田菁相關研究進展與應用現狀
2021-12-08 09:29:23賀亭亭邢錦城劉沖董靜朱小梅趙寶泉溫祝桂趙小慧洪立洲
江蘇農業科學 2021年22期
賀亭亭 邢錦城 劉沖 董靜 朱小梅 趙寶泉 溫祝桂 趙小慧 洪立洲


摘要:土壤鹽漬化是全球嚴峻的生態環境問題之一,我國鹽漬化土壤占全國可利用土地面積的4.9%。豆科草本植物田菁在逆境條件下具有很強的生存能力,目前已成為鹽土改良的先鋒植物之一,并且在工業和醫學領域也發現了其利用價值。對田菁2種不同的結瘤方式進行詳細論述,闡明田菁自身對脅迫環境的響應機制以及內生菌和根際微生物的協同抗逆作用,揭示了田菁莖瘤固氮根瘤菌對禾本科作物的促生作用,并總結了田菁作為夏季綠肥在農業上的應用現狀。田菁相關研究雖然已取得一定的進展,但碎片化的理論并不能構成體系。對現有的研究成果進行總結,以期為后續開展田菁相關的分子機制研究和生產應用奠定基礎。
關鍵詞:田菁;結瘤特點;耐逆機制;生產應用;土壤改良
中圖分類號:S551+.501?? 文獻標志碼: A
文章編號:1002-1302(2021)22-0043-07
收稿日期:2021-03-04
基金項目:沿海鹽堿地農業科學觀測實驗站開放課題(編號:YHS201804);江蘇農業科技援疆-克州鹽堿地改良利用示范基地項目[編號:JATS(2019)228]。
作者簡介:賀亭亭(1986—),女,山東煙臺人,博士,助理研究員,主要從事田菁新材料選育及其促生微生物相關研究。E-mail:ht142857@163.com。
通信作者:邢錦城,碩士,副研究員,主要從事耐鹽植物新品種選育和植物逆境生理方面的研究。E-mail:sdauxxx@163.com。
2018年農業農村部根據中共中央辦公廳、國務院辦公廳《關于創新體制機制推進農業綠色發展的意見》的有關部署,制定了《農業綠色發展技術導則(2018—2030年)》。該導則中提到了綠肥與作物生產相結合的種植模式與利用技術。研究發現,綠肥不但能夠改善土壤理化性質,增強土壤肥力,還能夠減少化肥施用量、降低土壤重金屬含量,改善土壤連作障礙。近年來,綠肥在土壤改良和污染治理過程中發揮了重要作用。
田菁[Sesbania cannabina (Retz.)Poir.]為一年生豆科草本植物,羽狀復葉,高度可達2.0~3.5 m,根系發達,具有2種不同的結瘤方式且固氮能力強、生物量大,是優質的夏季綠肥,在東北、華北、華東、華南、華中和西北地區廣泛種植。田菁具有耐鹽、耐澇、耐旱、耐貧瘠等特點,并且具有較強的抵御病、蟲、草害的能力,但目前的研究主要集中于田菁的耐澇和耐鹽方面。自20世紀60—90年代間,田菁作為綠肥開始對其進行研究,至今已在農業、工業、醫學等方面取得了部分研究成果。本研究從田菁自身入手,闡述其為適應環境所采用的不同結瘤方式,及其響應環境脅迫的應答機制,揭露田菁根系內外微生物的協同抗逆作用,解析田菁及其招募的莖瘤固氮根瘤菌在農業生產中的作用,并總結了田菁在農業、工業、醫學領域的應用現狀及前景,以期為田菁及其相關的深入研究提供借鑒。
1 田菁不同結瘤方式的研究
