氮肥對鎘脅迫下龍葵葉綠素含量和葉綠素熒光特性的影響
2021-12-11 04:36:06唐星林劉光正
西南林業大學學報 2021年6期
關鍵詞:植物
唐星林 劉 斌 劉光正 王 莉 嚴 晶
(江西省林業科學院,江西 南昌 330032)
土壤重金屬污染是我國面臨的主要環境問題之一[1]。《全國土壤污染狀況調查公報》[2]顯示我國土壤總的點位超標率為16.1%,其中,鎘(Cd)的點位超標率高達7.0%。土壤Cd易被植物吸收,并通過食物鏈傳遞給人或者動物,嚴重威脅著人類的健康[3]。Cd是植物非必需元素,其生物毒性很強[4]。研究表明,Cd脅迫會損害植物器官,抑制植物生長[5]。植物體內Cd過量積累會破壞生物膜[6],影響植物體內各種酶的活性[7]。Cd脅迫還會導致植物葉綠素含量、光系統Ⅱ(PSⅡ)量子轉化效率、電子傳遞速率等降低,光合作用受到抑制,最終導致植物生物量下降[8]。氮素是植物生長所必需的大量元素。研究表明,Cd脅迫會減少植物對氮、磷等營養元素的吸收[9],而增施氮肥可以增強植物對Cd脅迫的抗性[10]。研究表明:合理施用氮肥能有效提高Cd脅迫下植物葉綠素含量,保護葉綠體結構,防止葉片出現失綠癥[11-13];以增大PSⅡ反應中心光化學效率和電子傳遞速率[14-15],增強光合作用;進而緩解Cd對植物的毒害,提高植物生物量和Cd積累量[16]。此外,人們發現Cd脅迫下低中氮處理能有效提高葉綠素含量和PSⅡ實際光化學量子產量,但高氮處理沒有顯著的促進作用[17]。這說明氮肥對Cd脅迫的緩解作用還與氮肥施用濃度有關。
龍葵(Solanum nigrum)為茄科(Solanaceae)茄屬(Solanum)多年生草本植物,是我國發現的Cd超積累植物,具有生物量大、適于刈割等特性,是實施土壤重金屬植物修復的優良材料[18]。研究表明,氮肥對Cd脅迫下龍葵生長、抗氧化系統和Cd積累等均有顯著促進作用[16,19]。光合生理是植物最重要的生理活動之一,研究施氮對Cd脅迫下龍葵光合生理的影響,可以更好地揭示氮肥在植物緩解Cd毒害過程中的作用。但Cd脅迫下施氮對龍葵葉綠素含量和PSⅡ功能的影響仍不清楚。葉綠素熒光參數可以反映逆境脅迫下葉片PSⅡ功能的改變情況,是研究植物光合生理特性與逆境脅迫關系的內在探針[20]。捕光色素分子內稟特性決定了葉片對光能的吸收、激發和轉換等過程,可以反映葉片對光能的吸收與利用,同一植物在不同環境條件下具有不同的內稟特性[21]。為此,本研究以龍葵為試驗材料,分析不同濃度氮添加對Cd脅迫下龍葵葉綠素含量、葉綠素熒光特性和捕光色素分子內稟特性等的影響,得出適宜的氮肥施用濃度,為進一步利用龍葵修復Cd污染土壤提供參考依據。
