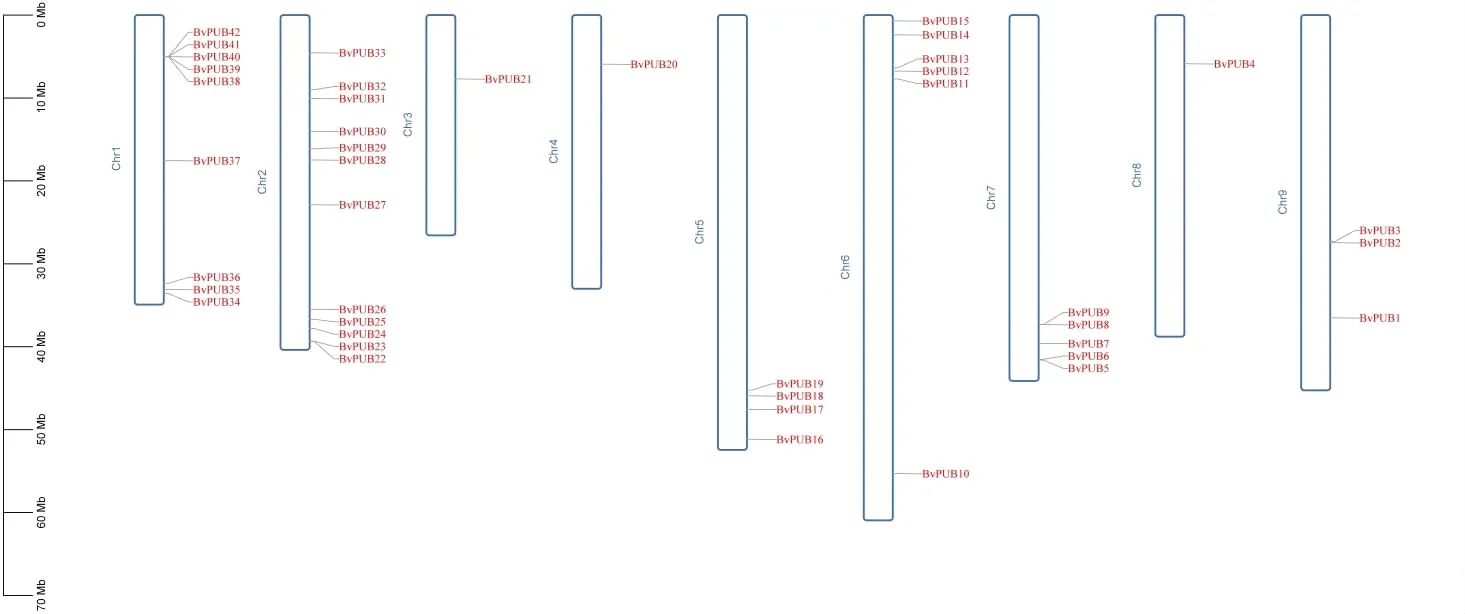
圖1 甜菜PUB基因染色體定位示意圖
2.3 PUB基因序列分析及保守結構域分析
利用甜菜PUB基因序列構建系統進化樹,并結合甜菜基因組注釋文件對甜菜PUB基因序列進行分析,繪制保守基序示意圖(圖2b)和基因結構示意圖(圖2c)。結果顯示,PUB基因包含多個保守的motif,保守的motif構成的保守的結構域對于賦予PUB蛋白不同的生物學功能具有重要作用。這些保守的結構域主要包括U-box結構域,Pkinase結構域、Pkinase-Tyr結構域、Usp結構域、Arm結構域以及KAP結構域。基因序列分析結果顯示,不同的PUB基因之間包含內含子與外顯子個數存在差異。

圖2 PUB基因結構分析
2.4 序列比對及分類
根據蛋白質的結構特征和系統進化樹比較,將擬南芥中PUB蛋白分為7類。以擬南芥PUB蛋白分類為依據,對甜菜PUB蛋白進行分類分析,構建系統發育進化樹(圖3)。結果顯示,甜菜PUB蛋白位于ClassⅡ、ClassⅢ、ClassⅣ類型中,說明這42個甜菜PUB蛋白與擬南芥的ClassⅡ、ClassⅢ、ClassⅣ類型PUB蛋白結構更相近。這為進一步研究甜菜PUB蛋白家族提供一定基礎。利用MEME在線網站對甜菜PUB蛋白的保守基序進行分析(圖4),結果顯示甜菜PUB蛋白包含保守的約70個氨基酸組成的U-box型結構域(圖5)。