田菁攜帶有休眠的不定根原基,當被水淹沒時能夠發展成根,進而產生莖瘤,此外還能在側根基部形成根瘤。因此,田菁具有多種結瘤特征和雙根瘤菌侵染途徑,具體如圖1所示[1]。在非淹澇條件下,根瘤菌通過根毛卷曲和形成管狀感染線(infection thread,IT)侵入田菁(圖1 右側)。在淹澇環境下,乙烯大量積累[2],會通過妨礙結瘤因子(nod factor,NF)誘導的鈣振蕩來抑制根毛感染。因此,一種替代嚴格侵染機制的策略發展開來。根瘤菌通過側根基部的表皮裂隙進入外皮層,在細胞間形成感染袋(infection pocket,IP),該過程需要活性氧(reactive oxygen species,ROS)、乙烯和赤霉素(gibberellin,GA)的參與,之后以IP為基礎形成IT,進而產生根瘤(圖1左側)。
在蒺藜狀苜蓿(Medicago truncatula)發生根毛感染的細胞中,NF誘導的鈣振蕩有個快速上升的斜率和一個緩慢下降的斜率[3]。而淹澇條件下生長的田菁根毛中NF誘導的鈣振蕩速率相比之下更快
且勻稱,根毛只是生長和變形而不會被感染。在加入乙烯抑制劑后,鈣振蕩頻率與蒺藜狀苜蓿相似,發現根毛中出現細胞感染的現象[1]。將細胞中鈣-鈣調素依賴性蛋白激酶(CCaMK)基因敲除后,IT和根瘤無法產生,但IP可以正常形成,說明IP的形成并不依賴于鈣振蕩。田菁細胞間感染過程中,雖然SymRK基因不參與皮層感染和根瘤起始信號傳導,但對于IT是必需的,在細菌釋放到植物細胞中及建立功能性共生體的過程中發揮重要作用[4]。田菁根毛的細胞內感染和苜蓿以相同的方式進行,并且需要共同Sym途徑[4]。以上結果表明,細胞間感染在形態學和分子水平上均不同于默認的根毛感染途徑。
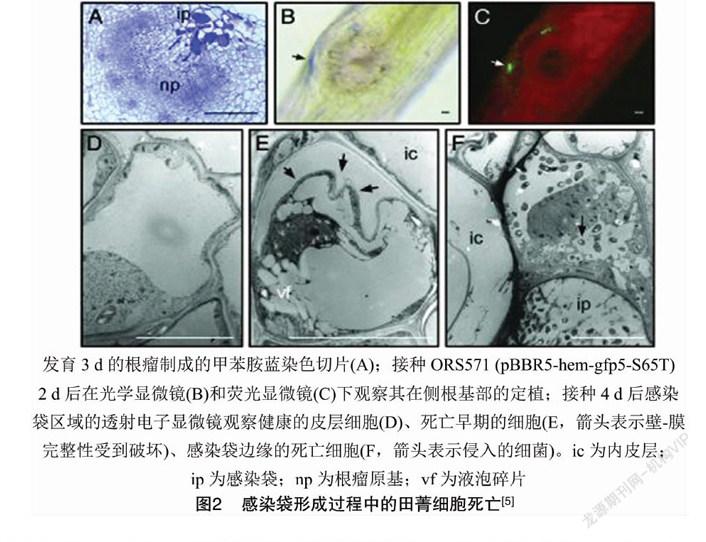
通過透射電子顯微鏡結合綠色熒光蛋白標記細菌發現,細菌的定植伴隨著水培毛萼田菁(Sesbania rostrata)根毛局部細胞的死亡(圖2)。乙烯、ROS合成抑制劑和H2O2清除劑均能夠阻斷Nod因子誘導的根毛卷曲,也沒有細菌入侵、感染袋或根瘤原基形成的跡象,表明內源性乙烯和ROS介導了這些響應過程[5]。毛萼田菁水培根中,乙烯介導了Nod因子觸發側根基部的細胞分裂,增加具有死亡特征的細胞數量,積累酚類化合物,形成帶有殘存細胞壁的空腔,從而強化通氣組織,成為淹澇脅迫反應的一部分[5]。盡管在陸生植物中,乙烯參與停滯生長和成熟,但在半水生植物中,乙烯能夠刺激植株的生長和分裂過程[6]。研究表明,在毛萼田菁中,根瘤原基的起始取決于ROS,H2O2與細菌不同階段的侵染共定位,但在根瘤原基中未檢測到。H2O2首先在與細菌接觸的外皮層細胞的細胞質/膜壁上產生。后來,位于感染袋兩側的植物細胞被大量的H2O2包圍,并具有細胞死亡的特征。細胞間基質和細胞壁以及一些細胞內感染線中含有大量的H2O2[5]。H2O2作為大分子交聯的底物,可能對于細胞壁的硬度和感染線的生長有正向作用[7]。細胞基質中的H2O2似乎并未損害細菌,這可能是由于根瘤菌產生了保護性酶并被胞外多糖所屏蔽[8]。 以上研究結果表明,活性氧和乙烯對水培毛萼田菁側根基部結瘤起到正向作用。
2 田菁耐逆性的研究
田菁具有耐澇、耐鹽、耐旱、耐貧瘠等特點,在脅迫環境下具有較強的生存能力,但是目前的理論研究主要集中于耐澇性和耐鹽性2個方面。
2.1 耐澇