1 材料與方法
1.1 供試土壤
供試土壤為黃壤,取自江西省林業科學院試驗基地,取土深度為0~20 cm。土壤風干、磨碎并過5 mm篩。將黃壤和泥炭按質量比1∶1均勻混合成試驗種植土,其基本理化性質為pH值5.75、水解性氮53.7 mg/kg、有效磷95.0 mg/kg、速效鉀110.0 mg/kg、有機質75.58 mg/kg。
1.2 試驗設計
試驗在江西省林業科學院科研大棚內進行。參考江西省農用土壤Cd污染現狀[22]和魏朵花等[15]的研究進行Cd脅迫和施氮處理,采用完全隨機區組試驗設計,共設置5個處理(見表1)。其中,Cd濃度以Cd2+計。按設定濃度將重金屬Cd2+以CdCl2·2H2O形式拌入種植土,并施入不同濃度的CO(NH2)2,充分混勻,平衡18 d后裝入塑料花盆(上口徑18 cm、下口徑13 cm、高15 cm)。2019年5月28號,將飽滿的龍葵種子用10 g/L KNO3溶液浸泡24 h,超純水沖洗干凈,播種于育苗穴盤中,育苗基質為泥炭,放置于江西省林業科學院科研大棚(28°45′N, 115°49′E)。待幼苗長出5片真葉后(萌發后20 d),選擇長勢一致的幼苗移栽到塑料花盆中進行試驗處理,單株定植,每個處理重復6盆。在盆下放置塑料托盤,將滲漏液倒回盆中,防止污染物和營養物質損失。進行日常養護管理,試驗期間不追施任何肥料。

表1 試驗設計Table 1 Experimantal design mg/kg
1.3 試驗方法
1.3.1 葉片色素含量的測定
葉綠素熒光參數測定結束后,取植物葉片,擦拭干凈,采用打孔法測定葉片色素含量[23]。
1.3.2 葉綠素熒光參數測定
葉綠素熒光參數采用便攜式脈沖調制葉綠素熒光儀PAM?2500(Walz,德國)測定。在Cd脅迫和施氮處理35 d后選取植株頂部生長旺盛的全展葉,測定其熒光誘導動力學參數。先將葉片暗適應30 min以上,打開測量光(小于1 μmol/(m2·s))測定初始熒光(Fo),再在飽和脈沖(脈沖強度為10 000 μmol/(m2·s),脈沖時間0.8 s)激發下測定最大熒光(Fm)。隨后,葉片在光化光(光強為1 160 μmol/(m2·s))照射5 min以上,采用內置程序將光化光光合有效輻射(PAR)梯度設置為1 160、1 663、785、619、363、271、141、101、64、31、6 μmol/(m2·s),測定光下穩態熒光(Fs)、光下最大熒光(Fm′)和光下最小熒光(Fo′),每個光化光梯度照射30 s。計算參數:PSⅡ最大光化學量子產量(Fv/Fm)、PSⅡ有效光化學量子產量(Fv′/Fm′)、光化學淬滅系數(qP)、非光化學淬滅系數(NPQ)和電子傳遞速率(ETR)、天線熱耗散(D)、光化學反應耗散(PH)和非光化學反應耗散(E)[24-25]。每個處理隨機選擇3株進行測定。