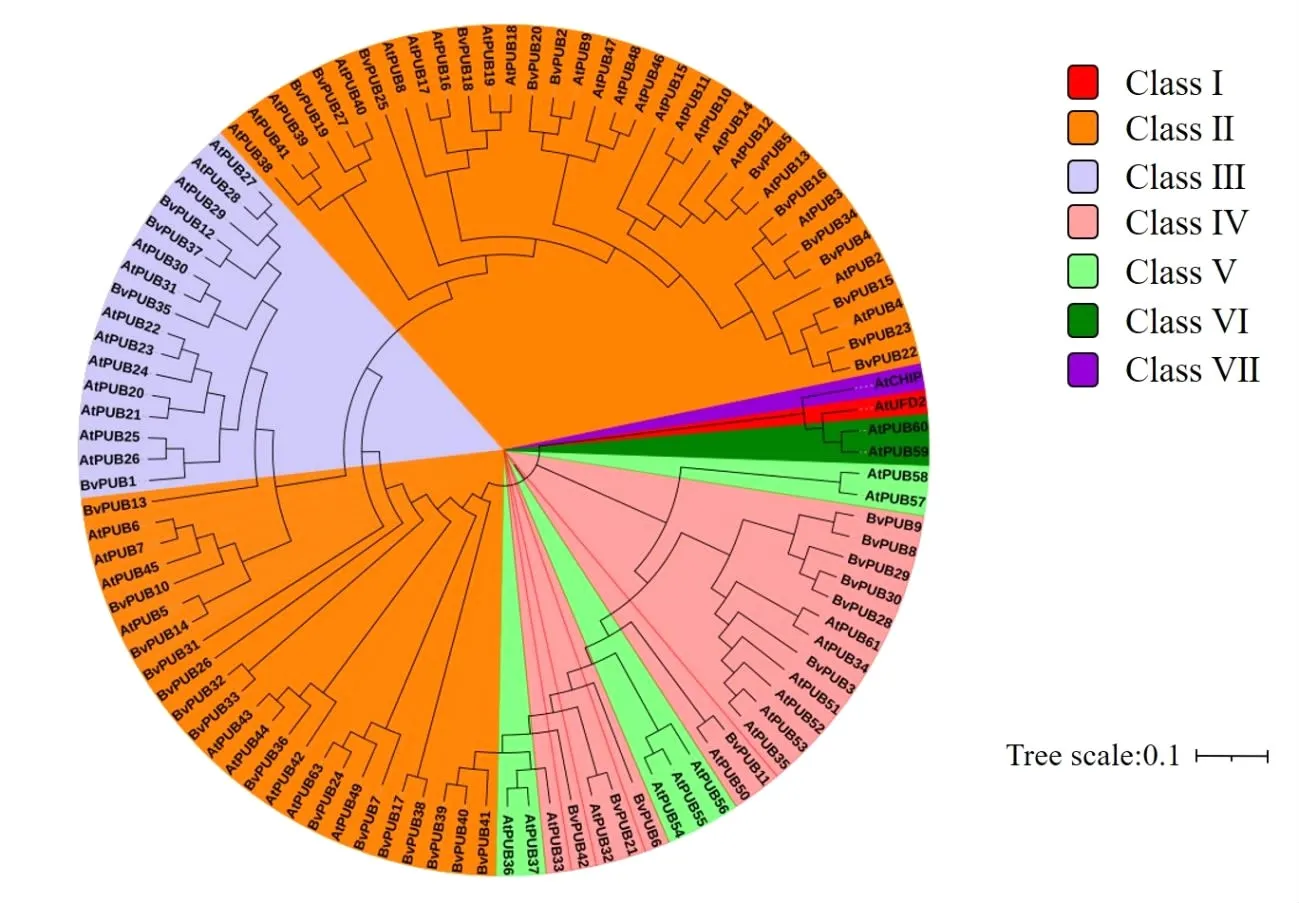
圖3 系統發育進化樹

圖4 PUB蛋白保守基序示意圖

圖5 U-box保守結構域示意圖
2.5 甜菜PUB蛋白互作網絡
利用STRING網站,將甜菜PUB蛋白與擬南芥蛋白進行比對并繪制PPI。結果顯示,BvPUB5、BvPUB18、BvPUB 34、BvPUB 35分 別 與 擬 南 芥PUB13(63.7%)、PUB19(43.1%)、AT3G01400(75.7%)、AT3G49810(64.2%)有較高的同源性,并且它們之間具有互作關系。PUB13與Ras相關蛋白RABA4b存在相互作用關系,RABA4b屬于Rab家族中的小GTPase亞家族,是一種調節膜轉運的蛋白質。AT3G49810可以與BKI1相互作用,BKI1是一種質膜定位的磷蛋白質,可以直接與BRI1的激酶域相互作用從而抑制激酶活性。PUB19可以與26S蛋白酶體非ATP酶調節亞基11同源物ATS9相互作用,同時ATS9又與26S蛋白酶體非ATP酶調節亞基RPN1A和RPN1B相互作用。這些結果說明甜菜PUB蛋白可能作為E3泛素連接酶對不同種蛋白質進行泛素化修飾,也可與26S蛋白酶體相互作用導致泛素化蛋白質降解。