在淹澇脅迫下,生長的田菁與對照組相比可以看到大量的不定根和白色海綿根,根瘤數量明顯增加且生物量增大[9](圖3),根瘤內部微生物中有明顯的多羥基丁酸(polyhydroxybutyricacid,PHB)顆粒。已知PHB顆粒可以作為多余碳的吸收池和還原動力,當細菌暴露在限氧條件下時,可以作為碳和能量的儲備[10]。根系淹澇會影響根瘤中的氮代謝,并導致一系列代謝變化,改變碳水化合物、能量和氨基酸水平[11]。田菁根瘤中的代謝產物差異集中于次級代謝產物、氨基酸和糖類,對照組和脅迫組表現出不同的聚類,差異率約達到80%[9]。其中苯丙素受非生物刺激合成量增加,而基于苯丙素的聚合物(如木質素、木栓質和單寧)對植物的穩定性具有很大貢獻[12]。田菁淹澇脅迫處理3 h,對根系轉錄組分析發現,5個上調的差異表達基因被注釋為編碼醇脫氫酶,乙醇發酵為田菁根部提供了能量,但這些差異表達基因(DEG)在淹澇脅迫27 h的田菁中均未顯示出表達差異[13]。可能是由于淹澇脅迫誘導了田菁根皮層中通氣組織的形成,增加了組織孔隙度,促進了氧在根內的運輸(圖3)。由于低氧誘導的轉錄因子可以介導厭氧反應相關基因的表達,在淹澇脅迫早期,bHLH轉錄因子家族的TF顯著上調,而在淹澇脅迫后期,乙烯響應元件結合蛋白顯著表達。淹澇脅迫田菁根中類胡蘿卜素、類黃酮、類苯丙烷和脂肪酸合成基因下調表達,這些結果與脅迫導致根系總碳水化合物和淀粉合成相關酶、二磷酸腺苷(ADP)-葡萄糖焦磷酸化酶(AGPase)活性下降的研究結果一致,說明田菁根系可以通過調節消耗和產出響應淹澇脅迫[14]。
2.2 耐鹽性
植株幼苗對NaCl耐性的綜合評價分析表明,光葉決明、決明、紫穗槐、海濱木槿和田菁的綜合評價值依次升高,分別為0.260、0.287、0.477、0.726、0.758,由此可見,田菁對NaCl脅迫具有較高的耐性[15]。但在鹽分脅迫處理下,田菁生長和生理指標均受到一定程度的影響(表1)[15]。200 mmol/L NaCl處理叢枝菌根(arbuscular mycorrhiza,AM)與田菁的共生體后,田菁光系統Ⅱ效率值、碳水化合物含量以及ADP-葡萄糖焦磷酸化酶和淀粉合酶的活性得到提高,而葉綠素熒光的非光化學猝滅系數出現降低[16]。與對照相比,AM田菁共生體中與超氧化物歧化酶(SOD)、過氧化氫酶(CAT)、谷胱甘肽還原酶(GR)和過氧化物酶(POX)相關的基因表達水平上調,與抗壞血酸鹽和藻酸鹽代謝以及類胡蘿卜素生物合成的相關途徑得到增強[17]。田菁中與光合作用、活性氧清除和特定轉錄因子有關的基因上調表達,通過與AM共生進一步提高了其自身的耐鹽性[16]。鹽脅迫后,叢枝菌根-田菁共生體中,菌根數量增加,田菁根部脫落酸(abscisic acid,ABA)濃度上升,內源獨腳金內酯(strigolactone,SL)水平提高[18]。NaCl和外源ABA、H2O2能夠誘導SL生物合成基因CCD7、CCD8和信號基因MAX2的上調表達,說明激素調節在田菁的耐鹽過程中發揮著積極的作用[19]。除了叢枝菌根之外,研究人員還從田菁根瘤菌及周圍土壤的宏基因組中克隆到一個命名為RSA的極端嗜鹽淀粉酶,最適NaCl濃度為 5 mol/L,屬于極端嗜鹽酶。RSA嗜鹽淀粉酶是從非嗜鹽菌田菁根瘤菌中獲得的,打破了嗜鹽酶從嗜鹽菌中獲得的傳統[20]。由此可見,田菁除了自身具備一定的耐鹽能力外,與微生物的相互作用進一步增強了其對鹽脅迫環境的適應能力。
3 田菁莖瘤在生產上的應用研究