1.3.3 光合電子流對光響應的機理模型模擬
利用光合電子流機理模型擬合龍葵光合電子流對光響應的曲線,得出初始斜率(a)、最大電子傳遞速率(ETRmax)和飽和光強(PARsat),以及描述捕光色素分子物理特性的參數(捕光色素分子數(N0)、處于最低激發態的捕光色素分子數(Nk)、捕光色素分子本征光能吸收截面(σik)、捕光色素分子有效光能吸收截面(σik′)),模型的擬合效果用決定系數(R2)表示[26-27]。
1.4 分析方法
光合電子流對光的響應曲線基于光合計算平臺(http://photosynthetic.sinaapp.com/)進行處理。采用單因素方差分析和Tukey HSD法對試驗數據進行方差分析和差異顯著性檢驗(P<0.05)。數據處理和作圖由R語言(R?3.5.1)完成。圖和表中的數據為平均值±標準誤差。
2 結果與分析
2.1 不同施氮量對Cd脅迫下龍葵葉片色素含量的影響
由表2可知,與CK相比,Cd脅迫下龍葵葉綠素a、葉綠素b、類胡蘿卜素含量和N0無顯著變化,但均有下降趨勢。Cd脅迫下龍葵σik顯著大于CK(P<0.05)。與CdN0相比,CdN1、CdN2和CdN3龍葵葉綠素a含量分別顯著增大了52.5%、87.6%和102.2%(P<0.05),葉綠素b含量分別顯著增大了60.5%、92.0%和105.8%(P<0.05),類胡蘿卜素含量分別顯著增大了53.2%、83.3%和91.6%(P<0.05),N0分別顯著增大了68.9%、137.4%和94.5%,σik分別顯著減少了37.3%、52.1%和58.7%(P<0.05)。龍葵葉綠素a、葉綠素b、類胡蘿卜素含量、N0、σik在CdN2與CdN3處理間無顯著差異。這說明Cd脅迫下增施氮肥對龍葵葉片色素含量和捕光色素分子數有顯著的促進作用。
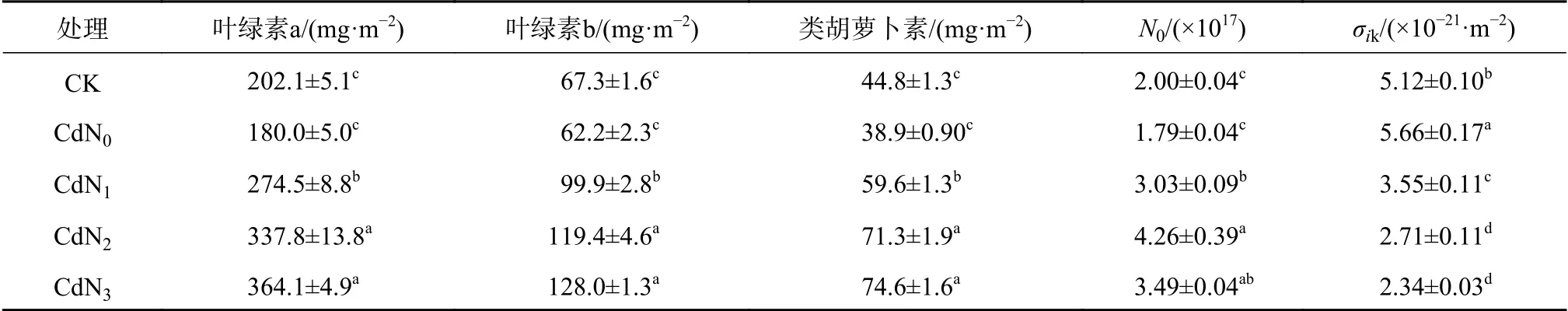
表2 氮肥對Cd脅迫下龍葵葉片色素含量的影響Table 2 Effects of nitrogen supply on pigment contents of S. nigrum under cadmium stress
2.2 氮肥對Cd脅迫下龍葵葉綠素熒光參數的影響
