綠僵菌mad2敲除株構建及其生物學和誘導植物響應的功能分析
2021-12-07 05:43:32蔡霓閆多子農向群王廣君涂雄兵張澤華
中國農業科學 2021年22期
關鍵詞:植物
蔡霓,閆多子,農向群,王廣君,涂雄兵,張澤華
綠僵菌2敲除株構建及其生物學和誘導植物響應的功能分析
蔡霓,閆多子,農向群,王廣君,涂雄兵,張澤華
中國農業科學院植物保護研究所植物病蟲害生物學國家重點實驗室,北京 100193
【目的】昆蟲病原真菌綠僵菌兼具植物內生性。已知黏附素MAD2是綠僵菌兩種黏附蛋白之一,在實現綠僵菌與植物的黏附、定殖中起重要作用,但其作用機理知之甚少。本研究通過構建金龜子綠僵菌()2敲除突變株(Δ2),探究MAD2蛋白對綠僵菌生物學功能的影響。【方法】從NCBI中獲取2前后基因組DNA序列,設計擴增2前后同源臂特異性引物,以綠僵菌基因組DNA為模板,擴增得到前后同源臂基因S1、S2;設計特異性引物Hyg-F/R,以pKH-KO載體為模板,擴增得到帶有啟動子序列的潮霉素基因;再通過overlap PCR構建2的同源敲除盒S1H、S2H;最后利用PEG介導的原生質體轉化,獲得穩定遺傳的2敲除株。通過對比敲除株與野生株(WT)的生長特性、黏附作用、殺蟲毒力以及誘導花生共生基因轉錄水平的變化,分析MAD2蛋白的生物學功能。【結果】原生質體轉化獲得了敲除2的同源重組轉化子;敲除株與野生株對比分析表明,敲除株的孢子萌發率顯著低于野生株,萌發中時間比野生株延長5.47 h;培養12 h和14 h時,敲除株的菌絲長度顯著均小于野生株,分別為野生株的77.8%和76.3%;培養12 d的產孢量也比野生株減少33.3%。敲除株對洋蔥內表皮的黏附力明顯降低,但對蝗蟲后翅的黏附性無顯著影響。敲除2并不影響綠僵菌對家蠶的毒力。敲除株處理花生12 h后,與野生株處理相比,花生共生受體、鈣信號解碼相關基因(、和)、脂質氮素轉運相關基因(1、24、2)的轉錄水平出現顯著下調;而與空白對照相比2缺失后轉錄水平仍有一定的上調和的轉錄水平產生顯著抑制對2124的轉錄無明顯影響。【結論】金龜子綠僵菌黏附素MAD2影響菌株的孢子萌發、早期菌絲生長、產孢及對植物的黏附力,但對昆蟲的黏附和殺蟲毒力無影響;在菌株與花生互作早期,MAD2觸發了花生共生基因的轉錄。
金龜子綠僵菌;黏附素MAD2;基因敲除;黏附性;殺蟲毒力;生長特性
0 引言
【研究意義】金龜子綠僵菌()是一種重要的昆蟲病原真菌,在自然界中廣泛分布,兼具昆蟲寄生、土壤腐生和植物內生的生活方式[1-2]。近年的研究證明綠僵菌能與植物建立內共生關系[3],并參與調節植物的生長抗逆和病害持續防控[4-6],但共生建立過程并不清楚。黏附素2(MAD2)被報道與綠僵菌黏附植物有關[7],研究MAD2對綠僵菌生物學的影響以及在調控綠僵菌與植物互作中發揮的作用具有重要意義。【前人研究進展】真菌與植物共生的建立是二者相互作用的過程,黏附是互作的早期階段。黏附包括表面附著和分泌黏附物質固著等進程,微生物的黏附物質對特定宿主物種的識別和結合有高度特異性[8]。目前已發現綠僵菌有兩種黏附蛋白,黏附素1(MAD1)和黏附素2(MAD2)。比較兩種黏附素基因在不同種類和株系的綠僵菌中核苷酸序列保守性,發現2相對于1有較大的進化差異[7],MAD2由306個氨基酸組成,有3個主要結構域,結構類似于白色念珠菌(s)的細胞壁ALS(agglutinin-like sequence)蛋白[9]。相對于MAD1對昆蟲表皮的黏附作用,MAD2是支持綠僵菌黏附植物的重要物質,并且在植物提取物處理綠僵菌后,2在4 h和12 h表達顯著上調[10],說明2的進化更有可能是綠僵菌適應植物的重要因素。近期研究發現MAD1蛋白處理的花生()根6 h時已激活膜受體基因和,并抑制花生免疫級聯基因、、及防御相關基因、的表達,推測MAD1抑制免疫有助于綠僵菌在花生根組織上定殖[11]。有關綠僵菌施用后調節植物對微量元素的吸收、促進植物生長和增強植物抗性的報道很多,例如,將棕色綠僵菌()應用于土壤,可以改善高粱()對鐵的吸收并促進其生長[4];在馬鈴薯()植株上施用棕色綠僵菌后,馬鈴薯生物量和植株磷含量顯著增加[12]。對同位素15N的追蹤發現,羅伯茨綠僵菌()通過菜豆()和柳枝稷()內共生將昆蟲體內的氮轉移到植物中,而綠僵菌定殖后,植物也將光合作用產物提供給綠僵菌[13-15]。與未施用綠僵菌的植物相比,綠僵菌定殖后的菜豆側根和根毛形成數量顯著增加,但缺失2的綠僵菌處理植物后,根毛增殖的時間較野生株處理顯著延長[16]。在大豆()中應用金龜子綠僵菌LHL07后,植株的大豆異黃酮含量顯著提高,抗非生物脅迫的能力顯著增強[5],而用不同種的綠僵菌處理玉米()后,發現綠僵菌影響玉米早期的營養生長,使植株能夠較早地建立對生物和非生物脅迫的抗性優勢,增加生長量,而這種促進作用在施用綠僵菌2敲除株時消失[17]。