3 討論
植物生長和發育依賴于植物感知和響應環境及內源信號的能力,因此對植物的適當刺激和適當響應之間的相互作用十分復雜。除了信號傳輸外,信號轉導途徑本身的調節也很重要[14]。泛素介導的蛋白水解作為重要的翻譯后過程在響應發育或適應環境脅迫的真核信號傳導途徑的調節過程中起到重要作用。蛋白質泛素化通過調節各種肽豐度、酶活性或蛋白質定位從而在植株生長發育的各個階段以及抵抗各種生物脅迫和非生物脅迫過程中起關鍵作用[15]。E3泛素連接酶作為泛素化修飾過程中重要的酶,在植物響應非生物脅迫中起到重要作用。U-box型E3泛素連接酶小麥中TaPUB1通過調控干旱脅迫相關基因的表達,提高轉基因植株對干旱的耐受性。除此之外,可促進TaPYL4和TaABI5的降解,降低轉基因小麥對ABA的敏感性[16-17]。鹽脅迫下,TaPUB26影響某些抗氧化酶基因的轉錄表達,破壞細胞的抗氧化酶系統,降低轉基因植株的耐鹽性[18]。在鹽脅迫下上調表達,AtPUB30作為植物E3泛素連接酶可泛素化BKI1并導致其降解,從而負調控植株的耐鹽性[19]。然而AtPUB31在植株響應鹽脅迫過程中起到正向調控的作用[20]。本研究以前期轉錄組數據作為基礎從中篩選鹽脅迫下差異表達的甜菜PUB基因,并對其進行生物信息學分析,為接下來進行U-box型泛素連接酶的研究提供了更充分的理論基礎及參考。