表3中光下葉綠素熒光參數均采用1 663 μmol/(m2·s) PAR下測定的數據。與CK相比,Cd脅迫下龍葵Fv/Fm、Fv′/Fm′和ETR無顯著變化,但均有下降趨勢。CdN2、CdN3處理下龍葵Fv/Fm、Fv′/Fm′顯著大于CdN0(P<0.05),NPQ顯著小于CdN0(P<0.05)。CdN3處理下龍葵qP顯著大于CdN0(P<0.05)。不同處理間ETR的大小順序為CdN3>CdN2>CdN1>CdN0、CK(P<0.05)。Cd龍葵Fv/Fm、Fv′/Fm′、qP、NPQ在CdN2與CdN3處理間(或CdN1與CdN0處理間)均無顯著差異。
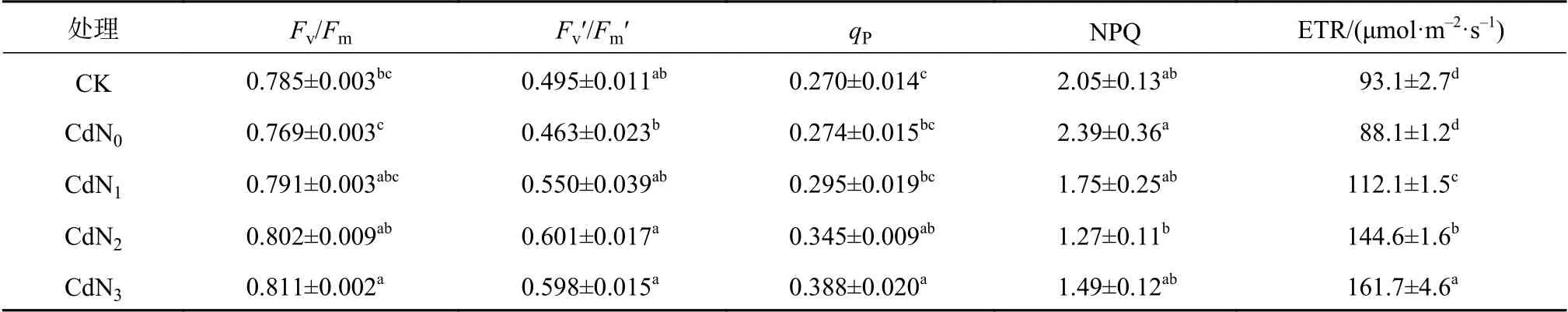
表3 氮肥對Cd脅迫下龍葵葉綠素熒光參數的影響Table 3 Effects of nitrogen supply on chlorophyll fluorescence parameters of S. nigrum under cadmium stress
2.3 氮肥對Cd脅迫下龍葵快速光曲線的影響
由圖1可知,不同處理下龍葵葉片ETR隨PAR的增大而逐漸增大。當PAR<800 μmol/(m2·s)時,ETR幾乎呈線性增加,當PAR>800 μmol/(m2·s)時,ETR呈緩慢增大。當PAR<141 μmol/(m2·s)時,不同處理間ETR差異不顯著。當PAR>141 μmol/(m2·s)時,Cd脅迫下施氮處理ETR顯著大于CK和CdN0處理(P<0.05)。
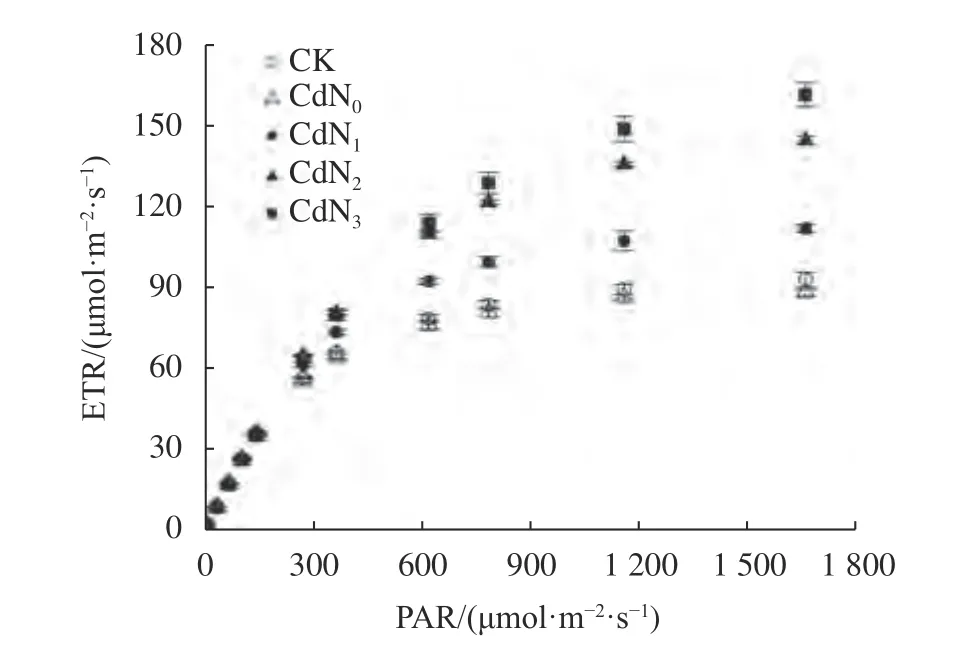
圖1 Cd脅迫下增施氮肥龍葵ETR光響應曲線Fig. 1 Light-response curves of ETR in S. nigrum under cadmium stress by supplement nitrogen
由表4可知,龍葵a、ETRmax和PARsat在不同處理間均存在顯著差異(P<0.05)。與CK相比,Cd脅迫下龍葵ETRmax和PARsat無顯著變化,但均有所減小。不同處理間a的大小順序為CK、CdN0、CdN1>CdN2>CdN3,ETRmax的大小順序為CdN3>CdN2>CdN1>CdN0、CK。CdN3的PARsat顯著大于CdN0(P<0.05)。
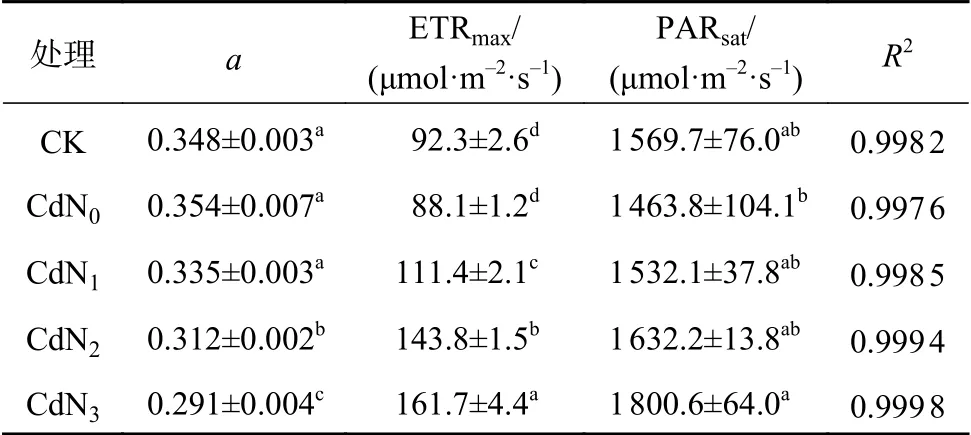
表4 氮肥對Cd脅迫下龍葵葉片a、ETRmax和PARsat光合參數的影響Table 4 Effects of nitrogen supply on a, ETRmax, and PARsat of S. nigrum under cadmium stress