田菁莖瘤固氮根瘤菌ORS571(Azorhizobium caulinodans ORS571)與毛萼田菁共生,既能形成莖瘤也能形成根瘤,首次由Dreyfus從莖瘤中分離出來[21]。ORS571具有自生固氮能力,也可以與宿主共生固氮。ORS571不但能夠在田菁、銀合歡和菜豆的根部結瘤,還能夠定殖于谷類作物的根部[22]。已有研究表明,ORS571能夠在自然條件下與小麥共生,顯著提高小麥的含氮量和干物質量[23]。ORS571可從根部侵染小麥,定殖于根的表皮、皮層、維管系統的細胞間隙和細胞內,并可以向上遷移到達葉片[24](圖4)。田間試驗結果表明,ORS571浸種侵染對小偃22、小偃6號、西農979、陜253、綿陽19、鄭引1號、周麥18和矮抗58等小麥品種均有較明顯的促生作用。浸種侵染后,幼苗平均根長和株高相較于對照組分別增加了17.04%和8.37%,并且葡萄糖有助于ORS571的浸種侵染和定殖[25]。RT-PCR結果表明,被ORS571侵染的小麥根部中6條與營養元素代謝有關的miRNA相對表達量呈現出先上調后下調的趨勢。通過靶基因預測,miR164、miR167和miR169分別作用于NAC1基因、ARF8基因和HAP基因,能夠促進側根的起始和出現,增加側根的數量,促進根的形態建成和對氮素的吸收。miR398表達受抑制后COX基因表達量增加,參與氧化還原反應,生成的ATP可以為植物根部從土壤中吸收氮素等必需的營養元素提供充足的能量。miR399表達量增加致使靶基因UBC表達下調,可以增加植物對無機磷的吸收和轉運,應對磷素的缺乏[24]。
經過對ORS571全基因組測序后發現,基因組內含有一個共生島,其中包含了多個轉移相關的基因和與結瘤因子合成相關的基因。田菁根瘤菌共生島在屬間能夠發生水平轉移,進而擴大根瘤菌宿主范圍。中慢生根瘤菌屬(Mesorhizobium)的3種根瘤菌均可接受共生島,而中華根瘤菌屬(Sinorhizobium)部分菌株可接受共生島,轉移的頻率為10-8~10-7。共生島轉移后,受體菌便獲得了與新宿主結瘤的能力,固氮酶活性測定后發現所結莖瘤同樣具有固氮活性。將毛萼田菁、紫云英、玉米作為誘導植物,分別檢測其對ORS571共生島水平轉移頻率的影響,發現共生島的轉移頻率至少提高了10倍,不同植物提高的程度不同,可能與植物根系的分泌物成分相關,具體原因還有待研究[26]。
4 田菁在土壤改良中的應用研究
4.1 改善土壤結構
田菁的種植能夠顯著降低土壤容重(6.1%)和黏砂粒(14.2%)含量,提高土壤大團聚體(62.9%)、微團聚體(13.1%)、平均質量直徑(18.0%)和總孔隙度(5.1%),改善土壤結構[28]。黃河三角洲連續3年種植田菁后發現,土壤水分的初始入滲率和穩定入滲率分別是對照的2.53、2.88倍,表明其對土壤結構具有明顯的改善作用[29]。
4.2 提升土壤肥力
松嫩平原連續3年種植田菁后發現地表生物量(不包含田菁)增加了15.2倍,0~20 cm耕層土壤pH值由10.3降到8.3,可溶性鹽含量由5.61 g/kg降到3.07 g/kg,有機質含量由20.03 g/kg升至22.21 g/kg,堿解氮含量由85.4 mg/kg升至 96.3 mg/kg[29]。天津濱海地區0~20 cm耕層土壤種植田菁后平均脫鹽率高達45.97%,0~80 cm土層土壤脫鹽率為10.81%,其脫鹽效果優于苜蓿、蘇丹草和堿蓬[30]。田菁秸稈不做還田處理的情況下會造成耕層土壤中速效P含量和速效K含量降低。江蘇鹽城濱海種植田菁和草木樨一個生長季后,土壤速效P含量降幅為2.17%~35.6%,翻壓處理后速效P含量和速效K含量逐年回升[31]。上海市郊主要園藝場設施大棚因廣泛分布的Ca2+、Mg2+次生鹽漬化土壤而造成耕作障礙,利用γ-聚谷氨酸和田菁聯合修復后,Ca2+、Mg2+處理效果顯著,最大去除率分別達到90.26%和69.92%[32]。
