工程酵母助力真菌嵌合萜類合酶的快速系統(tǒng)挖掘
2021-11-29 06:40:32范震潘海學(xué)唐功利
合成生物學(xué) 2021年5期
關(guān)鍵詞:功能
范震,潘海學(xué),唐功利
(1 中國科學(xué)院大學(xué)杭州高等研究院,化學(xué)與材料科學(xué)學(xué)院,浙江 杭州 310024;2 中國科學(xué)院上海有機(jī)化學(xué)研究所,生命有機(jī)化學(xué)國家重點(diǎn)實(shí)驗(yàn)室,上海 200032)
萜類是自然界中數(shù)量最龐大的一類天然產(chǎn)物,至今已有超過10 萬種[1-2]。萜類存在于所有生命領(lǐng)域中,在植物、真菌和海洋無脊椎動(dòng)物中最為普遍,萜類化合物在防御、調(diào)節(jié)和信息交流等方面發(fā)揮著重要作用,并被廣泛用作藥物、香料和調(diào)味品等[3-4]。盡管萜類數(shù)量龐大且結(jié)構(gòu)多樣,但所有的萜類天然產(chǎn)物骨架都由五碳結(jié)構(gòu)單元二甲基烯丙基焦磷酸(dimethylallyl pyrophosphate,DMAPP)和異戊烯基焦磷酸(isopentenyl pyrophosphate,ⅠPP)縮合形成,因此萜類也被稱為類異戊二烯類化合物[2]。五碳結(jié)構(gòu)單元DMAPP和ⅠPP在異戊烯基轉(zhuǎn)移酶(PT)的催化下,以常規(guī)的頭尾連接的方式縮合成不同長(zhǎng)度聚異戊二烯基焦磷酸,如FPP(farnesyl diphosphate)、GPP(geranyl diphosphate)、GGPP(geranylgeranyl diphosphate)、GFPP(geranylfranesyl diphosphate)(圖 1)[2]。除此之外,PT 還可以催化非常規(guī)類型的類異戊二烯焦磷酸的縮合,如以非頭尾連接方式的碳鏈縮合[5],順式(Z型)雙鍵的碳鏈縮合[6]等,因評(píng)論的工作中不涉及該類PT,本文對(duì)此不做過多的評(píng)述。萜類合酶(TS)利用PT 合成的鏈狀前體合成萜類的基本骨架,在萜類化合物的形成和萜類結(jié)構(gòu)的多樣性中發(fā)揮了至關(guān)重要的作用[2]。PT 和TS 負(fù)責(zé)形成的萜類基本骨架,再經(jīng)過各種修飾,如氧化、官能團(tuán)引入等,最終生成更加多樣的萜類產(chǎn)物。
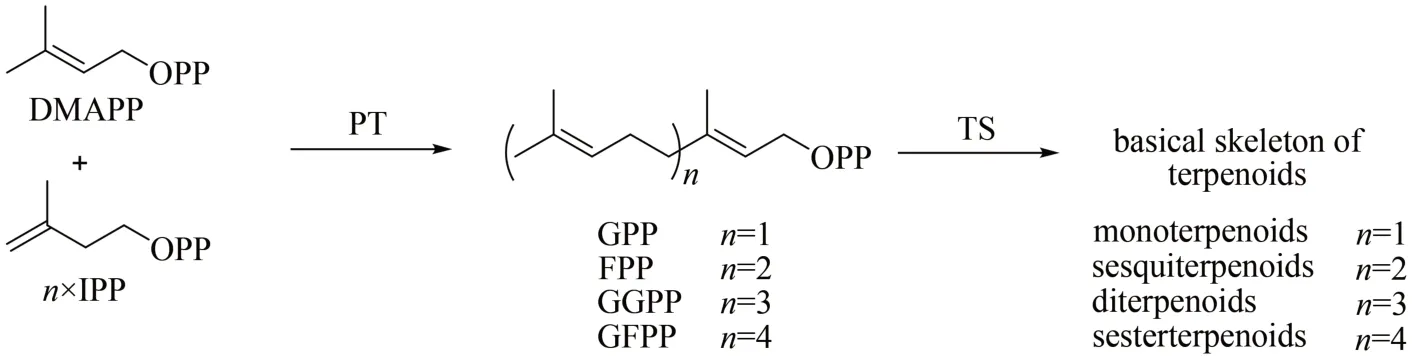
圖1 異戊烯基轉(zhuǎn)移酶(PT)和萜類合酶(TS)催化萜類基本骨架形成的簡(jiǎn)略過程Fig.1 Formation of terpenoid skeletons through the catalysis by isopentenyl transferase(PT)and terpene synthase(TS)