2.4 氮肥對Cd脅迫下龍葵捕光色素分子內稟特性的影響
由圖2可知,隨PAR的增大,不同處理下龍葵葉片Nk逐漸增大,σik′逐漸減小。與CK相比,Cd脅迫下Nk有增大趨勢,σik′有減小趨勢。Cd脅迫下增施氮肥導致Nk增大,σik′減小。CdN2的Nk高于CdN1、CdN3。CdN3的σik′低于CdN1、CdN2。
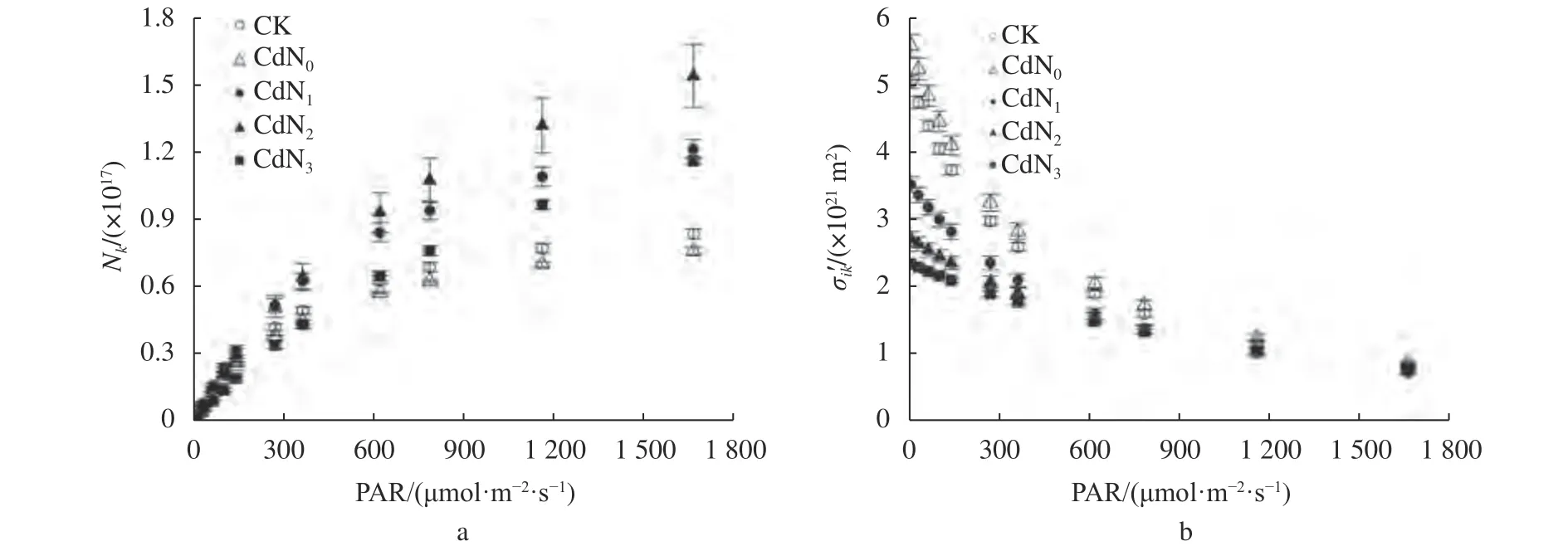
圖2 Cd脅迫下增施氮肥龍葵葉片Nk和σik′的光響應曲線Fig. 2 Light-response curves of Nk and σik′ for S. nigrum under cadmium stress by supplement nitrogen
2.5 葉片吸收光能的分配
由圖3可知,隨著PAR的增加,不同處理龍葵葉片D和E分別從(26.8%~30.3%)、(0.6%~3.4%)增加到(40.2%~53.7%)、(33.7%~39.3%),PH從(12.6%~23.2%)減小到(69.0%~70.2%)。PAR≤363 μmol/(m2·s)時不同處理下龍葵葉片吸收光能的分配均表現為PH>D>E,當PAR≥1 160 μmol/(m2·s)時表現為D>E>PH。當PAR=619 μmol/(m2·s)時,CK、CdN0和CdN1處理表現為D>PH>E,CdN2和CdN3處理為PH>D>E。

圖3 Cd脅迫下增施氮肥龍葵葉片D、PH和E的光響應變化Fig. 3 Light-response curves of thermal energy dissipation, photochemical reaction and non-photochemical energy dissipation in S. nigrum under cadmium stress by supplement nitrogen
表5所示為飽和光強下龍葵葉片吸收光能的分配情況。在1 663 μmol/(m2·s) PAR下,龍葵葉片吸收的光能有40.0%~53.7%用于天線熱耗散,12.6%~23.2%用于光化學反應,33.7%~39.3%用于非光化學反應。在1 663 μmol/(m2·s) PAR下,龍葵D和PH在不同處理間差異顯著(P<0.05),E無顯著差異。與CK相比,Cd脅迫下龍葵D、PH和E無顯著變化,但D有增大趨勢,PH和E有減小趨勢。CdN2、CdN3處理下龍葵D顯著小于CdN0(P<0.05)。不同處理間PH的大小順序為CdN3>CdN2>CdN1>CdN0、CK(P<0.05)。