4.3 吸收土壤重金屬
土壤中過高含量的重金屬能夠抑制植株的固氮作用。但毛萼田菁的莖瘤巧妙避免了與重金屬土壤的直接接觸,從而正常發揮固氮作用。在研究毛萼田菁在鉛鋅礦尾礦中結瘤、固氮和重金屬影響的試驗中發現,保留莖瘤與去除莖瘤相比,植株全氮量和氮積累量分別提高了12.4%~34.0%和43.3%~131.4%,對鎘、銅、鉛、鋅4種重金屬的積累量逐漸遞增,其中根部含量顯著高于莖和葉,并隨著土壤介質中重金屬含量的增加而增加,尾礦土壤中植株鉛和鋅的含量分別是純土壤中的3.2、4.8倍[33]。
4.4 改善后茬作物品質抑制雜草生長
在溫室內種植甜玉米和田菁并全量還田后,發現后茬茄子果實數量分別較對照提高了23.61%和9.72%,總產量分別提高了24.15%和8.19%。田菁還能有效改善茄子品質,與對照相比,維生素C含量增加32.96%,可溶性糖含量增加10.56%,其效果顯著優于高丹草[34]。此外,利用田菁淋溶液和田菁秸稈模擬還田試驗發現,田菁可以通過化感作用抑制發育初期的惡性雜草稗草和三葉鬼針草的生長,尤其對稗草的影響最為顯著。同時,在田菁的影響下,受體植物所在土壤中微生物數量和土壤酶活性得到顯著提升[35]。因此,合理利用田菁不但能夠活化土壤,提升土壤肥力,還能提高下茬作物品質,抑制田間雜草生長。
5 田菁的應用前景及展望
在醫學領域,刺田菁[Sesbania bispinosa (Jacq.)]種子中含有2.01%的左旋多巴,已證明對帕金森病有效[36-37]。Shivamadhu等的研究表明,田菁葉片中的蛋白水解物可能在止血和傷口愈合過程中發揮作用[38]。在工業方面,田菁胚乳中的田菁膠已經被廣泛應用在建筑、陶瓷、紡織、造紙和電池制造等領域中。已有試驗表明,田菁膠能夠替代目前大量進口的瓜爾膠,有望大幅降低產品成本,提高企業利潤[39]。在農業方面,田菁因其結瘤固氮作用,無法消除NO-3型次生鹽漬化土壤造成的連作障礙[32]。該問題可以通過田菁與需氮量大的禾本科作物輪作的方式得到解決。該種模式不但能夠調節土壤肥力,還能均衡利用土壤養分和防治病、蟲、草害,達到增產增收的目的。田菁具有較強的耐逆性,除了其自身相關的響應機制外,植株內部定殖的內生菌和根際微生物均增強了其在不良環境中的生存能力。除了田菁莖瘤固氮根瘤菌ORS571之外,從田菁根際土壤中獲得的普沙根瘤菌 (Rhizobium pusense)YIC4105被用來防治有毒物質根皮苷積累引起的蘋果幼苗連作障礙,并有望開發為具有促生功能的微生物接種劑和菌肥[40]。另一種從田菁根際土壤中分離得到的菌種N102能夠顯著提高鹽脅迫下的玉米株高、根長、葉綠素含量、植株干質量等相關指標,并且對多種植物具有促生耐鹽作用,具有成為鹽堿地作物菌肥的潛力[41]。田菁根瘤及其根際土壤中還包含大量的固氮、解磷、解鉀等功能性微生物。