TS 催化萜類基本骨架形成過程中,首先形成高活性的碳正離子中間體,從而引發(fā)環(huán)化串聯(lián)反應(yīng),該環(huán)化串聯(lián)反應(yīng)也是自然界中最復(fù)雜的反應(yīng)之一[2]。根據(jù)TS起始引發(fā)碳正離子形成策略的不同,將TS 分為Ⅰ型和Ⅱ型[2]。Ⅰ型萜類合酶含有富含天冬氨酸(Asp)和與金屬陽離子配位的兩個(gè)保守基序DDXXD/E和(N,D)XX(S,T)XXXE,金屬離子通過結(jié)合聚異戊二烯基焦磷酸鏈狀前體的焦磷酸基團(tuán)來固定底物,同時(shí)促進(jìn)前體烯丙基焦磷酸基的C—O 鍵的電離,從而產(chǎn)生高活性的碳正離子中間體,進(jìn)而引發(fā)環(huán)化串聯(lián)反應(yīng)[圖2(a)]。而Ⅱ型萜類合酶通常僅含有呈酸性的DXDD 保守基序,通過其中的Asp 殘基將異戊二烯基中的雙鍵質(zhì)子化,生成引發(fā)環(huán)化反應(yīng)的碳正離子中間體[圖2(b)][7-9]。

圖2 Ⅰ型和Ⅱ型萜類合酶(TS)催化環(huán)化的過程的兩例Fig.2 Two cases for cyclization catalyzed by TypeⅠand TypeⅡterpene synthases(TSs)
通常情況下TS和PT單獨(dú)行使其功能,但在一些酶中兩個(gè)(TS 和PT/TS)活性結(jié)構(gòu)域同時(shí)存在,構(gòu)成可連續(xù)反應(yīng)的兩個(gè)獨(dú)特活性位點(diǎn),因此賦予這些酶雙功能性[10]。目前發(fā)現(xiàn)的雙功能萜類合酶有:Ⅰ型TS+Ⅱ型TS 酶,Ⅱ型TS+PT 酶,Geosmin合成酶,以及Ⅰ型 TS+PT 酶(圖 3)[10]。在理論上,這些雙功能酶兩個(gè)活性位點(diǎn)空間上是相互接近的,因此具有增加產(chǎn)物量的優(yōu)勢(shì)[10-12]。

圖3 4種嵌合萜類合酶催化反應(yīng)范例Fig.3 Catalytic reactions of 4 chimeric terpene synthases
真菌特有的嵌合萜類合酶(PTTS)指的是Ⅰ型TS+PT 酶,包括一個(gè)C 端PT 結(jié)構(gòu)域和一個(gè)N 端Ⅰ型TS 結(jié)構(gòu)域。這些PTTS 可以直接利用ⅠPP和DMAPP 作為共底物形成二萜和二倍半萜骨架[10,13]。自嵌合二萜合酶 PaFS 和嵌合二倍半萜合酶 AcOS 發(fā)現(xiàn) 以來[14-15], 僅有 約 20 個(gè) PTTS 被報(bào)道,并且PTTS 催化生成的化合物大多具有新穎的碳骨架。根據(jù)其催化環(huán)化的機(jī)制,可將PTTS 分為A 型和 B 型,A 型是在 GFPP 的 C1-C11/C10-C14 之間環(huán)化(C1-Ⅳ-Ⅴ),形成5~15環(huán)系統(tǒng),B型是在GFPP 或 GGPP 的 C1-C15/C14-C18 處環(huán)化 (C1-Ⅲ-Ⅳ),而生成5~11環(huán)系統(tǒng)(圖4)[13]。
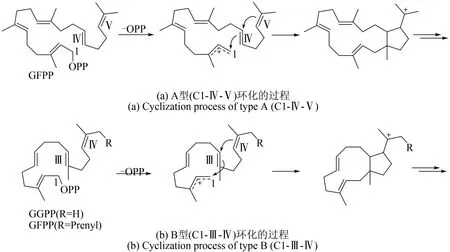