很多學者還通過不同方法證實綠僵菌能定殖植物根部。從菜豆植株的不同部位進行真菌分離,發現綠僵菌在植物根部被分離出來[18]。利用乳酚棉蘭染色,顯示羅伯茨綠僵菌以菌絲聚集的形式存在于柳枝稷根皮層細胞內和細胞間隙之間[16],另外利用以增強型綠色熒光蛋白(enhanced green fluorescent protein,eGFP)標記的金龜子綠僵菌處理玉米,14 d后發現,根內有沿細胞延伸的菌絲[19]。這些都證實綠僵菌的植物內生性,但有關綠僵菌定殖植物的過程和機理極少闡述,MAD2在綠僵菌定殖植物過程中發揮的作用并不清楚。【本研究切入點】前期對綠僵菌處理花生根的轉錄組進行了分析,發現共生建立相關的基因有差異響應[20]。本文在綠僵菌影響花生共生基因表達的基礎上,對2缺失后綠僵菌菌株的生長特性、毒力,以及對花生根共生相關基因的影響進行研究。【擬解決的關鍵問題】利用同源重組的方法構建2缺失突變株,探究2在綠僵菌生長以及植物定殖中的作用,為研究與利用2以及綠僵菌的改良提供生物材料和相關功能數據。
1 材料與方法
試驗于2019—2020年在中國農業科學院植物保
護研究所植物病蟲害生物學國家重點實驗室完成。
1.1 供試生物材料
金龜子綠僵菌Ma9菌株為本實驗室保存的潮霉素敏感菌株,添加1%酵母浸出粉的PDA為適用培養基。基因敲除載體pKH-KO含帶有啟動子序列的潮霉素抗性基因,由中國農業科學院植物保護研究所李梅研究員惠贈。家蠶()4齡幼蟲由山東青州廣通蠶農場室內飼養,用于菌株生物測定。東亞飛蝗()為本實驗室飼養,取后翅用于菌株對昆蟲黏附能力測定。洋蔥從超市購買,取內表皮用于菌株對植物黏附能力測定。花生‘魯花11’品系,從農資商店采購種子,室內種植,14日齡幼苗用于2敲除株(Δ2)與野生株(wild-type,WT)對花生根處理,檢測共生相關基因差異響應。
1.2 mad2敲除盒的構建
1.2.1 菌絲體培養和總DNA提取 將新培養的孢子用0.05%的無菌Tween-80配制成孢子懸浮液,通過血球計數板計數后吸取適量接種于200 mL產菌絲培養基(1 L ddH2O中含MgSO42 g,蔗糖20 g,酵母粉10 g,K2HPO45 g),至終濃度1×106孢子/mL,28℃培養3 d。菌絲通過真空泵抽濾,并用100 mL無菌水淋洗兩次,抽干后收集菌絲體,放入液氮中速凍。利用Plant Genomic DNA purification Kit試劑盒(Genemark生物技術有限公司)提取綠僵菌的基因組DNA。
1.2.2和2前后同源臂的克隆 設計的5對特異性引物見表1。以引物對mad2 S1-F/R、mad2 S2-F/R分別對Ma9基因組DNA進行PCR擴增;以引物對Hyg-F/R對載體pKH-KO進行PCR擴增。依照Takara公司PCR高保真擴增酶說明書配制反應體系,反應溫度為95℃ 5 min,95℃ 30 s,55℃ 30 s,72℃ 1.5 min,4℃保溫,循環35次。PCR產物經1%瓊脂糖凝膠電泳檢測后,用PCR產物純化回收試劑盒(Biomed公司)回收,獲得同源前臂S1、同源后臂S2和帶啟動子Ptrpc序列的。參考徐芳等[21]使用overlap PCR進行序列拼接,最終獲得同源前臂與潮霉素連接片段S1H,同源后臂與潮霉素連接片段S2H(圖1)。

表1 載體構建及目標片段特異性引物
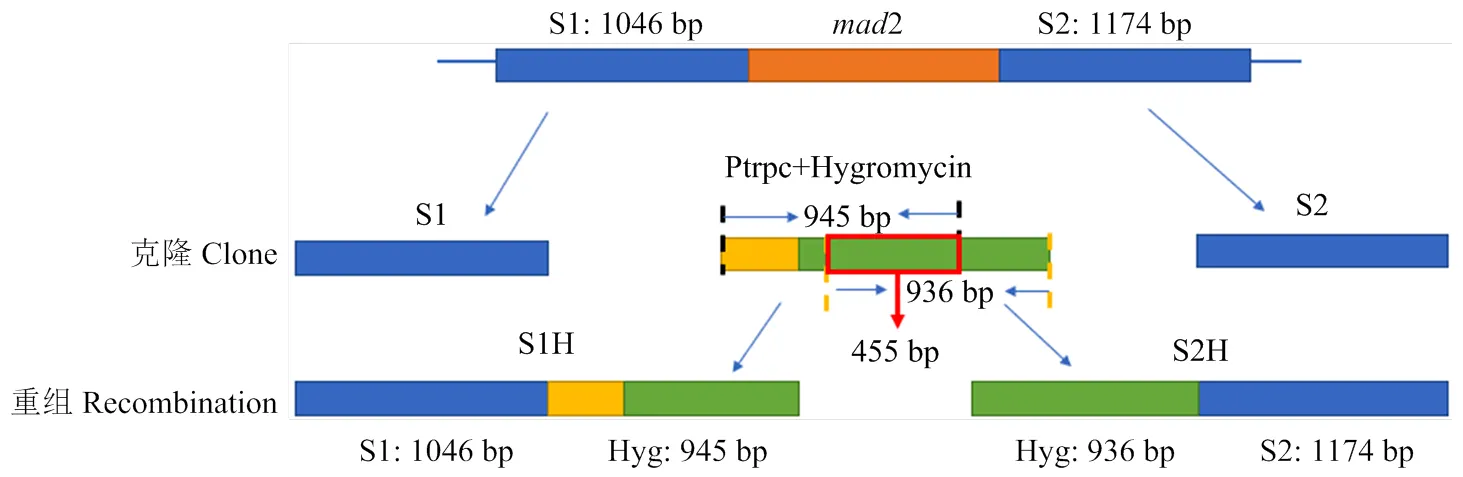
圖1 敲除盒構建步驟
1.3 敲除盒轉化綠僵菌原生質體
1.3.1 原生質體制備 菌絲體培養:接種金龜子綠僵菌Ma9孢子懸浮液3×108孢子于100 mL產菌絲培養基,27℃、180 r/min 振蕩培養20 h,得幼嫩菌絲體。用Miracloth濾布(22—25 μm,北京諾博萊德科技有限公司)過濾,用10 mL 0.7 mol·L-1的NaCl溶液沖洗3次,收集菌絲。
菌絲的酶解:取0.1 g濕菌絲置于50 mL的離心管中,加入20 mL用0.7 mol·L-1NaCl配制的0.1%蝸牛酶液(北京博奧拓達科技有限公司),于30℃、80 r/min解離3 h。用Miracloth過濾除去殘余菌絲,得到原生質體,并用20—30 mL 0.7 mol·L-1的NaCl溶液清洗兩次。用STC緩沖液(蔗糖200 g,1 mol·L-1Tris-HCl(pH 8.0)50 mL,CaCl25.55 g,加ddH2O定容至1 L)重懸,離心清洗兩次,將原生質體懸于600 μL STC buffer中,顯微鏡下計數,調終濃度至(2—5)×107個/mL。
1.3.2 原生質體轉化及轉化子檢驗 參照王曉玲等[22]CaC12-PEG介導原生質體轉化法,將敲除盒S1H、S2H加入綠僵菌Ma9原生質體,與40% PEG 8000混合后在28℃孵育20 min。然后涂布在含低熔點瓊脂糖的TB3(酵母提取物3 g,酶水解酪素3 g,蔗糖200 g,蒸餾水定容至1 L)固體培養基(含Amp濃度100 μg·mL-1,潮霉素濃度300 μg·mL-1)上,28℃恒溫培養3 d,獲得菌落為假定轉化子。