因此,在農業生產中以提高產量、提升質量、降低農藥和化肥施用量、增強耐逆性為目標,以田菁在土壤中招募的特色微生物為基礎,篩選出能夠定殖于作物、蔬菜、果樹根際甚至其內部的有益菌種,研制成菌劑和菌肥具有廣闊的市場前景。現有研究成果表明,長喙田菁細胞間感染與根毛感染相比復雜性較低[42],因此,更容易在非豆科植物中實現應用。結合微生物可以通過基因組島水平轉移的特點,未來田菁有可能在功能性微生物應用到非豆科農作物生產過程中發揮重要橋梁作用。此外,田菁已成為研究根毛感染和細胞間感染的新型模式生物,可以在許多方面補充豆類模型的基礎研究。未來,隨著相關研究的深入,田菁有可能在更多的領域發揮作用。
參考文獻:
[1]Capoen W,Oldroyd G,Goormachtig S,et al.Sesbania rostrata:a case study of natural variation in legume nodulation[J]. New Phytologist,2010,186(2):340-345.
[2]Bailey-Serres J,Voesenek L A C J.Flooding stress:acclimations and genetic diversity[J]. Annual Review of Plant Biology,2008,59:313-339.
[3]Ehrhardt D W,Wais R,Long S R.Calcium spiking in plant root hairs responding to Rhizobium nodulation signals[J]. Cell,1996,85(5):673-681.
[4]Capoen W,Goormachtig S,de Rycke R,et al.SrSymRK,a plant receptor essential for symbiosome formation[J]. Proceedings of the National Academy of Sciences of the United States of America,2005,102(29):10369-10374.
[5]Mitra S,Mukherjee A,Wiley-Kalil A,et al.A rhamnose-deficient lipopolysaccharide mutant of Rhizobium sp.IRBG74 is defective in root colonization and beneficial interactions with its flooding-tolerant hosts Sesbania cannabina and wetland rice[J]. Journal of Experimental Botany,2016,67(19):5869-5884.
[6]Bragina T V,Rodionova N A,Grinieva G M.Ethylene production and activation of hydrolytic enzymes during acclimation of maize seedlings to partial flooding[J]. Russian Journal of Plant Physiology,2003,50(6):794-798.