圖4 PTTS催化的兩種環(huán)化類型Fig.4 Two cyclization reactions catalyzed by PTTS
隨著基因組測(cè)序技術(shù)的發(fā)展,需要功能鑒定的PTTS 數(shù)量不斷增加,對(duì)PTTS 研究產(chǎn)生了一些有待回答的問題,如PTTS 在真菌中是廣泛分布的還是局限于某些特定的群體,以及哪些機(jī)制控制著PTTS 的功能進(jìn)化等,回答這些問題需要對(duì)大量的 PTTS 進(jìn) 行 功 能 表 征[16]。 雖 然 米 曲 霉(Aspergillus oryzae)已經(jīng)被廣泛應(yīng)用于真菌基因的異源表達(dá),且該體系表征過多種絲狀真菌來源的PTTS[17-18],然而受限于米曲霉復(fù)雜的遺傳操作、較長(zhǎng)的培養(yǎng)周期,這個(gè)系統(tǒng)不適合大量未知PTTS功能的快速驗(yàn)證。高效萜類前體供給的大腸桿菌和釀酒酵母底盤,為法尼烯、紫杉二烯和青蒿酸[19-21]等的大量生產(chǎn)提供了有效的途徑。此外,以代謝工程改造的酵母來進(jìn)行藥物核心結(jié)構(gòu)骨架的生產(chǎn),隨后經(jīng)過簡(jiǎn)單的化學(xué)后修飾可合成最終藥品,極大簡(jiǎn)化了藥物化學(xué)合成的過程(圖5)[22-23]。最近武漢大學(xué)劉天罡教授課題組與美國田納西大學(xué)陳峰教授課題組合作,使用高產(chǎn)萜類前體的酵母系統(tǒng)和高通量自動(dòng)化平臺(tái)實(shí)現(xiàn)了對(duì)PTTS 功能的快速驗(yàn)證[16]。
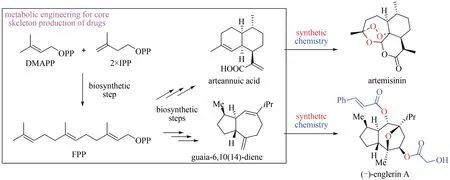
圖5 代謝工程結(jié)合化學(xué)合成的方法合成青蒿素和(-)-englerin AFig.5 Synthesis of artemisinin and(-)-englerin A by metabolic engineering combined with synthetic chemistry
首先,合作團(tuán)隊(duì)系統(tǒng)分析了NCBⅠ和UniProt數(shù)據(jù)庫中所有真菌來源的基因以及JGⅠ數(shù)據(jù)庫中已經(jīng)測(cè)序的477 種真菌的基因組,共找到了227 個(gè)PTTS 同源基因,其中包括已經(jīng)鑒定功能的20 個(gè)PTTS基因。對(duì)這227個(gè)PTTS同源基因的生物信息學(xué)分析發(fā)現(xiàn),PTTS 基因只存在于雙核菌亞界(Dikarya),其中224 個(gè)分布在子囊菌(Ascomycota)中, 另外 3 個(gè) PTTS 基因存在于擔(dān)子菌(Basidiomycota)的一個(gè)種中。