參照上述1.2方法提取轉化子基因組DNA,以潮霉素引物Hyg-F/R,及2敲除驗證引物OUT-mad2- F/R進行PCR檢驗,判斷確定轉化成功的同源重組子,即2敲除株。
1.4 mad2敲除株黏附性驗證
將金龜子綠僵菌Ma9野生株和敲除株接種于PDAY平板上,28℃培養14 d獲得新鮮孢子粉,以0.05% Tween-20制備孢子懸浮液,經過血球計數板計數,將孢子濃度調整至1×107孢子/mL。參考劉艷微[23]的方法稍加改進,取蝗蟲后翅和洋蔥內表皮各3片,分別用37% H2O2殺菌5 min,置于直徑5 cm含有10 mL孢子懸浮液的培養皿中,30 r/min振蕩1 min,用無菌鑷子夾取材料平鋪于水瓊脂培養基上,28℃培養8 h后,用0.05%的Tween-20 30 r/min振蕩30 s以去除多余的分生孢子。處理的材料在光學顯微鏡下觀察,每片計數5個視野的分生孢子數量,統計分析孢子的黏附性。
1.5 mad2敲除株的生物學測定
孢子萌發率測定:同前1.2.1配制野生株與敲除株分生孢子懸浮液,將濃度調整至1×106孢子/mL,分別吸取10 μL懸浮液,均勻涂布于直徑5 cm、含有5 mL 1%的水瓊脂培養基的培養皿中,28℃培養。分別在6、10、14、18、24、36 h觀察孢子的萌發率。每次計數100個孢子,以出芽長度大于孢子長度1/2視為孢子已萌發,記錄萌發數和未萌發數,每皿記錄2組,記錄3皿。計算分生孢子萌發率,萌發率(%)=(孢子萌發總數/鏡檢孢子總數)×100。
采用Excel 2010進行回歸分析,得出綠僵菌孢子萌發率與萌發時間的回歸直線方程和相關性,計算孢子萌發率達到50%所需時間,即萌發中時。
生長速率測定:在記錄萌發率的同時,在10、12、14、18 h測定敲除株與野生株菌絲長度。在顯微鏡(重慶澳浦光電技術有限公司)下,將顯微鏡電腦顯示器分辨率參數設置為5 440×3 648(27×18 cm顯示框),將顯微鏡放大倍數調整至10×40,將視野調整清晰后進行視野捕獲,使用軟件中的測量工具對視野內孢子的長度(L)進行測量,隨機選取5個視野,每個視野中孢子數不少于20個,對5個視野中的孢子長度分別取平均數。根據分辨率計算菌絲長度:菌絲長度(μm)=(27/5440)×L/400×102。
產孢量測定:參照蔡守平等[24]的方法并加以改進。同前1.2.1配置野生株與敲除株分生孢子懸浮液,并調整濃度至1×106孢子/mL,用微量移液器分別點滴接種10 μL于PSAY平板中央,各重復5皿。待培養12 d后菌落表面完全覆蓋分生孢子,用直徑5 mm的打孔器在菌落半徑上距離菌落中心1/2處打取菌餅,每菌落按十字對稱取4個菌餅,置于定量的0.1%的Tween-80無菌水中,振蕩使孢子充分分散。用血球計數板測定孢子濃度并換算成單位面積(cm2)的產孢量,每皿血球計數板計數3次。
1.6 mad2敲除株的毒力測定
將4齡初的家蠶隨機分配至無菌的生測筐中(長×寬×高=30 cm×12 cm×9 cm),每框30頭,每個處理5個重復。分別收集同一時間培養的野生株與敲除株孢子粉,懸浮于無菌的0.05%的Tween-80水中,顯微計數后將孢子濃度調整至5×108孢子/mL。將噴霧塔(Burkard Potter Precision Laboratory)氣壓調整至80 kpa,用0.05%的吐溫水清洗3次,每次5 mL。每框試蟲噴菌4 mL,空白對照(Control-0.05% Tween)噴4 mL無菌的0.05%的Tween-80。30 min后,每框加入桑葉20 g。每天記錄家蠶死亡數,并補充足量桑葉,連續記錄11 d,計算每天死亡率,并繪制死亡率柱狀圖。
1.7 mad2敲除株誘導花生共生基因響應分析
花生種子表面消毒參照Mayo等[25]的方法,于70%乙醇中浸泡1 min,無菌水沖洗3次,再用1%次氯酸鈉溶液浸泡15 min,無菌水沖洗3次,完成消毒。將消毒后的種子置于含有50 mL ddH2O的直徑為15 cm的培養皿中,并置于28℃黑暗培養箱中催芽24 h,挑選健康且萌芽一致的種子播種于裝滿滅菌蛭石的直徑15 cm的花盆中,于培養室(L﹕D=14 h﹕10 h,28℃)培養,相對濕度為40%—60%。于花生生長第14天,分別向花生苗根部滴灌野生株與敲除株的分生孢子懸浮液(濃度為1.0×108孢子/mL),每株滴灌20 mL,對照(Control-0.1% Tween)為向植株根部滴灌0.1% Tween-80水溶液20 mL,每個處理重復3株。處理12 h后小心將根取出,將根部蛭石用蒸餾水沖洗去除干凈蛭石基質,用吸水紙吸干表面,快速將根剪成5—8 mm小段,立即置于液氮中。同一處理不同的重復進行混樣并分成3份,保存于-80℃備用。