[7]Wisniewski J P,Rathbun E A,Knox J P,et al.Involvement of diamine oxidase and peroxidase in insolubilization of the extracellular matrix:implications for pea nodule initiation by Rhizobium leguminosarum[J]. Molecular Plant-Microbe Interactions,2000,13(4):413-420.
[8]Santos R,Hérouart D,Puppo A,et al.Critical protective role of bacterial superoxide dismutase in Rhizobium-legume symbiosis[J]. Molecular Microbiology,2000,38(4):750-759.
[9]Krishnan H B,Oehrle N W,Alaswad A A,et al.Biochemical and anatomical investigation of Sesbania herbacea (Mill.) McVaugh nodules grown under flooded and non-flooded conditions[J]. International Journal of Molecular Sciences,2019,20(8):1824.
[10]Quelas J I,Mesa S,Mongiardini E J,et al.Regulation of polyhydroxybutyrate synthesis in the soil bacterium Bradyrhizobium diazoefficiens[J]. Applied and Environmental Microbiology,2016,82(14):4299-4308.
[11]Oliveira H C,Freschi L,Sodek L.Nitrogen metabolism and translocation in soybean plants subjected to root oxygen deficiency[J]. Plant Physiology and Biochemistry,2013,66:141-149.
[12]Wang L H,Zhang Y X,Qi X Q,et al.Global gene expression responses to waterlogging in roots of sesame (Sesamum indicum L.)[J]. Acta Physiologiae Plantarum,2012,34(6):2241-2249.
[13]Colmer T D.Aerenchyma and an inducible barrier to radial oxygen loss facilitate root aeration in upland,paddy and deep-water rice (Oryza sativa L.)[J]. Annals of Botany,2002,91(2):301-309.
[14]Ren C G,Kong C C,Yan K,et al.Elucidation of the molecular responses to waterlogging in Sesbania cannabina roots by transcriptome profiling[J]. Scientific Reports,2017,7:9256.
[15]魏秀君,殷云龍,蘆治國,等. NaCl脅迫對5種綠化植物幼苗生長和生理指標的影響及耐鹽性綜合評價[J]. 植物資源與環境學報,2011,20(2):35-42.
[16]Ren C G,Kong C C,Yan K,et al.Transcriptome analysis reveals the impact of arbuscular mycorrhizal symbiosis on Sesbania cannabina expose to high salinity[J]. Scientific Reports,2019,9:2780.
[17]Niyogi K K.PHOTOPROTECTION REVISITED:genetic and molecular approaches[J]. Annual Review of Plant Physiology and Plant Molecular Biology,1999,50(1):333-359.
[18]Ren C G,Kong C C,Xie Z H.Role of abscisic acid in strigolactone-induced salt stress tolerance in arbuscular mycorrhizal Sesbania cannabina seedlings[J]. BMC Plant Biology,2018,18(1):74.
[19]Pandey A,Sharma M,Pandey G K.Emerging roles of strigolactones in plant responses to stress and development[J]. Frontiers in Plant Science,2016,7:434.
[20]楊 媛,姚甜甜,熊海濤,等. 田菁根瘤菌嗜鹽α-淀粉酶的基因克隆表達與分子改造[J]. 廣西科學,2017,24(2):201-205.
[21]Dreyfus B L,Dommergues Y R.Nitrogen-fixing nodules induced by Rhizobium on the stem of the tropical legume Sesbania rostrata[J]. FEMS Microbiology Letters,1981,10(4):313-317.
[22]Senthilkumar M,Madhaiyan M,Sundaram S P,et al.Induction of endophytic colonization in rice (Oryza sativa L.) tissue culture plants by Azorhizobium caulinodans[J]. Biotechnology Letters,2008,30(8):1477-1487.
[23]李 濤. 柚皮素與菜豆根瘤菌CFN42中基因RHERS01180對田菁根瘤菌共生島水平轉移的影響[D]. 南京:南京農業大學,2017:12-13.