由此作者推測(cè),PTTS 基因起源于雙核菌亞界,擔(dān)子菌中PTTS 基因的缺失,很可能是在進(jìn)化過程中經(jīng)歷了頻繁的基因丟失導(dǎo)致的;另一種可能性是PTTS 基因起源于子囊菌,擔(dān)子菌中某些菌含有的PTTS 基因是通過基因的水平轉(zhuǎn)移獲得的。為了進(jìn)一步了解這些基因進(jìn)化的關(guān)聯(lián)性,研究者對(duì)227 個(gè)PTTS 基因進(jìn)行了系統(tǒng)的進(jìn)化分析。前期研究報(bào)道,PTTS 可分為 6 個(gè)分支(A~F)[10],處于 A、E 和 F 亞家族中的基因形成CladeⅠ,催化GFPP的A型(C1-Ⅳ-Ⅴ)環(huán)化,其余亞家族的基因形成Clade Ⅱ,催化GGPP 或 GFPP 的 B 型(C1-Ⅲ-Ⅳ)環(huán)化[10]。考慮到之前研究PTTS 數(shù)量較少,信息不夠全面,作者對(duì)得到的PTTS 做了更全面的進(jìn)化分析,結(jié)果顯示支持前期研究結(jié)果。并且進(jìn)化分析的結(jié)果顯示,在A~F 這6 個(gè)PTTS 亞家族中,每個(gè)亞家族都有90%以上的PTTS 基因功能是未知的,表明對(duì)PTTS生化功能認(rèn)識(shí)上還存在很大的不足[16]。
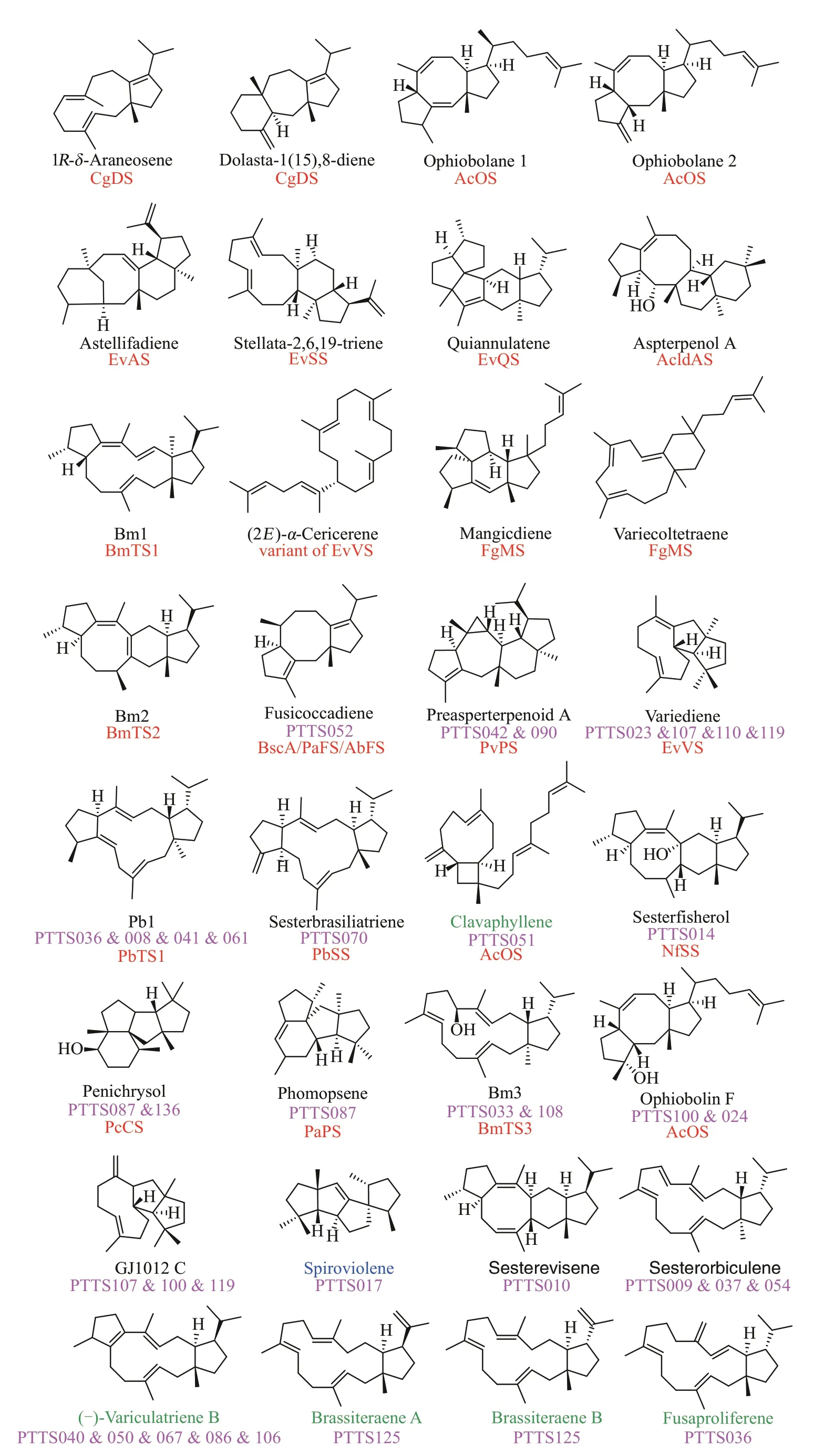
為了獲得更多的PTTS 催化功能信息,合作團(tuán)隊(duì)對(duì)上述PTTS 進(jìn)行了功能鑒定。首先依據(jù)氨基酸(aa)序列長(zhǎng)度(約700 aa)、序列相似性(<80%)、TS 結(jié)構(gòu)域 DDXXD/E 和 NSE/DTE 以及 PT 結(jié)構(gòu)域DDXXD/N 的保守序列這些特征,對(duì)PTTS 進(jìn)行了篩選,從中選出了74 個(gè)PTTS。因已報(bào)道的20 個(gè)PTTS 主要催化二倍半萜的產(chǎn)生,因此研究者在實(shí)驗(yàn)室前期構(gòu)建的高產(chǎn)萜類化合物的釀酒酵母Saccharomyces cerevisiaeYZL141 的基礎(chǔ)上[11],又過量表達(dá)了GFPP 合成酶和PTTS,構(gòu)建了一個(gè)高產(chǎn)二倍半萜的酵母底盤。該底盤一方面增加了GFPP 的代謝通量,使得嵌合的二倍半萜合酶的功能可得到快速鑒定,另一方面有助于釋放嵌合的二萜合酶利用GFPP 的潛力,使得嵌合的二萜合酶在合成二萜的同時(shí)還可合成二倍半萜。此外,他們利用高通量自動(dòng)化平臺(tái)批量構(gòu)建了74個(gè)PTTS的酵母工程菌株,該平臺(tái)的應(yīng)用大大加快了PTTS 功能鑒定的進(jìn)程。利用上述代謝工程策略,合作團(tuán)隊(duì)成功鑒定34個(gè)新的有功能的PTTS(圖6),鑒定的PTTS 的數(shù)量超過了目前已知功能PTTS 的總數(shù)。從34 個(gè)PTTS 的酵母工程菌株中共鑒定了24 個(gè)二萜和二倍半萜產(chǎn)物(圖6),其中2個(gè)是結(jié)構(gòu)新穎的二倍半萜 sesterevisene 和 sesterorbiculene。