表 5 氮肥對Cd脅迫下龍葵葉片吸收光能分配的影響Table 5 Effects of nitrogen supply on distribution of absorbed light energy in S. nigrum under cadmium stress
3 結論與討論
光合色素是葉片進行光合作用所必需的物質,主要包括葉綠素和類胡蘿卜素。其中,葉綠素在光能的吸收、傳遞和轉化過程中起主導作用,類胡蘿卜素則參與光能捕獲和光破壞防御過程[28]。重金屬脅迫會抑制葉片光合色素的合成,進而影響植物光合作用[29]。本研究發現Cd脅迫下龍葵葉綠素和類胡蘿卜素含量均呈現下降趨勢,這與蒼耳(Xanthium sibiricum)[30]和洋常春藤(Hedera helix)[20]等植物的研究結果類似。對Cd脅迫導致葉片色素含量下降的原因,主要有3種觀點:一是Cd脅迫影響Fe、Zn、Mg等營養元素的吸收,Cd2+取代葉綠素中的Mg2+而導致葉綠素破壞[31-32];二是Cd脅迫抑制葉綠素合成酶活性,增大葉綠素降解酶活性[33-34];三是Cd抑制了葉綠素合成前體物氨基乙酰丙酸(ALA)的合成[32]。研究表明,Cd脅迫下增施氮肥后,龍葵葉綠素和類胡蘿卜素含量顯著增加,這與結縷草(Zoysia japonica)[16]和小麥(Triticum aestivum)[12]等植物的研究結果一致。氮素能有效促進葉片色素的合成[35],這可以平衡Cd脅迫對葉片色素的抑制作用。張帆等認為Cd脅迫下增施氮肥可以解除Cd對葉綠素合成的抑制[13]。這說明增施氮肥可能通過促進葉綠素合成來提高龍葵的抗Cd能力。
PSⅡ反應中心是一個對重金屬脅迫比較敏感的光合機構[36],葉綠素熒光參數可以反映重金屬脅迫對PSⅡ光合活性及電子傳遞等功能的影響[37]。有研究表明,Cd脅迫會抑制葉片PSⅡ光化學功能[38],而氮素有助于提高PSⅡ的光化學活性[39]。Fv/Fm可以反映植物是否受到逆境脅迫。本研究表明,Cd脅迫下龍葵葉片Fv/Fm呈下降趨勢,說明龍葵受到一定程度的Cd脅迫。而Cd脅迫下增施氮肥使得龍葵Fv/Fm顯著增大,這說明施氮可以有效緩解Cd對龍葵的脅迫作用。Fv′/Fm′反映開放的PSⅡ反應中心原初光能捕獲效率,NPQ反映PSⅡ天線色素吸收的光能以熱能形式耗散的部分,對光合機構起保護作用[25]。本研究發現Cd脅迫下龍葵葉片Fv′/Fm′、ETR下降,NPQ增大,說明Cd脅迫下龍葵葉片PSⅡ光合反應中心實際量子產量下降,PSⅡ向PSI的電子傳遞受阻,使得同化力(NADPH和ATP)形成受阻,PSⅡ過剩光能增加,非光化學淬滅保護機制發揮作用,這與紫花香薷(Elsholtzia argyi)[40]和結縷草[16]等植物的研究結果一致。而Cd脅迫下增施氮肥后龍葵Fv′/Fm′、ETR上升,NPQ減小,說明氮肥能有效緩解Cd脅迫對PSⅡ功能的影響,提高葉片光能捕獲效率和電子傳遞速率,增加同化力(NADPH和ATP)的形成,為固碳反應提供更多的能量,進而增強對Cd毒害的抵抗能力。