取花生根樣品,置于液氮中研磨,然后用TRIzol?分離試劑盒(Invitrogen)提取總RNA。用Prime ScriptTM1st strand cDNA synthesis kit反轉錄試劑盒(TaKaRa公司)進行RNA反轉錄,得到cDNA。并用NanoPhotometer微量分光光度計(IMPLEN,德國)檢測濃度。
基于之前的轉錄組分析確定候選基因,設計的引物序列見表2,內參基因選用60S ribosomal protein L7()。用SYBR Green熒光定量試劑盒(TaKaRa公司)在7500型(ABI公司)實時熒光定量PCR系統進行反應。反應體系為2×SYBR Green Master Mix 10 μL,正反向引物各1 μL,cDNA 0.8 μL,補足ddH2O至20 μL。PCR擴增程序為95℃預變性2 min;95℃變性5 s,60℃復性30 s,40個循環。每樣品3次技術重復,基因相對表達量采用2-ΔΔCt的方法進行分析。
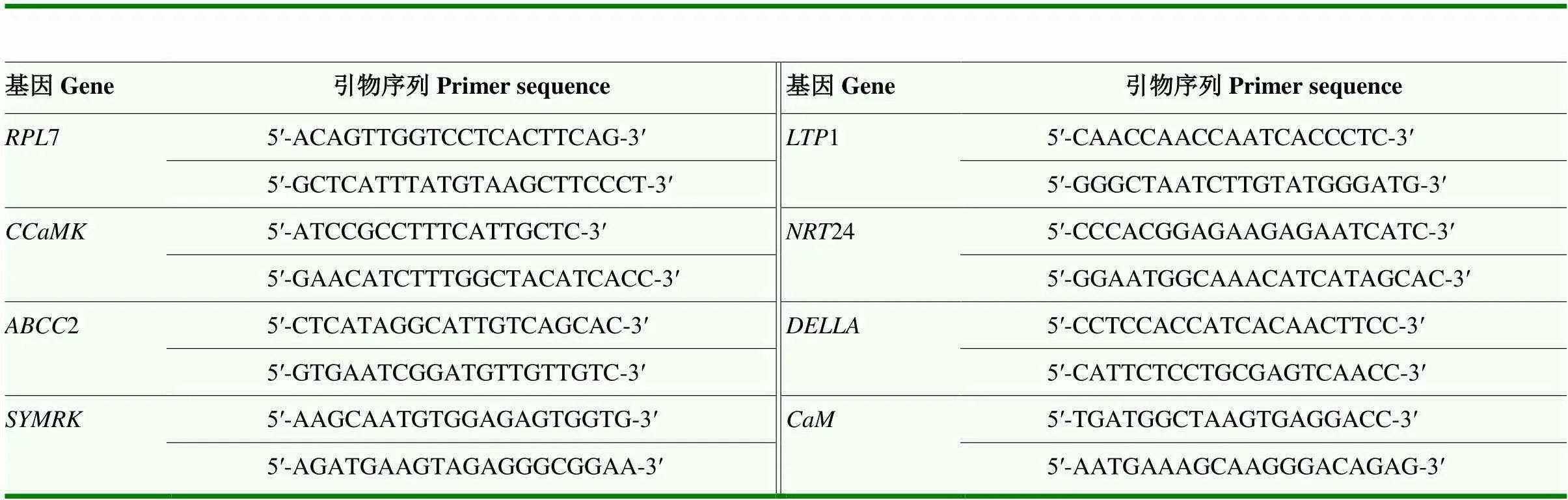
表2 候選基因及其特異性引物
1.8 數據處理
萌發率、生長速率以及產孢量的試驗數據利用SPSS 20.0進行分析,數據差異采用單因素-test進行比較。
家蠶死亡率(%)=家蠶死亡數/家蠶總數×100,死亡率統計結果采用SPSS 20.0軟件進行分析,用Duncan test進行多重比較。分析結果采用Graph Pad Prism 6軟件進行圖表制作。
花生中各基因表達試驗數據利用SPSS 20.0進行分析,用Duncan test進行多重比較。分析結果采用GraphPad Prism 6軟件進行圖表繪制。
2 結果
2.1 綠僵菌黏附素基因mad2敲除株的構建
2.1.12前后同源臂克隆 利用特異性引物mad2 S1-F/R擴增得到了1 046 bp的2同源前臂基因序列S1,利用引物mad2 S2-F/R擴增得到了1 174 bp的同源后臂基因序列S2,利用潮霉素引物Hyg-F/R擴增得到了含有Ptrpc啟動子的hygromycin潮霉素基因片段,共1 426 bp(圖2)。
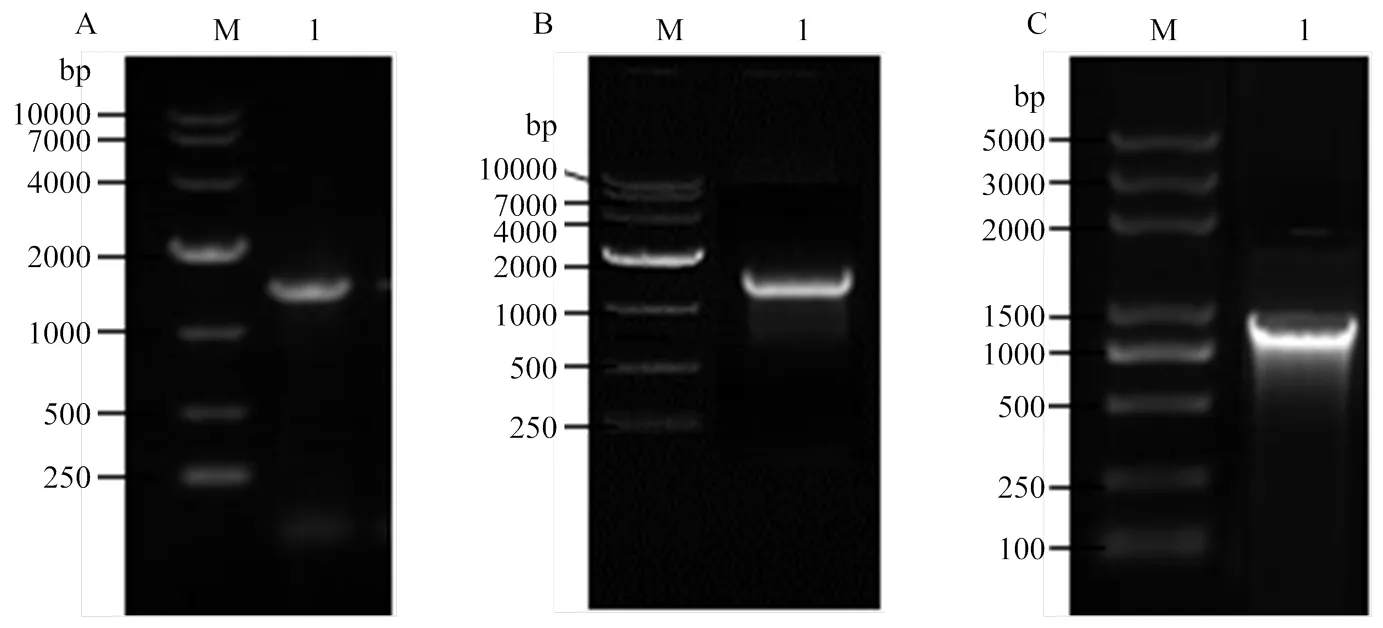
M: DNA marker; A-1: Hyg; B-1: S1; C-1: S2
通過overlap PCR,利用mad2 S1-F與Hyg S1-R引物將1 046 bp S1基因與945 bp潮霉素前端序列相連,獲得S1H;利用Hyg S2-F與mad2 S2-R 1 174 bp S2與潮霉素后936 bp序列相連,獲得S2H序列(圖3)。S1H 片段與S2H片段中間重疊部分的長度為455 bp(圖1)。
2.1.2 同源臂轉化綠僵菌及轉化子篩選 通過CaC12-PEG介導,將擴增得到的2基因同源臂敲除盒S1H、S2H片段轉化綠僵菌原生質體,原生質體再生后獲得了轉化子。轉化子在潮霉素平板上轉接3代,最終獲得10個穩定的轉化子。通過特異性引物Out-mad2-F/R和潮霉素Hyg-F/R分別對轉化子進行PCR檢測,顯示有5個轉化子的2基因片段擴增為陰性(圖4中2—5、9泳道),且同時潮霉素片段擴增為陽性(圖4中對應12—15、19泳道),說明這些轉化子已經成功敲除2,并在該基因位置處插入了潮霉素片段。選用第9號泳道轉化子進行后續研究。
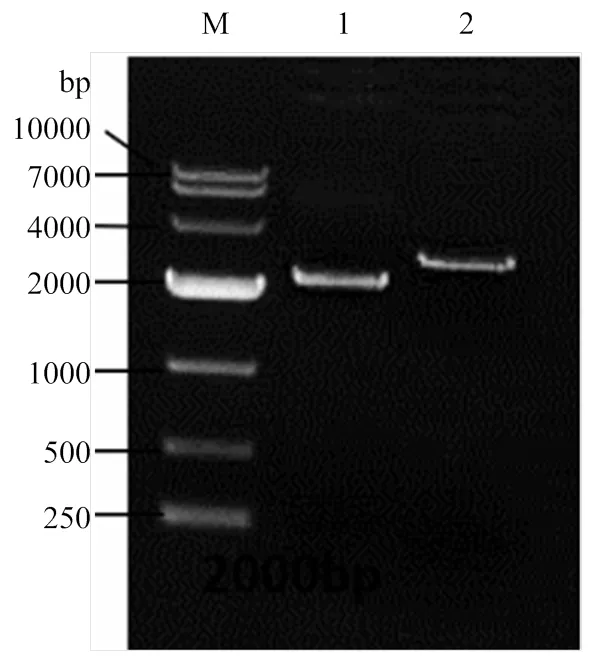
M: DL10000 DNA marker; 1: S1H; 2: S2H
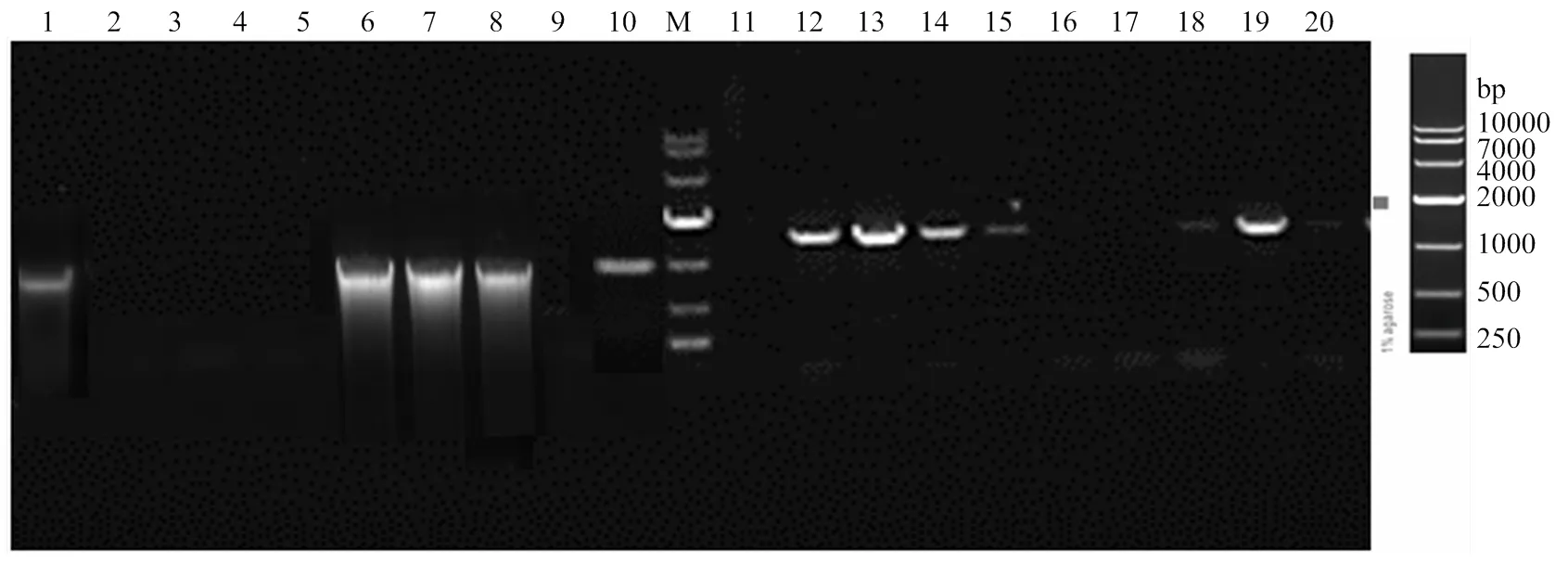
1—10:10個轉化子OUT-mad2-F/R引物擴增結果 PCR amplification from 10 transformants by primer OUT-mad2-F/R;11—20為10個轉化子Hyg-F/R引物擴增結果 PCR amplification from 10 transformants by primer Hyg-F/R primer
2.2 敲除mad2對菌株黏附力的影響
顯微鏡下觀察了綠僵菌野生株和敲除株孢子在洋蔥內表皮和蝗蟲后翅上的附著情況(圖5-A),顯示在處理后8 h,敲除株的分生孢子對洋蔥內表皮的吸附量顯著低于野生株,但二者對于蝗蟲后翅表皮的吸附量并無明顯差異(圖5-B),說明綠僵菌2敲除之后,降低了孢子對植物的黏附力,即黏附素MAD2有利于綠僵菌在植物上的黏附,但不影響對昆蟲的附著。
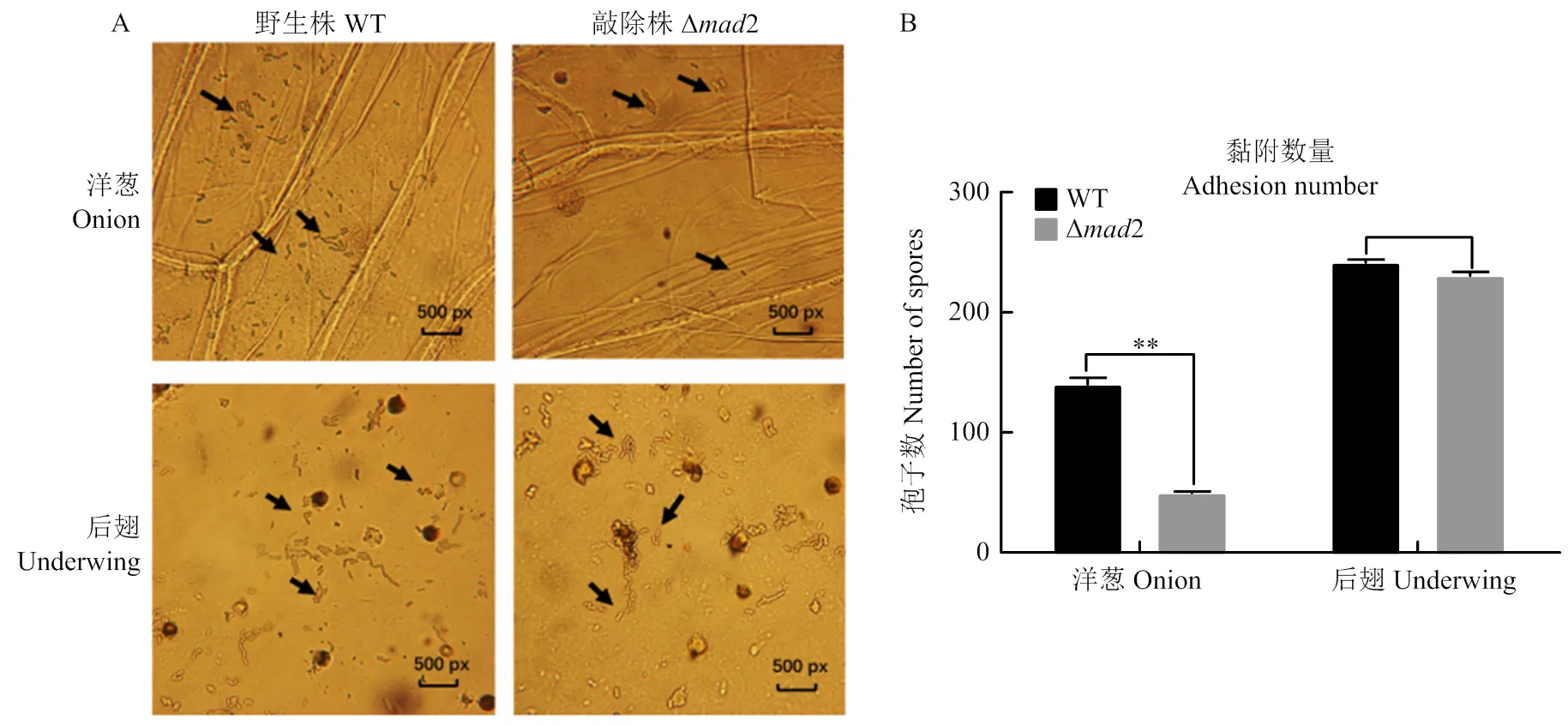