24 個(gè)產(chǎn)物中有11個(gè)是已報(bào)道的PTTS的產(chǎn)物(圖6),這表明這類酶在功能上進(jìn)化的一致性。有5個(gè)產(chǎn)物是已報(bào)道的植物來源二倍半萜合酶的產(chǎn)物(圖6),有1 個(gè)產(chǎn)物是鏈霉菌(Streptomyces violens)來源的二萜合酶產(chǎn)物(圖6),顯示了真菌、植物和細(xì)菌的基因功能趨同進(jìn)化的特征。54 種活性PTTS 共產(chǎn)37 種不同的萜類產(chǎn)物(圖6),以及新化合物的發(fā)現(xiàn)和鏈狀產(chǎn)物的產(chǎn)生(圖6)都表明PTTS 在進(jìn)化過程中蛋白質(zhì)序列的變化導(dǎo)致其功能多樣性。

圖6 54個(gè)PTTS催化合成的萜類化合物及其結(jié)構(gòu)Fig.6 Terpenoids and their structures synthesized under the catalysis of 54 PTTSs
采用酵母底盤細(xì)胞對(duì)PTTS 的起源和功能進(jìn)化進(jìn)行的系統(tǒng)研究,為讀者清晰地呈現(xiàn)了真菌特有的這類PTTS 在真菌中存在和進(jìn)化的基本樣貌,描述了這類酶催化功能的多樣性,并且大大擴(kuò)充了這類酶家族成員的數(shù)量以及萜類產(chǎn)物庫,為后續(xù)的萜類合成生物學(xué)研究提供了豐富的基因元件,也為相關(guān)基因簇的挖掘奠定了堅(jiān)實(shí)的基礎(chǔ)。另外,該研究不僅為以酵母為底盤進(jìn)行萜類化合物的挖掘提供了一個(gè)可借鑒有效的方法策略,而且也是將天然產(chǎn)物的發(fā)現(xiàn)從原始的自然界生物材料中鑒定,轉(zhuǎn)變?yōu)閺木W(wǎng)絡(luò)基因信息挖掘到人為設(shè)計(jì)實(shí)現(xiàn)生產(chǎn)的一個(gè)成功范例。
猜你喜歡
鐘表(2023年5期)2023-10-27 04:20:44
中華詩詞(2022年6期)2022-12-31 06:41:24
當(dāng)代陜西(2021年21期)2022-01-19 02:00:26
中學(xué)生數(shù)理化(高中版.高考數(shù)學(xué))(2020年1期)2020-02-20 13:23:44
經(jīng)濟(jì)技術(shù)協(xié)作信息(2018年11期)2019-01-14 03:07:20
中國科技論壇(2017年7期)2017-07-25 08:49:53
制造技術(shù)與機(jī)床(2017年3期)2017-06-23 08:11:33
媽媽寶寶(2017年2期)2017-02-21 01:21:24
國際漢語學(xué)報(bào)(2016年1期)2017-01-20 08:21:20
中國中醫(yī)藥現(xiàn)代遠(yuǎn)程教育(2014年22期)2014-03-01 04:32:55