此外,對U-box型蛋白的互作蛋白進行預測,26S蛋白酶體亞基相關蛋白、膜轉運相關蛋白、激酶等功能蛋白被預測到,雖然結果僅為預測結果,但與已經報道的植物PUB蛋白存在一定相似性,所以結果可信,可作為后續互作蛋白鑒定的候選蛋白。
4 結論
本研究以甜菜T710-Mu品系為研究對象,利用轉錄組測序數據獲得42個差異表達的PUB基因,對其進行生物信息學分析。發現甜菜PUB基因在甜菜9條染色體上均有定位,依據蛋白結構可將甜菜PUB蛋白歸為ClassⅡ、ClassⅢ、ClassⅣ中,它們具有保守的功能結構域,如U-box,Pkinase、Pkinase-Tyr、Usp、Arm以及KAP結構域,可能是潛在的E3泛素連接酶。這些結果可為接下來研究甜菜U-box型E3泛素連接酶提供理論支持。
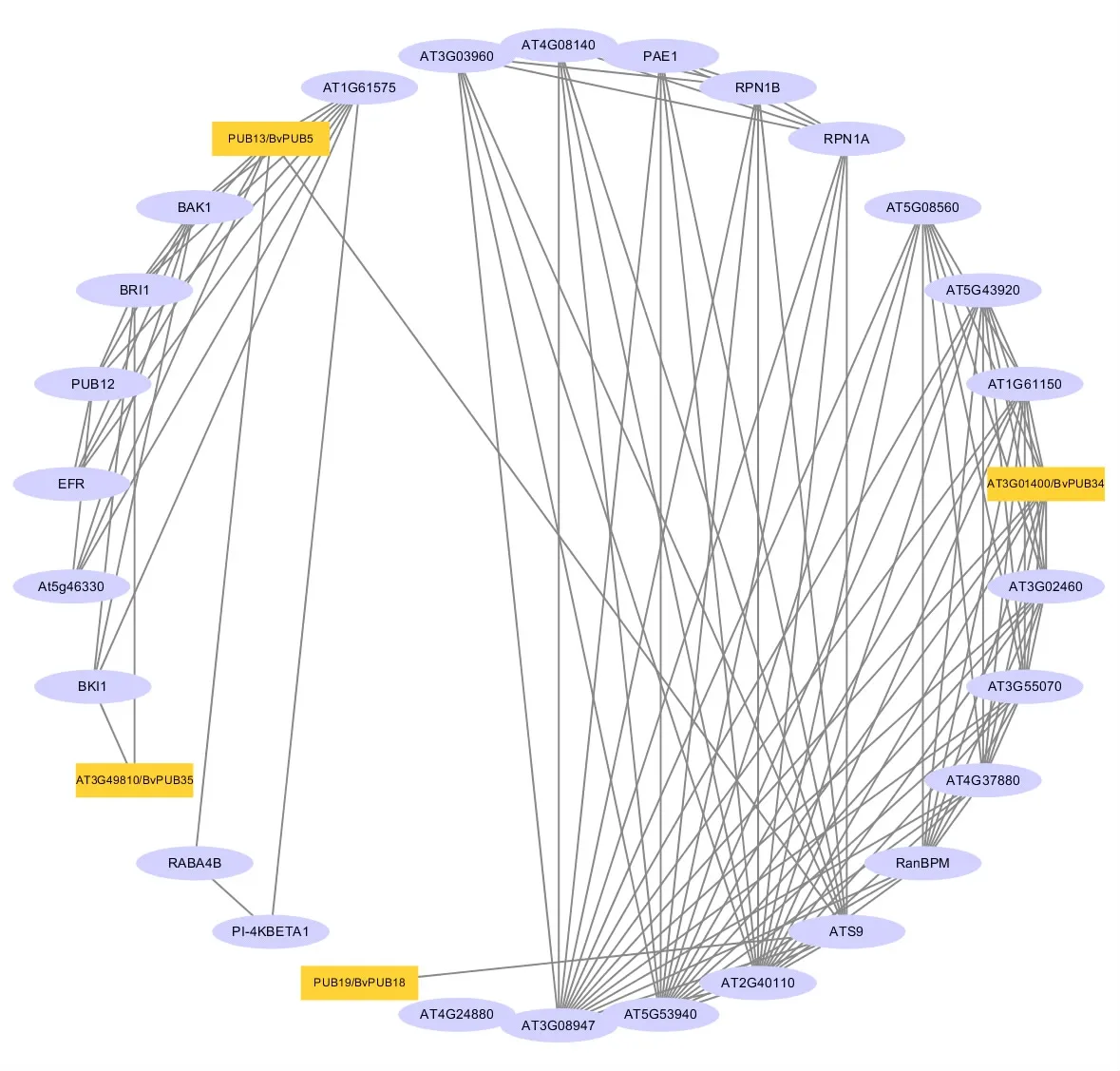
圖6 甜菜PUB蛋白互作網絡圖
猜你喜歡
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
山東工業技術(2016年15期)2016-12-01 05:31:22
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