A:光學顯微鏡10×20倍,箭頭所指為孢子adherent spores (pointed by arrows) by 10×20 light microscopy;B:**:相對于野生株有極顯著差異there is an extremely significant difference compared with WT (P<0.01)
2.3 敲除mad2對菌株生長的影響
敲除株的萌發率在6、10、14、18、24 h均顯著低于野生株,在培養36 h時,萌發率無顯著差異(圖6-A)。野生株的萌發中時為7.98 h,而敲除株的萌發中時為13.45 h,顯著長于野生株,表明2的缺失抑制孢子的萌發。在12、14 h時,敲除株的菌絲長度顯著短于野生株,分別短22.2%、23.7%(圖6-B),說明2有利于綠僵菌孢子的萌發和生長。對于產孢量,在培養第12天,野生株產孢量為(5.22±1.01)×108孢子/cm2,而敲除株產孢量為(3.48±0.17)×108孢子/cm2,比野生株顯著下降了33.3%,表明2的缺失影響了綠僵菌的產孢(圖6-C)。
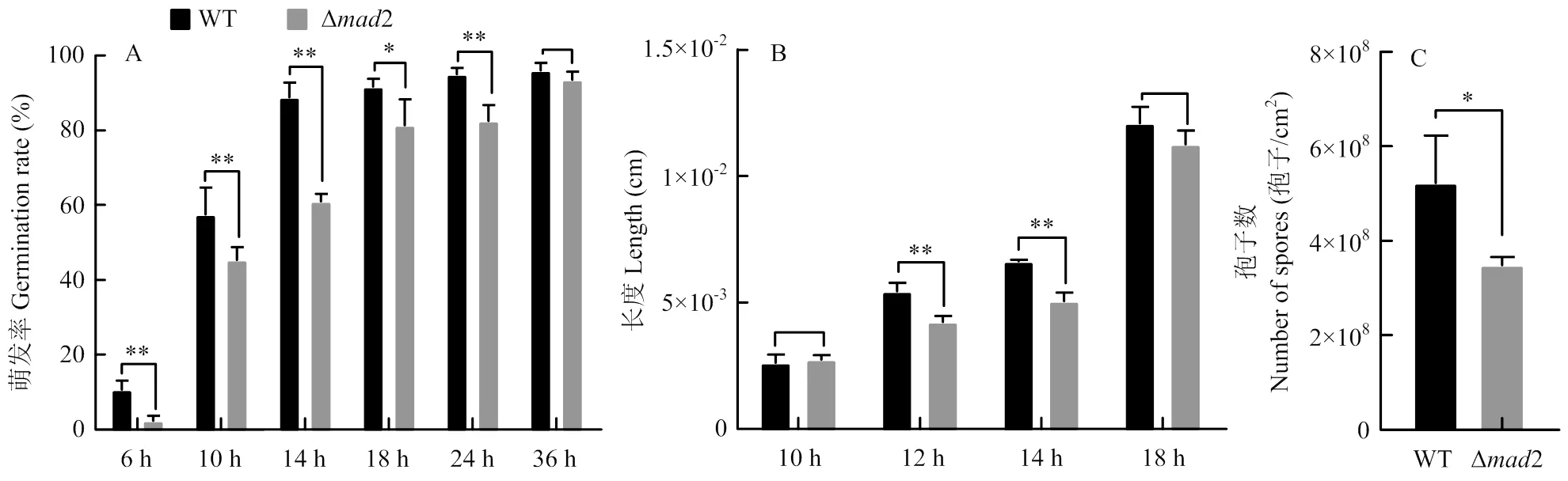
**:相對于野生株有極顯著差異there is an extremely significant difference compared with WT (P<0.01);*:相對于野生株有顯著差異there is a significant difference compared with WT (P<0.05)
2.4 敲除mad2對菌株毒力的影響
敲除株、野生株對家蠶均有較高毒力,與空白對照相比,在第6天的死亡率分別為9.34%和7.32%,均達到差異顯著水平,之后兩者處理的死亡率逐日上升,但一直無顯著差異,最終死亡率均達到80%以上(圖7),表明2的敲除并不影響綠僵菌的毒力。
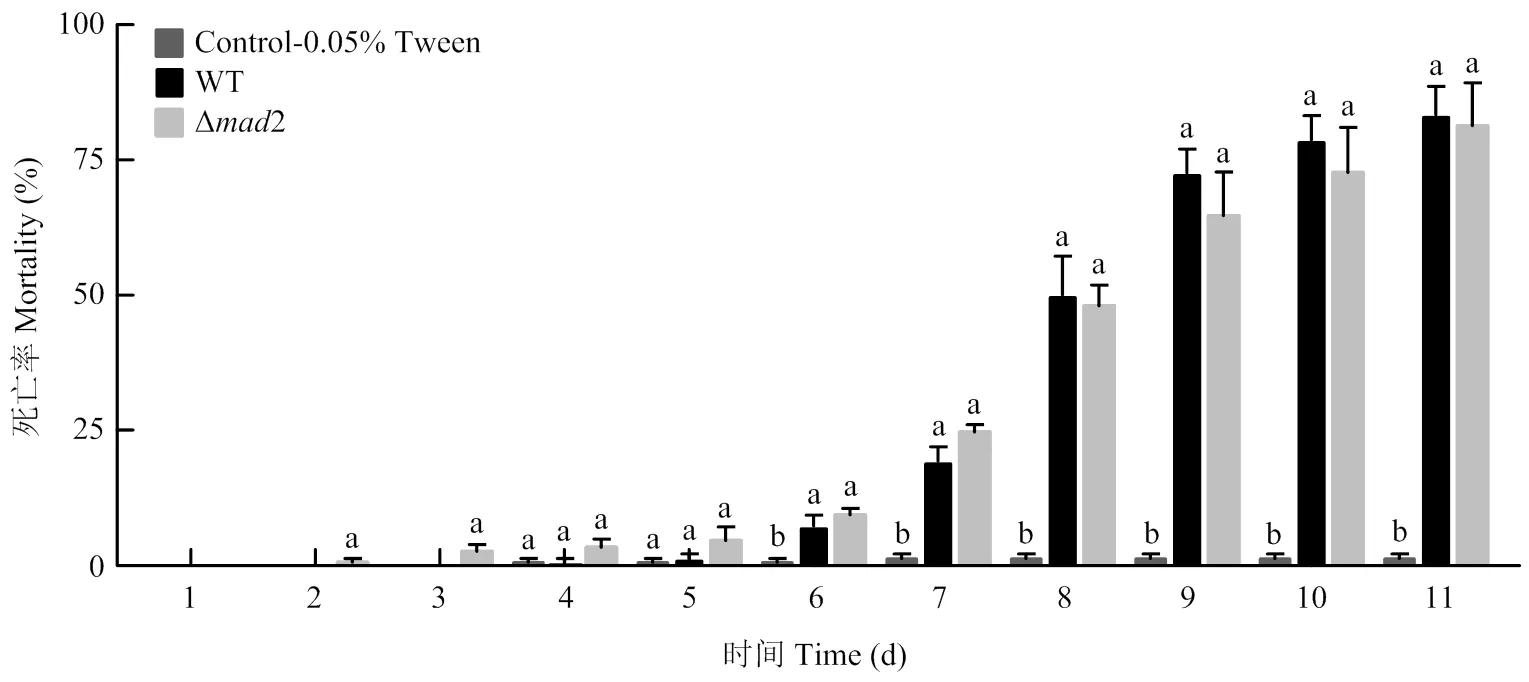
柱上不同字母代表不同處理間在0.05水平差異顯著Different letters on the column represent significant differences among different treatments at P<0.05 level。圖8同The same as Fig. 8
2.5 敲除株與野生株誘導下花生共生基因差異響應
檢測了綠僵菌野生株、敲除株處理花生根的7個共生相關基因轉錄水平,發現在處理12 h后,敲除株處理下花生根共生受體的轉錄水平顯著下降至野生株處理下的68.4%;同時,鈣調素蛋白基因、鈣-鈣調素依賴蛋白激酶基因、信號解碼的蛋白基因、ABC家族轉運蛋白基因2、脂質轉移蛋白基因1以及硝酸根轉運蛋白基因24的轉錄水平相較于野生株處理均顯著下降,其中和1分別下降至野生株的15.6%和10.8%,且降低到空白對照的16.5%。但相較于空白對照,2的敲除只對、和的轉錄水平產生顯著抑制,而對有一定的上調作用,對2、1、24的轉錄無明顯影響(圖8)。由此可見,2敲除后綠僵菌對共生相關基因的誘導表達減弱,說明綠僵菌的MAD2蛋白參與了觸發共生相關基因的轉錄,而不同基因受到的調控方式和調控程度不同。
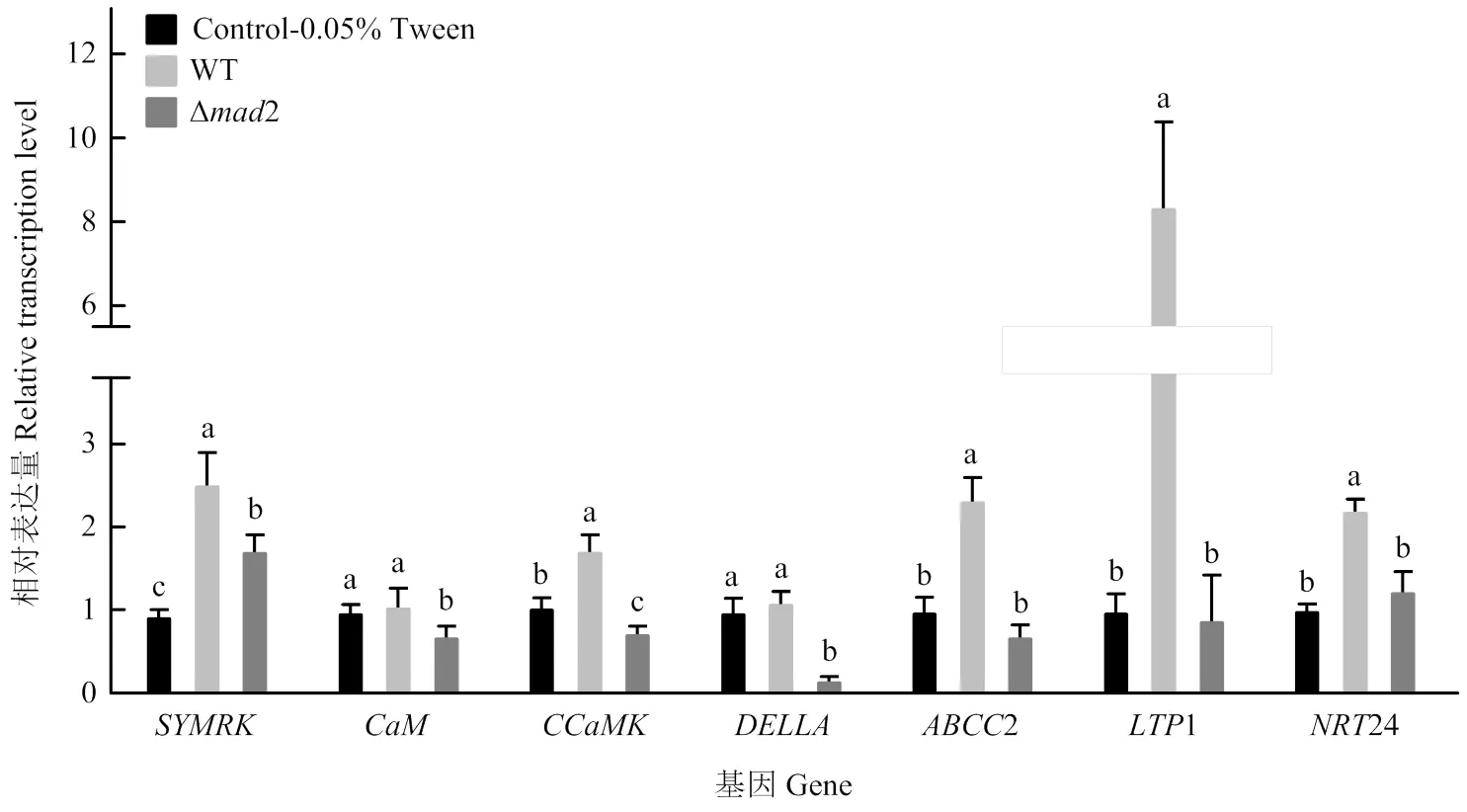
圖8 綠僵菌野生株與敲除株處理下花生根共生基因實時熒光定量PCR檢測
3 討論
3.1 黏附素MAD2影響綠僵菌生長和對植物的識別與附著
已報道黏附素MAD2與綠僵菌黏附植物有關,但其作用機理尚不明確。黏附是微生物侵入宿主過程中的第一步,黏附素分子是微生物與宿主識別、信息交流的關鍵因子。目前,白色念珠菌黏附素的特性和作用研究較清楚,該黏附素多肽由類凝集素序列(ALS)、菌絲胞壁蛋白1(hyphal wall protein 1,Hwp1)以及整合素基因(Int 1p)等基因編碼。這些基因與細胞間的識別、菌絲的形成以及毒力相關[26]。類凝集素ALS參與白色念珠菌黏附功能,尤其是ALS3蛋白對黏附作用至關重要,缺失會使真菌黏附于上皮細胞的能力明顯降低[27]。AlS3通過促進與宿主內生菌交聯形成混合生物膜,誘導宿主細胞內吞而產生黏附作用和介導入侵。例如,ALS3能與口腔格氏鏈球菌()的單鏈DNA結合蛋白(single strand DNA-binding protein,Ssb)交互作用,通過菌絲與體內鏈球菌相連接形成混合生物膜,促進白色念珠菌在體內穩定定殖[28]。在與宿主細胞表面E-鈣黏蛋白和N-鈣黏蛋白綁定后,ALS3能夠誘導上皮細胞內吞作用而促進白色念珠菌入侵[29]。而白色念珠菌Hwpl蛋白具有頰上皮細胞谷氨酰胺轉胺酶(transglutaminase,TGase)活性,與頰上皮細胞的蛋白質交聯,當敲除后就會降低對底物TGase的作用活性,因而降低與人頰黏膜上皮細胞的共價結合水平[30]。在敲除后,白色念珠菌毒力降低,對上皮細胞的附著減少40%,并且造成菌絲在奶粉-吐溫培養基和蜘蛛(spider)培養基上生長有缺陷[31]。