[24]陳永超,齊懷廷,王小晶,等. 田菁莖瘤固氮根瘤菌對小麥葉組織的促生作用研究[J]. 西北植物學報,2016,36(7):1383-1390.
[25]劉華偉,孫 超,楊 呼,等. 田菁莖瘤固氮根瘤菌對小麥種子侵染的促生作用及其在根系內的定殖[J]. 植物營養與肥料學報,2012,18(1):210-217.
[26]吳 萍. 植物誘導田菁根瘤菌共生島屬間水平轉移擴大根瘤菌宿主范圍的研究[D]. 南京:南京農業大學,2014:42-44.
[27]Qiu L,Li Q A,Zhang J B,et al.Migration of endophytic diazotroph Azorhizobium caulinodans ORS571 inside wheat (Triticum aestivum L) and its effect on microRNAs[J]. Functional & Integrative Genomics,2017,17(2/3):311-319.
[28]謝文軍,王濟世,靳祥旭,等. 田菁改良重度鹽漬化土壤的效果分析[J]. 中國農學通報,2016,32(6):119-123.
[29]侯賀賀,王春堂,王曉迪,等. 黃河三角洲鹽堿地生物措施改良效果研究[J]. 中國農村水利水電,2014(7):1-6.
[30]王立艷,潘 潔,肖 輝,等. 種植耐鹽植物對濱海鹽堿地土壤鹽分的影響[J]. 華北農學報,2014,29(5):226-231.
[31]朱小梅,溫祝桂,趙寶泉,等. 種植綠肥對濱海鹽漬土養分及鹽分動態變化的影響[J]. 西南農業學報,2017,8(30):1894-1898.
[32]唐 冬.γ-聚谷氨酸和耐鹽植物聯合修復設施栽培鹽漬化土壤[D]. 重慶:西南大學,2015:8-9.
[33]簡曙光,楊中藝. 莖瘤對長喙田菁在鉛鋅尾礦環境適應中的意義 Ⅱ.莖瘤對長喙田菁固氮和積累重金屬的影響[J]. 植物生態學報,2002,26(2):209-215.
[34]胡曉珊. 綠肥對溫室土壤性狀及后茬茄子產質的影響[D]. 海口:海南大學,2015:45-46.
[35]秦俊豪,溫 瑩,李君菲,等. 綠肥植物田菁的化感效應及對土壤肥力的影響[J]. 土壤,2015,47(3):524-529.
[36]Gautam B,Vadivel V,Stuetz W,et al.Bioactive compounds extracted from Indian wild legume seeds:antioxidant and type II diabetes-related enzyme inhibition properties[J]. International Journal of Food Sciences and Nutrition,2012,63(2):242-245.
[37]Katzenschlager R,Evans A,Manson A,et al.Mucuna pruriens in Parkinsons disease:a double blind clinical and pharmacological study[J]. Journal of Neurology,Neurosurgery,and Psychiatry,2004,75(12):1672-1677.
[38]Shivamadhu M C,Balaji K S,Jayarama S.Haemostatic property of new cystein protease(s) from Sesbania grandiflora:Its action on fibrinogens[J]. Biocatalysis and Agricultural Biotechnology,2017,12:10-14.
[39]李 睿,賈 鑫,王 晨,等. 田菁膠的改性和應用的研究進展[J]. 中國食物與營養,2019,25(7):52-55,20.
[40]任承鋼,李 巖,解志紅,等. 根際促生菌及其應用:CN201711374688.X[P]. 2018-04-20.
[41]王艷霞,解志紅,張 蕾,等. 田菁根際促生菌的篩選及其促生耐鹽效果[J]. 微生物學報,2020,60(5):1023-1035.
[42]Capoen W,Den Herder J,Rombauts S,et al.Comparative transcriptome analysis reveals common and specific tags for root hair and crack-entry invasion in Sesbania rostrata[J]. Plant Physiology,2007,144(4):1878-1889.