葉片的光能吸收能力同時受N0、σik和σik′的影響,其中,光能吸收截面反映捕光色素分子吸收光子的能力[26]。葉片捕光色素分子在吸收光能后會從基態躍遷到激發態,激發態的捕光色素分子通過光化學反應、葉綠素熒光和熱能等3種方式退回基態[41]。因此,處于激發態的捕光色素分子越多,處于基態的捕光色素分子就越少,不利于葉片光能的吸收[26]。本研究發現Cd脅迫下龍葵N0、Nk減小,σik、σik′增大,說明Cd脅迫下龍葵在捕光色素分子總數下降的情況下,可以通過增大捕光色素分子光子吸收能力和處于基態的捕光色素分子的比例來提高植物對光能的吸收。而Cd脅迫下增施氮肥使得龍葵N0、Nk增大,σik和σik′減小。這可能是由于Cd脅迫下施氮使得捕光色素分子總數增大,植物通過減小其捕光色素分子光能吸收能力和處于基態的捕光色素分子數的比例,以防止葉片吸收過量光能。葉片吸收光能的3個耗散途徑為D、PH和E[21]。D是指植物用于光合生化反應的能量。其余過剩的光能主要通過PH和E來消耗,防止其對光合機構的破壞[42]。有研究表明,Cd脅迫使得楨楠幼樹E增大,D和PH減少[42]。本研究發現Cd脅迫下龍葵D有增大趨勢,PH和E有減小趨勢,而增施氮肥后D顯著減小,PH顯著增大。其中,光化學耗散能量增大可能與葉綠素含量、Fv′/Fm′和ETR等的增大有關,說明Cd脅迫下增施氮肥有利于提高葉片吸收光能用于光化學反應的比例,增大葉片吸收光能的利用效率。
綜上所述,增施氮肥有助于提高Cd脅迫下龍葵葉片葉綠素含量、捕光色素分子總數、Fv/Fm、Fv′/Fm′、ETR、Nk和PH等光合生理指標,緩解Cd脅迫對PSⅡ功能和吸收光能分配的影響,進而增強龍葵對Cd脅迫的抗性,而且施加300 mg/kg CO(NH2)2對Cd脅迫下龍葵葉綠素合成和PSⅡ功能的促進作用大于施加150 mg/kg CO(NH2)2,但與施加450 mg/kg CO(NH2)2無明顯差異,因此,300~450 mg/kg CO(NH2)2是合適的施氮濃度,為進一步揭示氮肥對Cd脅迫下龍葵光合生理的影響機制及Cd污染土壤的治理實踐提供參考。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