綠僵菌黏附素MAD2與白色念珠菌胞壁凝集素分子AlS的結構十分相似,也由3個主要結構域組成,在其N-端有疏水信號肽,C-端具有預測的糖基磷脂酰肌醇細胞壁錨點,中間區域含有富含Thr的串聯重復序列,可通過快速形成穩定的H鍵與宿主蛋白和肽相結合[7]。Fu等[32]發現白色念珠菌Als 1p除了是侵染小鼠的毒力因子,還是菌株表皮生長因子1(efg1)的效應子,調節菌絲形成途徑。本研究發現綠僵菌黏附素MAD2也具有影響菌體生長和附著能力的作用。當缺失2后,孢子萌發率降低,菌絲生長率變慢,產孢量減少,對洋蔥表皮的黏附力下降,但對蝗蟲后翅的黏附性和對家蠶的毒力沒有影響,說明黏附素MAD2參與了對植物的附著,并涉及孢子萌發、菌絲生長和產孢,從而影響綠僵菌與植物之間共生關系的建立。
3.2 MAD2與綠僵菌非適條件的適應有關
自然界中,綠僵菌通過侵染各種昆蟲寄主獲得大量繁殖以擴增種群,昆蟲及其棲境提供了綠僵菌的適宜營養和生存條件。有研究表明,在昆蟲表皮培養基上,綠僵菌孢子快速萌發和生長,但此過程2的轉錄并無顯著變化,而在大豆根分泌液或水培上,綠僵菌緩慢生長,2轉錄從處理后2 h即開始積累,并伴隨菌絲生長逐漸增加,但在培養20 d的綠僵菌孢子粉中,2的拷貝數極低,說明MAD2參與綠僵菌與植物的早期互作[7]。相對的,2在非適環境條件下(如營養缺乏、甲殼素處理、番茄莖稈上)顯示上調表達,是一種抗逆進化的表現[33]。本研究中也發現,在水瓊脂(營養缺失)培養中,2敲除株在培養6、10、14、18、24 h各時間段的萌發率均顯著低于野生株,說明MAD2涉及綠僵菌在非適條件下的生長。
3.3 綠僵菌MAD2參與誘導植物SYMRK響應以調節植物生長和促進共生
植物體內鈣離子信號途徑涉及多種生理機能,包括與環境微生物共生的調控。在與叢枝菌根真菌的典型共生中,植物(共生受體激酶)接收外界信號因子,引起Ca2+的內流,胞內Ca2+激增,Ca2+作為二級信號向胞內傳遞,(鈣調素蛋白)具有保守的Ca2+結合結構域,在共生過程中具有信號轉導的功能[34-35]。(鈣-鈣調素依賴蛋白激酶)在鈣存在的情況下,Ca2+-與結合,促進構象變化,誘導底物蛋白磷酸化。磷酸化的與形成復合物,與GRAS轉錄因子(如DELLA蛋白)協同作用,啟動下游基因的表達[36]。前期研究表明,綠僵菌可以促進植物共生基因的轉錄[37]。本研究中,缺失2后,在野生株處理顯著上調的轉錄水平出現一定程度下降,而2缺失使鈣信號解碼的基因、、轉錄水平較野生株或空白對照均顯著下調,說明感應了MAD2及其他信號因子,被誘導上調表達,通過Ca2+信號轉導,引起響應,并使的轉錄受到顯著抑制。不僅是赤霉素信號通路的阻遏因子,還能通過與(依賴鈣離子蛋白激酶IMD3的底物)結合形成蛋白復合體,促進豆科植物原生質體中結核誘導相關基因的表達,進而促進根瘤菌共生[38]。2缺失對轉錄的強烈抑制說明MAD2在綠僵菌誘導植物生長調節、促進共生建立中具有重要作用。
3.4 綠僵菌MAD2可調動植物脂質和氮素轉運功能以參與共生建立
共生的后期階段涉及多種物質的合成與轉運,許多基因參與共生建立。2屬于ABC(ATP-binding cassette)轉運蛋白家族,參與植物體內激素、脂質、金屬離子、次生代謝物和外源物質的運輸[39],1(脂質轉移蛋白)被認為在胞間發生作用,負責脂質結合和轉運,參與植物脂質動員,角質層合成[40],24(硝酸根轉運蛋白)參與硝態氮的吸收,并且在大豆中發現,過表達或后大豆根瘤數目顯著增加[41]。在本研究中,野生株處理時對2124的轉錄上調至空白對照的2.34、8.32、2.17倍,而2缺失后相較于空白對照無明顯差異,說明MAD2是誘導2124上調的關鍵因子,并可能通過獨立途徑實現。可見,MAD2可調動宿主的轉運脂質和氮素功能,參與綠僵菌與植物共生的建立。
4 結論
成功構建了綠僵菌敲除2的穩定突變株。敲除株生物學分析表明,黏附素MAD2可正向調控綠僵菌的萌發、菌絲生長、產孢以及對植物的附著能力,但不影響綠僵菌的殺蟲毒力。MAD2被植物受體識別后,調節植物中共生基因(、、、)的轉錄,誘導植物脂質和氮素合成和轉運基因(2124)的轉錄上升。綜合顯示,黏附素MAD2與綠僵菌的生長、黏附和環境適應性有關,并且誘導與植物共生信號的交流,啟動對植物的附著和共生關系的建立。
[1] MENG Y, ZHANG X, GUO N, FANG W. Mrst12 implicated in the regulation of transcription factor aftf1 by fus3-mapk during cuticle penetration by the entomopathogenic fungus. Fungal Genetics and Biology, 2019, 131: 103244. DOI: 10.1016/j.fgb.2019.103244.
[2] PARSA S, ORTIZ V, GóMEZ-JIMéNEZ M I, KRAMER M, VEGA F E. Root environment is a key determinant of fungal entomopathogen
endophytism following seed treatment in the common bean,. Biological Control, 2018, 116: 74-81. DOI:10.1016/j.biocontrol.2016.09.001.
[3] STEINWENDER B M, ENKERLI J, WIDMER F, EILENBERG J, KRISTENSEN H L, BIDOCHKA M J, MEYLING N V. Root isolations ofspp. from crops reflect diversity in the soil and indicate no plant specificity. Journal of Invertebrate Pathology, 2015, 132: 142-148. DOI: 10.1016/j.jip.2015.09.007.
[4] RAYA-DíAZ S, SáNCHEZ-RODRíGUEZ A R, SEGURA-FERNáNDEZ J M, DEL CARMEN DEL CAMPILLO M, QUESADA-MORAGA E. Entomopathogenic fungi-based mechanisms for improved Fe nutrition in sorghum plants grown on calcareous substrates. Plos One, 2017, 12(10): e0185903. DOI: 10.1371/journal.pone.0185903.
[5] KHAN A L, HAMAYUN M, KANG S M, KIM Y H, JUNG H Y, LEE J H, LEE I J. Endophytic fungal association via gibberellins and indole acetic acid can improve plant growth under abiotic stress: an example oflhl10. BMC Microbiology, 2012, 12: 3. DOI: 10.1186/1471-2180-12-3.
[6] AKELLO J, SIKORA R. Systemic acropedal influence of endophyte seed treatment onandoffspring development and reproductive fitness. Biological Control, 2012, 61(3): 215-221. DOI:10.1016/j.biocontrol.2012.02.007.
[7] WANG C, ST LEGER R J. The MAD1 adhesin oflinks adhesion with blastospore production and virulence to insects, and the MAD2 adhesin enables attachment to plants. Eukaryotic Cell, 2007, 6(5): 808-816. DOI: 10.1128/EC.00409-06.
[8] NICHOLSON R L, EPSTEIN L. Adhesion of fungi to the plant surface//COLE G T. The Fungal Spore and Disease Initiation in Plants and Animals. Springer US, 1991: 3-23. DOI: https://doi.org/ 10.1007/978-1-4899-2635-7_1.
[9] SHEPPARD D C, YEAMAN M R, WELCH W H, PHAN Q T, FU Y, IBRAHIM A S, FILLER S G, ZHANG M, WARING A J, JR EDWARDS J E. Functional and structural diversity in the Als protein family of. The Journal of Biological Chemistry, 2004, 279(29): 30480-30489. DOI: 10.1074/jbc.M401929200.
[10] PAVA-RIPOLL M, ANGELINI C, FANG W, WANG S, POSADA F J, ST LEGER R. The rhizosphere-competent entomopathogenexpresses a specific subset of genes in plant root exudate. Microbiology, 2011, 157(1): 47-55. DOI: 10.1099/mic. 0.042200-0.
[11] 閆多子, 蔡霓, 王峰, 農向群, 王廣君, 涂雄兵, 張澤華. 綠僵菌黏附素MAD1體外表達及誘導花生響應的作用. 中國農業科學, 2021, 54(4): 744-753. DOI: 10.3864/j.issn.0578-1752.2021.04.007.
YAN D Z, CAI N, WANG F, NONG X Q, WANG G J, TU X B, ZHANG Z H. Expression in vitro ofadhesin MAD1 and its effect on inducing response in peanut. Scientia Agricultura Sinica, 2021, 54(4): 744-753. DOI: 10.3864/j.issn.0578- 1752.2021.04.007. (in Chinese)
[12] KRELL V, UNGER S, JAKOBS-SCH?NWANDT D, PATEL A V. Endophyticmitigates nutrient deficits in potato and improves plant productivity and vitality. Fungal Ecology, 2018, 34: 43-49. DOI: 10.1016/j.funeco.2018.04.002.
[13] BEHIE S W, BIDOCHKA M J. Ubiquity of insect-derived nitrogen transfer to plants by endophytic insect-pathogenic fungi: an additional branch of the soil nitrogen cycle. Applied and Environmental Microbiology, 2014, 80(5): 1553-1560. DOI: 10.1128/AEM. 03338-13.
[14] BEHIE S W, MOREIRA C C, SEMENTCHOUKOVA I, BARELLI L, ZELISKO P M, BIDOCHKA M J. Carbon translocation from a plant to an insect-pathogenic endophytic fungus. Nature Communications, 2017, 8: 14245. DOI: 10.1038/ncomms14245.
[15] BEHIE S W, ZELISKO P M, BIDOCHKA M J. Endophytic insect-parasitic fungi translocate nitrogen directly from insects to plants. Science, 2012, 336(6088): 1576-1577. DOI: 10.1126/science. 1222289.
[16] SASAN R K, BIDOCHKA M J. The insect-pathogenic fungus(clavicipitaceae) is also an endophyte that stimulates plant root development. American Journal of Botany, 2012, 99(1): 101-107. DOI: 10.3732/ajb.1100136.
[17] LIAO X, O’BRIEN T R, FANG W, ST LEGER R J. The plant beneficial effects ofspecies correlate with their association with roots. Applied Microbiology and Biotechnology, 2014, 98(16): 7089-7096. DOI: 10.1007/s00253-014-5788-2.
[18] BEHIE S W, JONES S J, BIDOCHKA M J. Plant tissue localization of the endophytic insect pathogenic fungiand. Fungal Ecology, 2015, 13: 112-119. DOI: 10.1016/j.funeco.2014. 08.001.
[19] CAI N, WANG F, NONG X Q, WANG G J, McNeill M, Cao G C, Hao K, Liu S F,ZHANG Z H. Visualising confirmation of the endophytic relationship ofwith maize roots using molecular tools and fluorescent labelling. Biocontrol Science and Technology, 2019, 29(11): 1023-1036. DOI: 10.1080/09583157. 2019.1641792.
[20] HAO K, WANG F, NONG X Q, MCNEILL M R, LIU S F, WANG G J, CAO G C, ZHANG Z H. Response of peanutroots to the presence of beneficial and pathogenic fungi by transcriptome analysis. Scientific Reports, 2017, 7(1): 964. DOI: 10.1038/s41598-017-01029-3.
[21] 徐芳, 姚泉洪, 熊愛生, 彭日荷, 侯喜林, 曹兵. 重疊延伸pcr技術及其在基因工程上的應用. 分子植物育種, 2006, 4(5): 747-750. DOI: 10.3969/j.issn.1672-416X.2006.05.025.
XU F, YAO Q H, XIONG A S, PENG R H, HOU X L, CAO B. SOE PCR and its application in genetic engineering. Molecular Plant Breeding, 2006, 4(5): 747-750. DOI: 10.3969/j.issn.1672-416X.2006. 05.025. (in Chinese)
[22] 王曉玲, 蔣伶活, 張澤華. 以G418抗性為選擇標記的金龜子綠僵菌原生質體轉化的研究. 安徽農業科學, 2007, 35(28): 8865-8866, 8868. DOI: 10.3969/j.issn.0517-6611.2007.28.020.
WANG X L, JIANG L H, ZHANG Z H. Protoplast transformation ofwith G418 resistance as selective marker. Journal of Anhui Agricultural Sciences, 2007, 35(28): 8865-8866, 8868. DOI: 10.3969/j.issn.0517-6611.2007.28.020. (in Chinese)
[23] 劉艷微. 轉基因球孢白僵菌工程菌株的構建及鑒定[D]. 哈爾濱: 哈爾濱師范大學, 2015. DOI: 10.7666/d.D678481.
LIU Y W. Construction and identification oftransgenicstrains[D]. harbin: harbin Normal University, 2015. DOI: 10.7666/d.D678481. (in Chinese)
[24] 蔡守平, 何學友, 曾麗瓊, 黃金水, 鐘景輝, 嵇保中. 感染星天牛幼蟲高致病力金龜子綠僵菌菌株的篩選. 中國生物防治學報, 2012, 28(2): 293-297. DOI: 10.3969/j.issn.2095-039X.2012.02.023.
CAI S P, HE X Y, ZENG L Q, HUANG J S, ZHONG J H, JI B Z. Susceptibility oflarvae to the entomopathogenic fungus,Chinese Journal of Biological Control, 2012, 28(2): 293-297. DOI: 10.3969/j.issn.2095-039X.2012.02.023. (in Chinese)
[25] MAYO S, COMINELLI E, SPARVOLI F, GONZáLEZ-LóPEZ O, RODRíGUEZ-GONZáLEZ A, GUTIéRREZ S, CASQUERO P A. Development of a qPCR strategy to select bean genes involved in plant defense response and regulated by theinteraction.Frontiers in Plant Science, 2016, 7: 1109. DOI: 10.3389/fpls.2016.01109.
[26] 廉翠紅, 劉維達. 與白念珠菌黏附和侵入相關的毒力因子研究進展. 國際皮膚性病學雜志, 2004, 30(5): 323-325. DOI: 10.3760/cma.j.issn.1673-4173.2004.05.018.
LIAN C H, LIU W D. Research progress on virulence factors related to adhesion and invasion ofInternational Journal of Dermatology and Venereology, 2004, 30(5): 323-325. DOI: 10.3760/cma.j.issn.1673-4173.2004.05.018. (in Chinese)
[27] W?CHTLER B, CITIULO F, JABLONOWSKI N, F?RSTER S, DALLE F, SCHALLER M, WILSON D, HUBE B.-epithelial interactions: dissecting the roles of active penetration, induced endocytosis and host factors on the infection process. PLoS ONE, 2012, 7(5): e36952. DOI: 10.1371/journal.pone. 0036952.
[28] SILVERMAN R J, NOBBS A H, VICKERMAN M M, BARBOUR M E, JENKINSON H F. Interaction ofcell wall als3 protein withSspb adhesin promotes development of mixed-species communities. Infection and Immunity, 2010, 78(11): 4644-4652. DOI: 10.1128/IAI.00685-10.
[29] MURCIANO C A, MOYES D L, RUNGLALL M, TOBOUTI P, ISLAM A, HOYER L L, NAGLIK J R. Evaluation of the role ofagglutinin-like sequence (Als) proteins in human oral epithelial cell interactions.PLoS ONE, 2012, 7(3): e33362. DOI: 10.1371/journal.pone.0033362.
[30] STAAB J F,Bradway S D, Fidel P L, Sundstrom P. Adhesive and mammalian transglutaminase substrate properties ofhwp1. Science, 1999, 283(5407): 1535-1538. DOI: 10.1126/science.283.5407.1535.
[31] GALE C A, BENDEL C M, MCCLELLAN M, HAUSER M, BECKER J M, BERMAN J, HOSTETTER M K. Linkage of adhesion, filamentous growth, and virulence into a single gene,. Science, 1998, 279(5355): 1355-1358. DOI:10.1126/ science.279.5355.1355.
[32] FU Y, IBRAHIM A S, SHEPPARD D C, CHEN Y C, FRENCH S W, CUTLER J E, FILLER S G, EDWARDS J E.ALS1p: an adhesin that is a downstream effector of thefilamentation pathway. Molecular Microbiology, 2002, 44(1): 61-72. DOI: 10.1046/j.1365-2958.2002.02873.x.
[33] BARELLI L, PADILLA-GUERRERO I E, BIDOCHKA M J. Differential expression of insect and plant specific adhesin genes,and, in. Fungal Biology, 2011, 115(11): 1174-1185. DOI: 10.1016/j.funbio.2011.08.003.
[34] BATISTI O, KUDLA J. Analysis of calcium signaling pathways in plants. Biochimica et biophysica acta, 2012, 1820(8): 1283-1293. DOI: 10.1016/j.bbagen.2011.10.012.
[35] 熊珊珊, 趙斌.叢枝菌根真菌的鈣調素基因在共生過程中的作用. 湖北農業科學, 2014, 53(13): 3177-3182. DOI: 10.3969/j.issn.0439- 8114.2014.13.052.
XIONG S S, ZHAO B. The roles of calmodulin gene ofin symbiosis process. Hubei Agricultural Sciences, 2014, 53(13): 3177-3182. DOI: 10.3969/j.issn.0439-8114.2014.13.052. (in Chinese)
[36] MACLEAN A M, BRAVO A, HARRISON M J. Plant signaling and metabolic pathways enabling arbuscular mycorrhizal symbiosis. The Plant Cell, 2017, 29(10): 2319-2335. DOI: 10.1105/tpc.17.00555.
[37] WANG F, NONG X Q, HAO K, CAI N, WANG G J, LIU S F, ULLAH H, ZHANG Z H. Identification of the key genes involved in the regulation of symbiotic pathways induced byin peanut () roots. 3 Biotech, 2020, 10(3): 124. DOI: 10.1007/s13205-020-2105-x.
[38] JIN Y, LIU H, LUO D, YU N, DONG W, WANG C, ZHANG X, DAI H, YANG J, WANG E. Della proteins are common components of symbiotic rhizobial and mycorrhizal signalling pathways. Nature Communications, 2016, 7: 12433. DOI: 10.1038/ncomms12433.
[39] 邵若玄, 沈憶珂, 周文彬, 方佳, 鄭炳松. 植物ATP結合盒(abc)轉運蛋白研究進展. 浙江農林大學學報, 2013, 30(5): 761-768. DOI: 10.11833/j.issn.2095-0756.2013.05.020.
SHAO R X, SHEN Y K, ZHOU W B, FANG J, ZHENG B S. Recent advances for plant ATP-binding cassette transporters.Journal of Zhejiang A&F University, 2013, 30(5): 761-768. DOI: 10.11833/j.issn. 2095-0756.2013.05.020. (in Chinese)
[40] CARVALHO A O, GOMES V M. Role of plant lipid transfer proteins in plant cell physiology: a concise review. Peptides, 2007, 28: 1144-1153. DOI: 10.1016/j.peptides.2007.03.004.
[41] 李國紀, 朱林, 曹金山, 王幼寧. 大豆和基因的克隆及功能探究. 作物學報, 2020, 46(7): 1025-1032. DOI: 10.3724/SP.J.1006.2020.94152.
LI G J, ZHU L, CAO J S, WANG Y N. Cloning and functional analysis ofandin soybean. Acta Agronomica Sinica, 2020, 46(7): 1025-1032. DOI: 10.3724/SP.J.1006.2020.94152. (in Chinese)
Adhesin gene2 knockout and functional effects on Biological characteristics and inducing plant responses in
CAI Ni, YAN DuoZi, NONG XiangQun, WANG GuangJun, TU XiongBing, ZHANG ZeHua
State Key Laboratory for Biology of Plant Diseases and Insect Pests, Institute of Plant Protection, Chinese Academy of Agricultural Sciences, Beijing 100193
【Objective】, an entomopathogenic fungus, is found also an endophyte. MAD2 is known to be one of the two adhesin proteins of, which plays a vital role in adhesion and colonization in plants, but its functional mechanism is poorly understood. The objective of this study is to explore the functional effect of MAD2 protein on thecharacteristics of growth, virulence, adhesion and inducing plant responses inby construction of the2 mutant strain (Δ2) ofstrain Ma9. 【Method】The genomic DNA sequences of2 anteroposterior were obtained from NCBI, and specific primers were designed to amplify2 homologous arm genes S1 and S2 by PCR based on genomic DNA template ofstrain Ma9. Meanwhile, Hyg-F/R primer pair was designed to amplify hygromycin gene with promoter sequence based on pKH-KO vector DNA template. Then, homologous knockout boxes S1H and S2H of2 were constructed by overlap PCR. Finally,2-knockout strains with stable inheritance were obtained by PEG-mediated protoplast transformation of the homologous knockout boxes. By comparing the biological characteristics of the knockout strains (Δ2) to the wild-type strains (WT), the effects of MAD2 protein on thecharacteristics ofgrowth, virulence, adhesion and inducing the peanut response of symbiosis genes were analyzed. 【Result】homologous recombination transformants with2 knockout were obtained by protoplast transformation.The spore germination rate of Δ2 was significantly decreased and spore semi-germination time was significantly prolonged 5.47 h compare to WT, as well as the mycelium length of Δ2was significantly shorter than WT in 12 h and 14 h incubation, which occupied 77.8% and 76.3% of WT, respectively. The sporulation in 12-day incubation was reduced by 33.3% compared to WT. The ability of Δ2 strain adhesion in onion was significantly decreased but showed no difference in adhere to the underwings of locust.In addition,2knockout did not affect the virulence ofto silkworm. In the peanut inoculated2-knockout strain for 12 h, the transcription level of symbiosis receptor gene,calcium signal decoding related genes (,,), lipid and nitrogen transfer related genes (1,24,2) was significantly down-regulated compared to the treatment of wt. While compared with blank control, Δ2 still had certainly up-regulatedtranscription level, significantly inhibited the transcription levels of,and, but had no effect on the transcription level of2,1 and24. 【Conclusion】adhesin protein MAD2 affects spore germination, initial growth of mycelium, sporulation quantity and plants adhesion, while has no effect on insect adhesion and virulence of strain, and MAD2 triggers the transcription of peanut symbiotic genes at the initial stage of interaction between the strain and peanut.
; adhesin protein MAD2; gene knockout; adhesion; virulence; growth trait
2021-04-14;
2021-05-12
國家重點研發計劃(2017YFD0201205,2018YFD0201002)、國家科技基礎資源調查專項(2019FY102002)、國家引才引智示范基地(YZJD202001045)
蔡霓,E-mail:741347573@qq.com。通信作者農向群,E-mail:xqnong@sina.com。通信作者王廣君,E-mail:guangjunwang@hotmail.com
(責任編輯 岳梅)
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
